
Bone Morphogenetic Protein-4 Impairs Retinal Endothelial Cell Barrier, a Potential Role in Diabetic Retinopathy
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Postmortem Human Retina Samples
2.2. Experimental Mice
2.3. Human Retinal Endothelial Cells
2.4. Assessment of BMP4 Expression and Localization in Human and Mouse Retinas
2.5. In Vitro Studies
2.5.1. Assessment of ZO-1 Distribution in HRECs Cells Using Immunofluorescence Staining
2.5.2. Assessment of Human Retinal Endothelial Cell Barrier Function
2.5.3. Assessment of the Effect of BMP4 Treatment on Phospho-Smad 1/5/9 and Phospho-p38 Levels
2.5.4. Measurement of Matrix Metalloproteinase Activity (MMPs) Activities
2.5.5. Statistical Analysis
3. Results
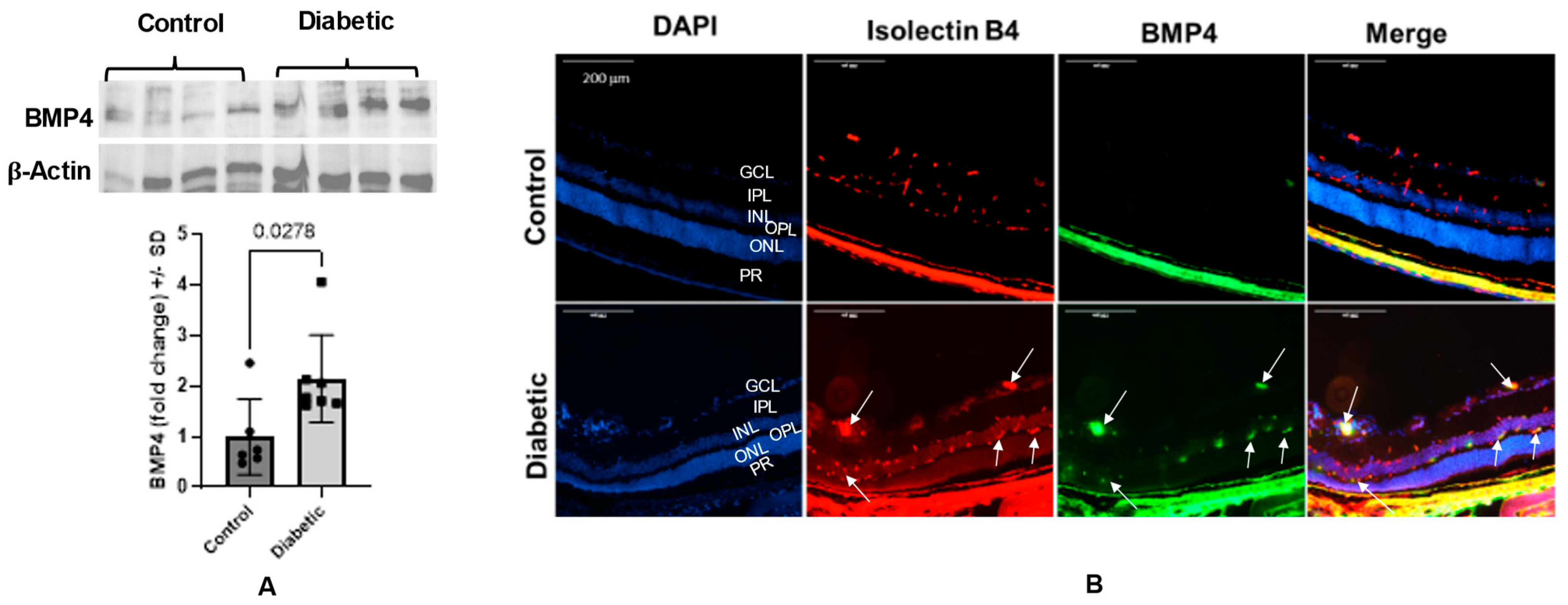
3.1. Diabetes Upregulates Retinal BMP4 in Human Subjects and Experimental Mice
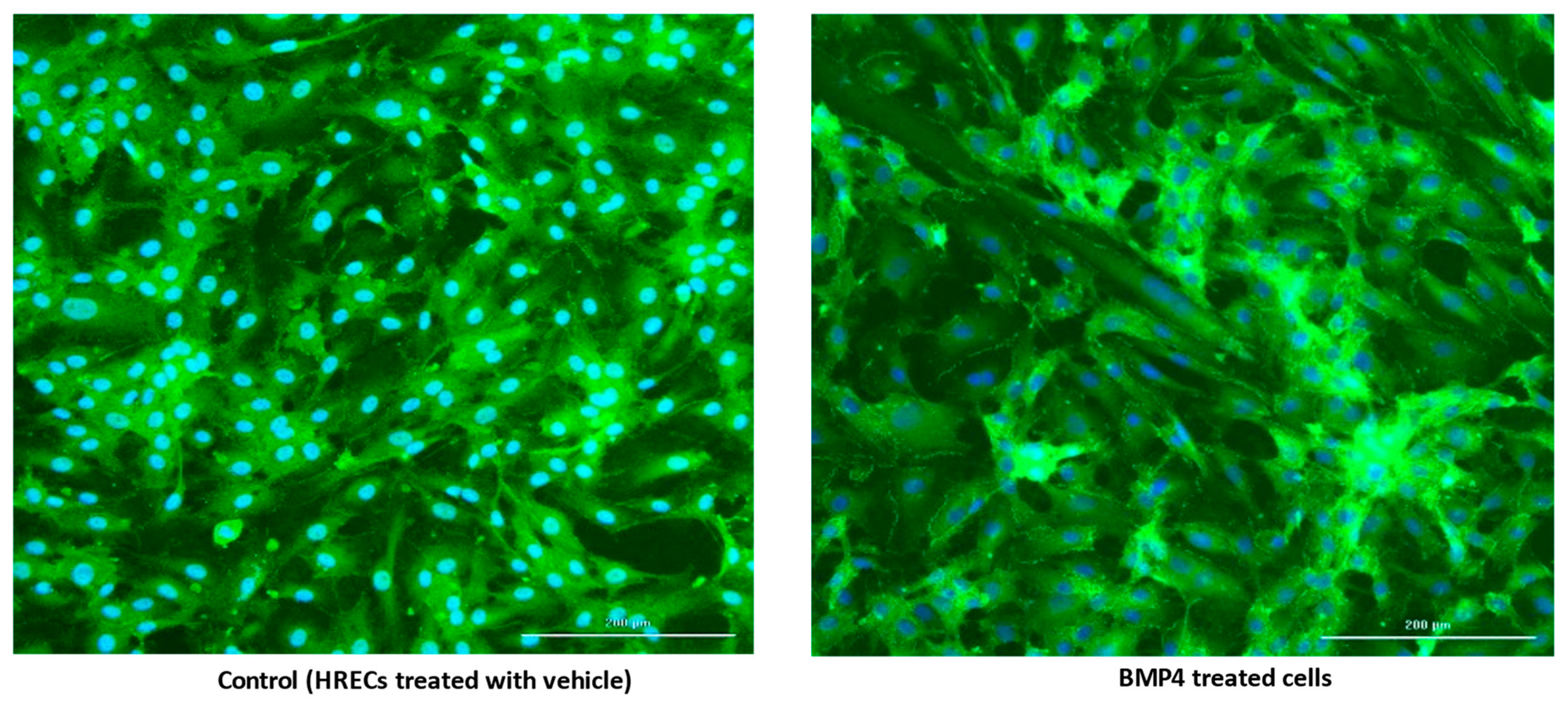
3.2. BMP4 Disrupts Human Retinal Endothelial Barrier Function In Vitro
3.3. BMP4 Activates the Canonical Smad Pathway and Non-Canonical p38-MAPK Pathway in HRECs
3.4. BMP4 Increases the Activity of Matrix Metalloproteinases (MMPs) in HRECs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- He, X.; Qi, S.; Zhang, X.; Pan, J. The relationship between the neutrophil-to-lymphocyte ratio and diabetic retinopathy in adults from the United States: Results from the National Health and nutrition examination survey. BMC Ophthalmol. 2022, 22, 346. [Google Scholar] [CrossRef] [PubMed]
- Taurone, S.; De Ponte, C.; Rotili, D.; De Santis, E.; Mai, A.; Fiorentino, F.; Scarpa, S.; Artico, M.; Micera, A. Biochemical functions and clinical characterizations of the sirtuins in diabetes-induced retinal pathologies. Int. J. Mol. Sci. 2022, 23, 4048. [Google Scholar] [CrossRef] [PubMed]
- Simo, R.; Hernandez, C. Novel approaches for treating diabetic retinopathy based on recent pathogenic evidence. Prog. Retin. Eye Res. 2015, 48, 160–180. [Google Scholar] [CrossRef]
- Rahman, M.S.; Akhtar, N.; Jamil, H.M.; Banik, R.S.; Asaduzzaman, S.M. TGF-β/BMP signaling and other molecular events: Regulation of osteoblastogenesis and bone formation. Bone Res. 2015, 3, 15005. [Google Scholar] [CrossRef]
- Shimasaki, S.; Moore, R.K.; Otsuka, F.; Erickson, G.F. The Bone Morphogenetic Protein System In Mammalian Reproduction. Endocr. Rev. 2004, 25, 72–101. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, T.; Watabe, T. Bone Morphogenetic Proteins. Cold Spring Harb. Perspect. Biol. 2016, 8, 233–241. [Google Scholar] [CrossRef]
- Dituri, F.; Cossu, C.; Mancarella, S.; Giannelli, G. The Interactivity between TGFβ and BMP Signaling in Organogenesis, Fibrosis, and Cancer. Cells 2019, 8, 1130. [Google Scholar] [CrossRef]
- Jensen, G.S.; Leon-Palmer, N.E.; Townsend, K.L. Bone morphogenetic proteins (BMPs) in the central regulation of energy balance and adult neural plasticity. Metabolism 2021, 123, 154837. [Google Scholar] [CrossRef]
- Yang, P.; Troncone, L.; Augur, Z.M.; Kim, S.S.J.; McNeil, M.E.; Yu, P.B. The role of bone morphogenetic protein signaling in vascular calcification. Bone 2020, 141, 115542. [Google Scholar] [CrossRef]
- Lee, H.W.; Chong, D.C.; Ola, R.; Dunworth, W.P.; Meadows, S.; Ka, J.; Kaartinen, V.M.; Qyang, Y.; Cleaver, O.; Bautch, V.L.; et al. Alk2/ACVR1 and Alk3/BMPR1A Provide Essential Function for Bone Morphogenetic Protein-Induced Retinal Angiogenesis. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.; Lee, H.W.; Pak, B.; Han, O.; Kim, M.; Jin, S.W. Transcriptomic analysis identifies novel targets for individual bone morphogenetic protein type 1 receptors in endothelial cells. FASEB J. 2021, 35, e21386. [Google Scholar] [CrossRef] [PubMed]
- Al-Shabrawey, M.; Hussein, K.; Wang, F.; Wan, M.; Elmasry, K.; Elsherbiny, N.; Saleh, H.; Yu, P.B.; Tawfik, A.; Ibrahim, A.S. Bone morphogenetic protein-2 induces non-canonical inflammatory and oxidative pathways in human retinal endothelial cells. Front. Immunol. 2021, 11, 568795. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Duffhues, G.; Williams, E.; Goumans, M.-J.; Heldin, C.-H.; Ten Dijke, P. Bone morphogenetic protein receptors: Structure, function and targeting by selective small molecule kinase inhibitors. Bone 2020, 138, 115472. [Google Scholar] [CrossRef] [PubMed]
- Vania, V.; Wang, L.; Tjakra, M.; Zhang, T.; Qiu, J.; Tan, Y.; Wang, G. The interplay of signaling pathway in endothelial cells—Matrix stiffness dependency with targeted-therapeutic drugs. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2020, 1866, 165645. [Google Scholar] [CrossRef]
- Cai, J.; Pardali, E.; Sánchez-Duffhues, G.; ten Dijke, P. BMP signaling in vascular diseases. FEBS Lett. 2012, 586, 1993–2002. [Google Scholar] [CrossRef]
- Dyer, L.A.; Pi, X.; Patterson, C. The role of BMPs in endothelial cell function and dysfunction. Trends Endocrinol. Metab. 2014, 25, 472–480. [Google Scholar] [CrossRef]
- Gallardo-Vara, E.; Gamella-Pozuelo, L.; Perez-Roque, L.; Bartha, J.L.; Garcia-Palmero, I.; Casal, J.I.; López-Novoa, J.M.; Pericacho, M.; Bernabeu, C. Potential role of circulating endoglin in hypertension via the upregulated expression of BMP4. Cells 2020, 9, 988. [Google Scholar] [CrossRef]
- Wang, L.; Cheng, C.K.; Yi, M.; Lui, K.O.; Huang, Y. Targeting endothelial dysfunction and inflammation. J. Mol. Cell. Cardiol. 2022, 168, 58–67. [Google Scholar] [CrossRef]
- Jank, M.; von Niessen, N.; Olivier, C.B.; Schmitt, H.; Anto-Michel, N.; Hilgendorf, I.; Bode, C.; Moser, M.; Esser, J.S.; Zhou, Q. Platelet Bone Morphogenetic Protein-4 Mediates Vascular Inflammation and Neointima Formation after Arterial Injury. Cells 2021, 10, 2027. [Google Scholar] [CrossRef]
- Ibrahim, A.S.; Tawfik, A.M.; Hussein, K.A.; Elshafey, S.; Markand, S.; Rizk, N.; Duh, E.J.; Smith, S.B.; Al-Shabrawey, M. Pigment epithelium-derived factor inhibits retinal microvascular dysfunction induced by 12/15-lipoxygenase-derived eicosanoids. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2015, 1851, 290–298. [Google Scholar] [CrossRef]
- Ibrahim, A.S.; Hussein, K.; Wang, F.; Wan, M.; Saad, N.; Essa, M.; Kim, I.; Shakoor, A.; Owen, L.A.; DeAngelis, M.M.; et al. Bone Morphogenetic Protein (BMP)4 But Not BMP2 Disrupts the Barrier Integrity of Retinal Pigment Epithelia and Induces Their Migration: A Potential Role in Neovascular Age-Related Macular Degeneration. J. Clin. Med. 2020, 9, 2293. [Google Scholar] [CrossRef] [PubMed]
- Benn, A.; Hiepen, C.; Osterland, M.; Schütte, C.; Zwijsen, A.; Knaus, P. Role of bone morphogenetic proteins in sprouting angiogenesis: Differential BMP receptor-dependent signaling pathways balance stalk vs. tip cell competence. FASEB J. 2017, 31, 4720. [Google Scholar] [CrossRef] [PubMed]
- de Vinuesa, A.G.; Abdelilah-Seyfried, S.; Knaus, P.; Zwijsen, A.; Bailly, S. BMP signaling in vascular biology and dysfunction. Cytokine Growth Factor Rev. 2016, 27, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Groppe, J.; Greenwald, J.; Wiater, E.; Rodriguez-Leon, J.; Economides, A.N.; Kwiatkowski, W.; Affolter, M.; Vale, W.W.; Izpisua Belmonte, J.C.; Choe, S. Structural basis of BMP signalling inhibition by the cystine knot protein Noggin. Nature 2002, 420, 636–642. [Google Scholar] [CrossRef] [PubMed]
- Cuny, G.D.; Paul, B.Y.; Laha, J.K.; Xing, X.; Liu, J.-F.; Lai, C.S.; Deng, D.Y.; Sachidanandan, C.; Bloch, K.D.; Peterson, R.T. Structure–activity relationship study of bone morphogenetic protein (BMP) signaling inhibitors. Bioorg. Med. Chem. Lett. 2008, 18, 4388–4392. [Google Scholar] [CrossRef] [PubMed]
- Mohedas, A.H.; Xing, X.; Armstrong, K.A.; Bullock, A.N.; Cuny, G.D.; Yu, P.B. Development of an ALK2-biased BMP type I receptor kinase inhibitor. ACS Chem. Biol. 2013, 8, 1291–1302. [Google Scholar] [CrossRef]
- Ibrahim, A.S.; Saleh, H.; El-Shafey, M.; Hussein, K.A.; El-Masry, K.; Baban, B.; Sheibani, N.; Wang, M.-H.; Tawfik, A.; Al-Shabrawey, M. Targeting of 12/15-Lipoxygenase in retinal endothelial cells, but not in monocytes/macrophages, attenuates high glucose-induced retinal leukostasis. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2017, 1862, 636–645. [Google Scholar] [CrossRef]
- Ibrahim, A.S.; Elshafey, S.; Sellak, H.; Hussein, K.A.; El-Sherbiny, M.; Abdelsaid, M.; Rizk, N.; Beasley, S.; Tawfik, A.M.; Smith, S.B. A lipidomic screen of hyperglycemia-treated HRECs links 12/15-Lipoxygenase to microvascular dysfunction during diabetic retinopathy via NADPH oxidase. J. Lipid Res. 2015, 56, 599–611. [Google Scholar] [CrossRef]
- Ibrahim, A.S.; Mander, S.; Hussein, K.A.; Elsherbiny, N.M.; Smith, S.B.; Al-Shabrawey, M.; Tawfik, A. Hyperhomocysteinemia disrupts retinal pigment epithelial structure and function with features of age-related macular degeneration. Oncotarget 2016, 7, 8532–8545. [Google Scholar] [CrossRef]
- Szulcek, R.; Bogaard, H.J.; van Nieuw Amerongen, G.P. Electric cell-substrate impedance sensing for the quantification of endothelial proliferation, barrier function, and motility. J. Vis. Exp. 2014, 85, e51300. [Google Scholar]
- Cabral-Pacheco, G.A.; Garza-Veloz, I.; Castruita-De la Rosa, C.; Ramirez-Acuña, J.M.; Perez-Romero, B.A.; Guerrero-Rodriguez, J.F.; Martinez-Avila, N.; Martinez-Fierro, M.L. The Roles of Matrix Metalloproteinases and Their Inhibitors in Human Diseases. Int. J. Mol. Sci. 2020, 21, 9739. [Google Scholar] [CrossRef] [PubMed]
- Hussein, K.A.; Choksi, K.; Akeel, S.; Ahmad, S.; Megyerdi, S.; El-Sherbiny, M.; Nawaz, M.; El-Asrar, A.A.; Al-Shabrawey, M. Bone morphogenetic protein 2: A potential new player in the pathogenesis of diabetic retinopathy. Exp. Eye Res. 2014, 125, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Mattar, T.; Friedrich, P.F.; Bishop, A.T. Effect of rhBMP-2 and VEGF in a vascularized bone allotransplant experimental model based on surgical neoangiogenesis. J. Orthop. Res. 2013, 31, 561–566. [Google Scholar] [CrossRef]
- Weiss, K.R.; Cooper, G.M.; Jadlowiec, J.A.; McGough, R.L., III; Huard, J. VEGF and BMP expression in mouse osteosarcoma cells. Clin. Orthop. Relat. Res. 2006, 450, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Zhang, Z.; Liu, X.; Wang, Q.; Hong, Y.; Li, X.; Liu, J. RNA sequencing reveals BMP4 as a basis for the dual-target treatment of diabetic retinopathy. J. Mol. Med. 2021, 99, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Pouliot, F.; Blais, A.; Labrie, C. Overexpression of a dominant negative type II bone morphogenetic protein receptor inhibits the growth of human breast cancer cells. Cancer Res. 2003, 63, 277–281. [Google Scholar] [PubMed]
- Townson, S.A.; Martinez-Hackert, E.; Greppi, C.; Lowden, P.; Sako, D.; Liu, J.; Ucran, J.A.; Liharska, K.; Underwood, K.W.; Seehra, J. Specificity and structure of a high affinity activin receptor-like kinase 1 (ALK1) signaling complex. J. Biol. Chem. 2012, 287, 27313–27325. [Google Scholar] [CrossRef]
- Voeltzel, T.; Flores-Violante, M.; Zylbersztejn, F.; Lefort, S.; Billandon, M.; Jeanpierre, S.; Joly, S.; Fossard, G.; Milenkov, M.; Mazurier, F.; et al. A new signaling cascade linking BMP4, BMPR1A, ΔNp73 and NANOG impacts on stem-like human cell properties and patient outcome. Cell Death Dis. 2018, 9, 1011. [Google Scholar] [CrossRef]
- Kulikauskas, M.R.; Bautch, V.L. The versatility and paradox of BMP signaling in endothelial cell behaviors and blood vessel function. Cell. Mol. Life Sci. 2022, 79, 77. [Google Scholar] [CrossRef]
- Mao, C.; Wang, Y.; Zhang, X.; Zheng, X.; Tang, T.; Lu, E. Double-edged-sword effect of IL-1β on the osteogenesis of periodontal ligament stem cells via crosstalk between the NF-κB, MAPK and BMP/Smad signaling pathways. Cell Death Dis. 2016, 7, e2296. [Google Scholar] [CrossRef]
- Raida, M.; Clement, J.H.; Leek, R.D.; Ameri, K.; Bicknell, R.; Niederwieser, D.; Harris, A.L. Bone morphogenetic protein 2 (BMP-2) and induction of tumor angiogenesis. J. Cancer Res. Clin. Oncol. 2005, 131, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Akeel, S.; El-Awady, A.; Hussein, K.; El-Refaey, M.; Elsalanty, M.; Sharawy, M.; Al-Shabrawey, M. Recombinant bone morphogenetic protein-2 induces up-regulation of vascular endothelial growth factor and interleukin 6 in human pre-osteoblasts: Role of reactive oxygen species. Arch. Oral Biol. 2012, 57, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, H.; Qiu, G.; Su, X.; Wu, Z. Synergistic Effects of Vascular Endothelial Growth Factor on Bone Morphogenetic Proteins Induced Bone Formation In Vivo: Influencing Factors and Future Research Directions. BioMed Res. Int. 2016, 2016, 2869572. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Chen, X. Transcription regulation of the vegf gene by the BMP/Smad pathway in the angioblast of zebrafish embryos. Biochem. Biophys. Res. Commun. 2005, 329, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Harding, A.; Cortez-Toledo, E.; Magner, N.L.; Beegle, J.R.; Coleal-Bergum, D.P.; Hao, D.; Wang, A.; Nolta, J.A.; Zhou, P. Highly efficient differentiation of endothelial cells from pluripotent stem cells requires the MAPK and the PI3K pathways. Stem Cells 2017, 35, 909–919. [Google Scholar] [CrossRef]
- Esser, J.S.; Steiner, R.E.; Deckler, M.; Schmitt, H.; Engert, B.; Link, S.; Charlet, A.; Patterson, C.; Bode, C.; Zhou, Q. Extracellular bone morphogenetic protein modulator BMPER and twisted gastrulation homolog 1 preserve arterial-venous specification in zebrafish blood vessel development and regulate Notch signaling in endothelial cells. FEBS J. 2018, 285, 1419–1436. [Google Scholar] [CrossRef]
- Perera, N.; Ritchie, R.H.; Tate, M. The role of bone morphogenetic proteins in diabetic complications. ACS Pharmacol. Transl. Sci. 2019, 3, 11–20. [Google Scholar] [CrossRef]
- Ran, R.J.; Zheng, X.Y.; Du, L.P.; Zhang, X.D.; Chen, X.L.; Zhu, S.Y. Upregulated inflammatory associated factors and blood-retinal barrier changes in the retina of type 2 diabetes mellitus model. Int. J. Ophthalmol. 2016, 9, 1591–1597. [Google Scholar] [CrossRef]
- Acharya, N.K.; Qi, X.; Goldwaser, E.L.; Godsey, G.A.; Wu, H.; Kosciuk, M.C.; Freeman, T.A.; Macphee, C.H.; Wilensky, R.L.; Venkataraman, V.; et al. Retinal pathology is associated with increased blood–retina barrier permeability in a diabetic and hypercholesterolaemic pig model: Beneficial effects of the LpPLA2 inhibitor Darapladib. Diabetes Vasc. Dis. Res. 2017, 14, 200–213. [Google Scholar] [CrossRef]
- Wang, M.H.; Ibrahim, A.S.; Hsiao, G.; Tawfik, A.; Al-Shabrawey, M. A novel interaction between soluble epoxide hydrolase and the AT1 receptor in retinal microvascular damage. Prostaglandins Other Lipid Mediat. 2020, 148, 106449. [Google Scholar] [CrossRef]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and biological attributes of matrix metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar] [PubMed]
- Kapoor, C.; Vaidya, S.; Wadhwan, V.; Kaur, G.; Pathak, A. Seesaw of matrix metalloproteinases (MMPs). J. Cancer Res. Ther. 2016, 12, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Kowluru, R.A.; Mishra, M. Regulation of matrix metalloproteinase in the pathogenesis of diabetic retinopathy. Prog. Mol. Biol. Transl. Sci. 2017, 148, 67–85. [Google Scholar] [PubMed]
- Kowluru, R.A.; Zhong, Q.; Santos, J.M. Matrix metalloproteinases in diabetic retinopathy: Potential role of MMP-9. Expert Opin. Investig. Drugs 2012, 21, 797–805. [Google Scholar] [CrossRef]
- Uemura, S.; Matsushita, H.; Li, W.; Glassford, A.J.; Asagami, T.; Lee, K.-H.; Harrison, D.G.; Tsao, P.S. Diabetes mellitus enhances vascular matrix metalloproteinase activity: Role of oxidative stress. Circ. Res. 2001, 88, 1291–1298. [Google Scholar] [CrossRef]
- Ünal, A.; Baykal, O.; Öztürk, N. Comparison of matrix metalloproteinase 9 and 14 levels in vitreous samples in diabetic and non-diabetic patients: A case control study. Int. J. Retin. Vitr. 2022, 8, 44. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Darwish, N.H.E.; Hussein, K.A.; Elmasry, K.; Ibrahim, A.S.; Humble, J.; Moustafa, M.; Awadalla, F.; Al-Shabrawey, M. Bone Morphogenetic Protein-4 Impairs Retinal Endothelial Cell Barrier, a Potential Role in Diabetic Retinopathy. Cells 2023, 12, 1279. https://doi.org/10.3390/cells12091279
Darwish NHE, Hussein KA, Elmasry K, Ibrahim AS, Humble J, Moustafa M, Awadalla F, Al-Shabrawey M. Bone Morphogenetic Protein-4 Impairs Retinal Endothelial Cell Barrier, a Potential Role in Diabetic Retinopathy. Cells. 2023; 12(9):1279. https://doi.org/10.3390/cells12091279
Chicago/Turabian StyleDarwish, Noureldien H. E., Khaled A. Hussein, Khaled Elmasry, Ahmed S. Ibrahim, Julia Humble, Mohamed Moustafa, Fatma Awadalla, and Mohamed Al-Shabrawey. 2023. "Bone Morphogenetic Protein-4 Impairs Retinal Endothelial Cell Barrier, a Potential Role in Diabetic Retinopathy" Cells 12, no. 9: 1279. https://doi.org/10.3390/cells12091279