Abstract
Fibrosing interstitial lung diseases (FILDs), e.g., due to idiopathic pulmonary fibrosis (IPF), are chronic progressive diseases with a poor prognosis. The management of these diseases is challenging and focuses mainly on the suppression of progression with anti-fibrotic drugs. Therefore, novel FILD treatments are needed. In recent years, cell-based therapy with various stem cells has been investigated for FILD, and the use of mesenchymal stem cells (MSCs) has been widely reported and clinical studies are also ongoing. Induced pluripotent stem cells (iPSCs) have also been reported to have an anti-fibrotic effect in FILD; however, these have not been as well studied as MSCs in terms of the mechanisms and side effects. While MSCs show a potent anti-fibrotic effect, the possibility of quality differences between donors and a stable supply in the case of donor shortage or reduced proliferative capacity after cell passaging needs to be considered. The application of iPSC-derived cells has the potential to overcome these problems and may lead to consistent quality of the cell product and stable product supply. This review provides an overview of iPSCs and FILD, followed by the current status of cell-based therapy for FILD, and then discusses the possibilities and perspectives of FILD therapy with iPSC-derived cells.
1. Introduction
Fibrotic interstitial lung disease (FILD) may have various causes and, depending on its etiology and progression, the course of the disease may be fatal. Although some patients may improve with anti-inflammatory treatment such as steroids and immunosuppressive drugs, idiopathic pulmonary fibrosis (IPF), the most common type of FILD, is relatively nonresponsive to such treatments and is often fatal because of its progressive course [1,2]. In cases of severe fibrosis, reversible changes are not expected, and further fibrosis is induced from the fibrotic areas, resulting in the progression of the disease [3]. In such cases, treatment focuses on controlling disease progression, and a radical cure is considered unlikely. Although lung transplantation is sometimes performed in severe treatment-refractory cases, the number of donors is limited, and not all critical patients are eligible to get transplanted [4]. In addition, even if transplantation was performed, survival rates are lower than in healthy or slowly progressing patients [4]. Therefore, there is an urgent and critical need for novel treatment modalities for progressive FILD.
Induced pluripotent stem cells (iPSCs) are artificially induced pluripotent stem cells that can be differentiated into specific cells for use in therapeutic applications [5]. iPSCs themselves have been reported to have anti-inflammatory effects, and studies are evaluating their potential clinical application as a cell-based therapy [6,7,8,9]. iPSCs also have a wide range of potential applications in drug discovery and disease research [10,11]. In recent years, iPSCs derived from various human leukocyte antigens (HLAs) have been prepared and stockpiled from the blood of healthy donors in Japan [12]. Matching these HLAs with recipients is enabling cell therapies with a reduced risk of immune rejection to be considered [12].
In recent years, clinical studies have evaluated the possibility of stem cell-based therapies for FILD. To date, there are no reports of clinical trials on iPSCs for the treatment of FILD; however, preclinical studies in animals have indicated that iPSCs may be effective. Nevertheless, the clinical application of undifferentiated iPSCs still faces a considerable number of challenges.
In this review, we will outline the current state of knowledge about cell-based cell therapy for the treatment of FILD, which has received much attention in recent years, and discuss the potential and future prospects of cell-based therapy with iPSC-derived cells.
1.1. An Overview of FILD
The most common cause of FILD is idiopathic pulmonary fibrosis (IPF), but there is a wide range of other causes, including fibrotic hypersensitivity pneumonitis, connective tissue disease-associated interstitial lung disease, pulmonary fibrosis due to sarcoidosis, and chronic drug-induced interstitial lung disease. In each of these diseases, the fibrosis becomes progressive in approximately 20% to 30% of cases, a condition referred to as progressive fibrosing interstitial lung disease (PF-ILD) [13,14]. The prognosis of PF-ILD has been reported to follow a similar pathology to that of fibrosis in the lungs derived from any originating disease, with an annual forced vital capacity (FVC) loss rate of approximately −190 mL/year [14]. IPF is reported to be chronically progressive, with a median survival of 3 to 5 years after diagnosis [1,2], and to have a similar course to progressive FILD [15].
Although the pathogenesis of FILD varies according to the causative disease, the changes that occur as fibrosis develops are reported to be similar. The mechanism of fibrosis is sustained injury to alveolar epithelial cells, and damage to microvascular endothelial cells and interstitium caused by chemical and immunological agents, such as smoking, drugs, autoimmune antibodies, or cells, which result in inflammation and abnormal wound healing in the pulmonary interstitium. Subsequently, fibroblasts, pericytes, epithelial cells, vascular endothelial cells, and fibrocytes are transformed into myofibroblasts, leading to fibrogenesis [3]. Myofibroblasts produce excess extracellular matrix and tissue remodeling occurs, and then the fibrotic lung tissue itself or hypoxia in the fibrotic area leads to the upregulated production of pro-fibrotic cytokines and activation of myofibroblasts, resulting in a loop-like progression of fibrosis [3,16,17,18,19,20,21]. This process involves pro-fibrotic cytokines such as transforming growth factor-β (TGF-β), platelet-derived growth factor (PDGF), and fibroblast growth factor (FGF) [22,23] and causes airway epithelial cell injury, differentiation into myofibroblasts, epithelial–mesenchymal transition, and the tissue deposition of extracellular matrix, which leads to fibrotic formation [24,25]. The fibrosis process also involves abnormalities in lipid metabolism and the disruption of the vascular network [21,25,26].
Although FILD is treated at its source [27], steroid therapy, the most widely used anti-inflammatory treatment in FILD, may lack efficacy [28,29]. Anti-fibrotic drugs such as nintedanib [30] and pirfenidone [31,32] have been used in the hope of suppressing fibrosis progression, but improvement in disease status is often difficult. In cases where patients are refractory to drug therapy, lung transplantation may be considered, but it has many drawbacks, including age restrictions, the limited number of donors and post-transplant immunosuppressive medications, and poor long-term prognosis [33].
As described above, once fibrosis occurs in FILD, it can lead to progressive fibrosis. Given that FILD is often difficult to treat with current drug therapies and treatment options are limited, the development of novel treatment strategies is highly desirable.
1.2. An Overview of iPSCs
iPSCs are artificially induced pluripotent stem cells that were initially generated from mouse cells in 2006 and then from human cells in 2007 [34,35]. Pluripotent stem cells are induced by introducing transcriptional factors, including OCT3/4, SOX2, KLF4, and C-MYC, into somatic cells [35]. iPSCs can differentiate into a variety of cell types, and if they are generated from donor cells, pluripotent stem cells can be generated with a lower risk of immune rejection. However, transplanting iPSCs in an undifferentiated state leads to teratoma formation. In addition, this may induce an immune response against them. Therefore, the transplantation of undifferentiated iPSCs is not practical in clinical applications [36,37,38,39]. In response to teratoma issues, some researchers have modified the reprogramming factor C-MYC to L-MYC, which does not promote tumorigenesis [40], and have devised methods to introduce reprogramming factors without integrating them into the genome (e.g., the plasmid method, or adenovirus method that transiently expresses reprogramming genes) [41,42].
The clinical use of iPSC-derived differentiated cells is being explored in various diseases and applications, including age-related macular degeneration [5], Parkinson’s disease [43,44], platelet production [45,46], and heart failure [47,48]. iPSCs have also been used in reproductive medicine, and it has been reported that functional oocytes have been successfully generated from male mice-derived iPSCs [49]. In addition, patient-specific iPSCs have potential as a strategy for drug screening [10,50]. iPSCs have been reported to have potential efficacy in FILD due to their anti-fibrotic properties [6,7,8,9], but as mentioned above, the transplantation of iPSCs themselves is difficult to utilize due to the teratoma formation. However, it is important to investigate the application of iPSC-derived cells for the treatment of FILD.
2. Current Status of Cell-Based Therapies for FILD
Over the years, several cells have been reported to act protectively against FILD [6,7,8,9,51,52,53,54]. For example, a study in 2003 by Ortiz et al. showed that when bone marrow-derived mesenchymal stem cells (BM-MSCs) were intravascularly injected into a mouse model of bleomycin (BLM)-induced interstitial lung disease, they migrated to the damaged lungs and transformed into alveolar epithelial type II (AT2)-like cells, leading to a reduction in inflammation and collagen deposition in lung tissue [51]. In addition to BM-MSCs [51,52,53,54,55,56,57,58,59,60,61,62,63,64], cell-based therapy with the following has also been reported to have benefits for pulmonary fibrosis: iPSCs [6,7,8,9], embryonic stem cells (ESCs) [65], resident lung MSCs [66], placenta-derived cell mixture [67,68], amnion MSCs [69], umbilical cord MSCs [70], amniotic fluid stem cells [71], adipose-derived MSCs (AD-MSCs) [72,73,74,75], hematopoietic stem cells (HSCs) [76], AT2 [77,78], lung spheroid cells [79], and prominin-1/CD133(+) epithelial progenitor cells (PEPCEs) [80]. The various cell-based therapies have a similar mechanism of anti-fibrotic action in the lung, with stem cells accumulating at inflammatory sites and reducing pro-fibrotic cytokines, actions that are thought to exert an anti-fibrotic effect. Furthermore, these cell-based therapies have also been reported to be useful in combination with existing anti-fibrotic therapies [64] and thus hold promise as novel therapies that can be added to current treatments.
In this section, we will review the current status of cell-based therapies for FILD (summarized in Figure 1 and Table 1).
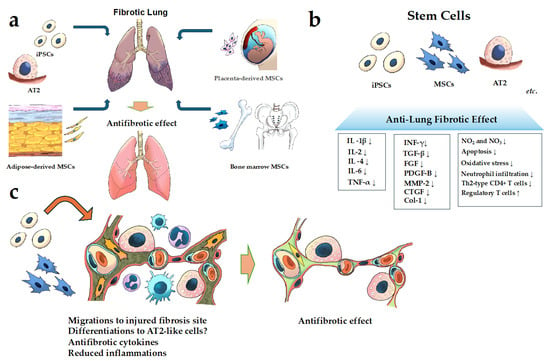
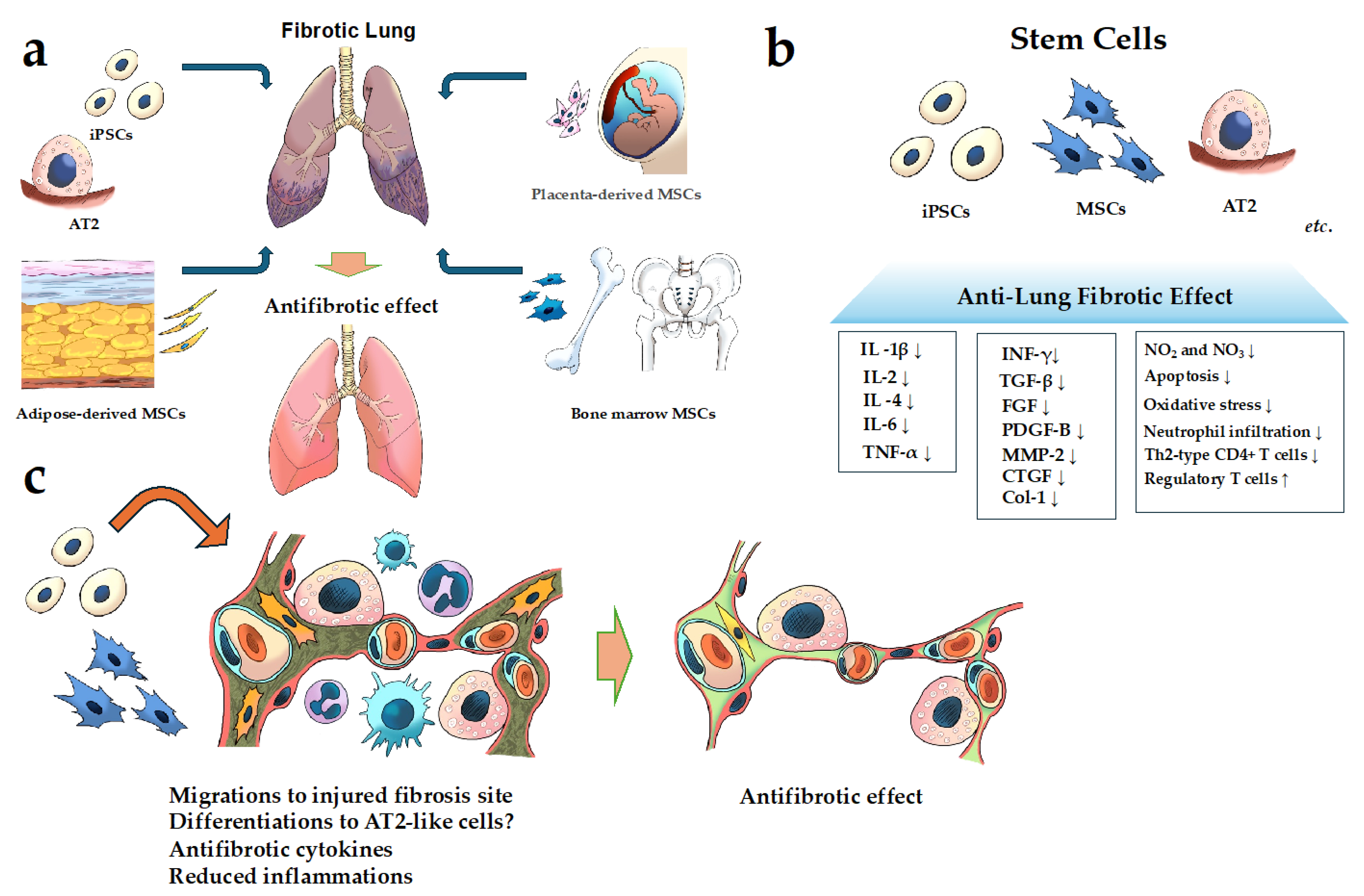
Figure 1.
Schematic of cell-based therapy for FILD. (a) Examples of the cells used in cell-based therapy in pulmonary fibrosis; (b,c) the mechanism of the action of the anti-fibrotic effect of stem cells for pulmonary fibrosis. It has been reported that MSCs, iPSCs, and AT2 have anti-fibrotic effects through several mechanisms described in (b,c). Abbreviations: AT2, alveolar epithelial type II; Coll, collagen; CTGF, connective tissue growth factor; IL, interleukin; FGF, fibroblast growth factor; INF-γ, interferon gamma; MMP, matrix metalloproteinase; MSCs, mesenchymal stem cells; PDGF, platelet-derived growth factor; TGF-β, transforming growth factor-beta 1; Th2-type, T helper 2 type; TNF-α, tumor necrosis factor-α. (↑; indicates upregulation or increase. ↓; indicates downregulation or decrease.)

Table 1.
Preclinical studies on cell-based therapy for fibrosing interstitial lung diseases with stem cells. Summarizes the progress to date in cell-based therapy for FILD.
2.1. FILD Treatment with MSCs
MSCs are the most-studied stem cells in cell-based therapy for pulmonary fibrosis. An animal study reported that myelosuppression with busulfan exacerbates fibrosis in a mouse model of BLM-induced interstitial lung disease, suggesting that BM-MSCs may be involved in defense mechanisms against pulmonary fibrosis as a physiological effect [55]. Therefore, BM-MSCs, and other stem cells, may be useful against pulmonary fibrosis.
The anti-fibrotic mechanisms of the action of MSCs are thought to be influenced by factors such as transformation into AT2-like cells (AT2 is involved in alveolar epithelial cell regeneration) [51,58], reduced TGF-β [54,57,61,63,64,69,74,75], reduced FGF [57], reduced PDGF [57], reduced connective tissue growth factor (CTGF) [59,61], the involvement of matrix metalloproteinases (MMPs; with varying increases and decreases, depending on the literature and subtype) [61,69,73,81], reduced inflammatory cytokines (tumor necrosis factor alpha [TNF-α], interleukin [IL]-1α, IL-2, IL-1β, IL-4, and interferon gamma) [55,56,60], increased migration factors (granulocyte colony-stimulating factor [G-CSF] and granulocyte-macrophage colony-stimulating factor [GM-CSF]) [55], the involvement of hepatocyte growth factor (HGF; HGF is responsible for the restoration of alveolar epithelial cells) [53,60], the involvement of stanniocalcin-1 (a mitochondria-related hormone that improves the cell survival) [57], and the involvement of nitric oxide metabolites (the downregulation of NO2 and NO3, which are thought to exert pro-fibrotic effects) [54]. Further details are provided in Table 1.
Another important issue for stem cell therapy is to find ways to enhance the anti-fibrotic efficacy of cell therapy, and various methods have been investigated with MSCs. One study reported that hypoxic exposure may enhance the anti-fibrotic effect of MSCs by increasing their anti-apoptotic and anti-oxidant factors [59], which can be considered as a potential mechanism for clinical application. Furthermore, culturing MSCs in serum-free media was found to enhance their anti-fibrotic effect, suggesting that the choice of culture medium is also an important aspect to consider [63]. In addition, efforts have been made to enhance the anti-fibrotic effect of MSCs by modifying the molecular biology of the stem cells, and one study investigated the anti-fibrotic effect of MSCs overexpressing miRNAs [62]. In that study, let-7d, a microRNA (miRNA) with anti-fibrotic activity, was overexpressed in MSCs and administered on day 7 after the administration of BLM, which resulted in a faster recovery of weight loss and reduced Col-1 expression, but did not lead to an apparent improvement in fibrosis [62]. Similarly, a study reported an enhanced anti-fibrotic effect of BM-MSCs by the overexpression of HGF [53]. Oncostatin M, which is involved in fibroblast proliferation, has been reported to be elevated in bronchoalveolar lavage fluid from patients with pulmonary fibrosis, and pretreatment with oncostatin M was found to further suppress fibrosis in BLM-induced mouse models [61]. Thus, in addition to the administration of MSCs, exploring methods to enhance their function is also an important issue in cell therapy.
In addition, there is a possibility that MSCs have different effects depending on the age of the donor. In BLM-induced interstitial lung disease mouse models, pulmonary fibrosis showed improvement with AD-MSCs derived from young donor mice (4 months) but not with AD-MSCs derived from aged donor mice (22 months); the authors speculated that the difference in the anti-fibrotic effect of the AD-MSCs was due to the lower fibrotic MMP-2 levels in AD-MSCs from young mice compared with those from old donors [73]. When considering the clinical applications of MSCs, it is necessary to fully assess the origin of the cells and the number of passages.
As described above, many studies have evaluated the use of MSCs in pulmonary fibrosis, and many innovative ways to enhance their efficacy are being investigated.
2.2. FILD Treatment with iPSCs
The administration of iPSCs has also been investigated for the treatment of interstitial lung disease [6,7,8,9]. In the model of BLM-induced interstitial lung disease, iPSCs, similar to MSCs, were shown to accumulate in the lung, the site of inflammation, and to have an anti-fibrotic effect [7,8]. Other stem cells, including MSCs and those differentiated from iPSCs into AT2-like cells, also accumulate in the lung at the site of inflammation [8], suggesting that accumulation at the site of inflammation is a cellular property of the stem cells themselves. With respect to the lung, a limited number of reports have described the type of cells that differentiate after the administration of iPSCs. Similarly, relatively few detailed studies have examined the presence of MSCs after cell transplantation, but iPSCs are more pluripotent than MSCs, so they may be more relevant when considering the clinical application of stem cells.
The mechanism of the anti-fibrotic effect of iPSCs is similar to that of other stem cells and is thought to be due to a reduction in inflammatory cytokines such as IL-1, IL-2, IL-10, TNF-α, and monocyte chemotactic protein 1 (MCP1) [7], reduced TGF-β [9], and increased anti-fibrotic chemokine, interferon-gamma-induced protein 10 (IP10) [7]. There have been reports of cytokine changes in the conditioned medium of iPSCs similar to those in iPSCs [7,9], and the factors and pathways by which these changes are induced are the subject of further investigation. Research has found that the TGF-β1/Smad2/3 cascade [9] and the Wnt/β-catenin cascade are involved as signaling pathways in pulmonary fibrosis [6], and the signaling involved in the cell-based therapy of iPSCs for FILD is also becoming increasingly clear. Furthermore, one study showed a significant improvement in survival when iPSCs were administered to BLM-induced mouse models, indicating their possible suitability for clinical applications [7]. The iPSCs used in this study were generated without c-MYC to reduce the tumorigenic risk [83], and the method was aimed at clinical application.
iPSCs have also been reported to be useful in various lung diseases other than FILD. For example, they were found to protect against ventilator-induced lung injury via the nuclear factor-κB pathway [84], against hyperoxia-augmented ventilator-induced lung injury via the Src-dependent signaling pathway [85], against acute lung injury induced by ischemia–reperfusion via the suppression of high-mobility group box-1 [86], an ameliorating effect on hyperoxia-induced lung injury by reducing inflammatory cytokines [87], and to have a protective effect in endotoxin-induced acute lung injury by improving nuclear factor-κB activity and the accumulation of neutrophils [88]. Only animal studies using iPSCs themselves have been reported so far.
The above outlines the use of iPSCs in cell-based therapy for FILD. Although fewer studies have been performed with iPSCs than with MSCs, iPSCs appear to exhibit similar anti-fibrotic effects. However, as mentioned above, clinical application using iPSCs themselves is difficult due to tumor formation.
2.3. FILD Treatment with Other Cells
Thus far, we have outlined various studies on cell-based therapy involving MSCs and iPSCs. As previously noted, other stem/progenitor cells have also been reported to have anti-fibrotic properties. For example, AT2 cells were found to act as progenitor cells in the adult lung and to contribute to alveolar repair in the event of lung injury [89] and the intratracheal administration of AT2 in BLM-induced interstitial lung disease mouse models localized to the lung, the site of injury, and suppressed lung fibrosis [77,78].
HSCs have also been studied in the treatment of FILD [76], and the administration of HSCs overexpressing keratinocyte growth factor to BLM-induced pulmonary fibrosis mouse models suppressed fibrosis by reducing TNF-α, CCL-2, and CCL-9 and increasing surfactant protein C-positive cells (AT2 cells) [76].
Cell therapy has also been performed with PEPCs and has shown anti-fibrotic activity against BLM-induced pulmonary fibrosis in mouse models. PEPCs, which are thought to be derived from the bone marrow, localize to the lung, and are thought to have anti-fibrotic effects, thereby differentiating into AT2 and upregulating inducible nitric oxide synthase [80]. Other reports suggest that lung spheroids (formed by MSC, alveolar epithelial cells type I [AT1], AT2, and club cells) also suppress fibrosis [79].
Some studies have reported on the use of differentiated cells derived from iPSCs [8] and ESCs [65] in models of interstitial lung disease, where the intratracheal administration of differentiated AT1 and AT2 derived from iPSCs in BLM-induced mouse models resulted in the suppression of fibrosis [8]. These results showed a significant improvement in the differentiated cell-treated group compared with undifferentiated iPSCs, suggesting the potential use of iPSC-derived differentiated cells for cell therapy [8]. In yet another report, MSCs differentiated from ESCs were studied in pulmonary fibrosis and showed the suppression of fibrosis [82]. The MSCs used in this study were developed by a pharmaceutical company and are already being tested in clinical research on COVID-19 [90].
As described above, various types of stem/progenitor cells other than MSCs and iPSCs are being studied as cell-based therapies for the treatment of FILD. Despite the fact that many types of stem cells have been reported to have protective effects against FILD, to our knowledge no studies have compared the anti-fibrotic effects of each stem cell type on a large scale. Therefore, when considering clinical applications, the selection of stem cells to be used may also be relevant.
2.4. Clinical Trials
Clinical research has been performed in IPF, the most common type of FILD, with AD-MSCs stromal vascular fraction [91,92], placental MSCs [93], AT2 [89], and BM-MSCs [94,95,96]. Table 2 summarizes the clinical trials performed in recent years in FILD (or IPF).
Table 2.
Clinical trials on cell-based therapy for fibrosing interstitial lung diseases with stem cells.
Between 2013 and 2018, phase I clinical trials were performed primarily to confirm safety. The studies ranged in duration from 4 weeks to 2 years and evaluated adverse events, mortality, progression-free survival, lung function, and exercise capacity. None of the studies showed any apparent adverse events, and the 2-year median survival and disease progression were comparable to the epidemiologic data [92].
Subsequently, a randomized phase I/IIA clinical trial with allogeneic human BM-MSCs in IPF patients was reported in 2020, the first of its kind to compare BM-MSC treatment with placebo in IPF patients [96]. On the basis of previous reports, this study used high-dose allogeneic BM-MSCs (2 × 108 cells) [95], and it assessed safety, tolerability, and efficacy as the endpoints [96]. The study found no substantial differences in significant adverse events or mortality between the two groups and showed efficacy in the BM-MSC-treated group in the 6 min walk distance (at 13 weeks), diffusing capacity of the lung for carbon monoxide (at 26 weeks), and FVC (at 39 weeks), and FVC at 52 weeks was higher than the baseline FVC [96]. Considering that the goal of FILD treatment with compounds such as pirfenidone and nintedanib is to curb the loss in FVC, the finding that cell therapy with MSCs also resulted in an increase in FVC was a decisive step forward from previous therapies. This study concluded that therapy with high doses of allogeneic MSCs was a safe and promising method to reduce disease progression [96].
As described above, phase II clinical trials have investigated cell therapies for IPF, and although they have collected comparative data only in a small number of cases, they have demonstrated the usefulness of BM-MSCs. Despite the limited number of studies, BM-MSCs are currently considered to be an effective candidate for cell-based therapies.
3. Points of Concern with Recent Cell-Based Therapy
As mentioned above, MSCs can be derived from a variety of organs, and AD-MSCs in particular have a high proliferative capacity and well-established culture techniques. However, the number of proliferations is limited compared to iPSCs. In other words, there is a potential risk of supply shortages. In addition, the efficacy of AD-MSCs may vary depending on the age of the donor from which they are derived [73]. Therefore, it is not simply a question of whether any AD-MSCs are acceptable. This suggests that it is difficult to maintain the homogeneity of quality.
BM-MSCs, which have been used in many clinical trials for FILD to date, are widely used and there are also concerns about their stable supply due to the limited number of donors with this invasive procedure. Placental MSCs and AT2 have similar problems, making it difficult to obtain enough cells for clinical application.
Because of these problems, iPSCs with high self-renewal potential may be a useful option as a cell source for FILD cell-based therapy. The use of iPSCs may solve the problems of consistent quality and stable supply associated with the use of MSCs. Previous reports about cell-based therapy have also highlighted the importance of product quality [97].
4. The Problems of Using iPSCs Themselves to Treat FILD
Many papers have reported that stem cells, including iPSCs, exhibit anti-fibrotic effects in the lung, and some MSCs have been found to be useful in clinical applications. Although the usefulness of iPSCs in terms of their anti-fibrotic effects has been reported, clinical applications have not yet been developed. As described above, there are several challenges to the clinical application of iPSCs themselves, particularly concerns about tumorigenesis [38] and the difficulty of accurately assessing differentiation after the administration of iPSCs. Although there is evidence that iPSCs transplanted for FILD accumulate in the lung, the site of inflammation [8], they are systemically disseminated when administered intravascularly. Because pluripotent stem cells such as iPSCs can be induced to differentiate into various cell types depending on the stimuli of the environment in which they are seeded, the administration of undifferentiated iPSCs is virtually infeasible as a clinical application unless their differentiation is assessed after administration. For example, if iPSCs inadvertently differentiate into hematopoietic stem cells, there may be a risk of future graft-versus-host disease. For this reason, clinical applications that use iPSCs themselves will be difficult to develop unless the cells demonstrate greater efficacy than MSCs or other stem cells with safety.
Although there is a paucity of the literature comparing the anti-fibrotic effects of iPSCs and differentiated cells, some studies have compared the anti-fibrotic effects of iPSCs and iPSC-derived AT2-like cells and found stronger anti-fibrotic effects in the latter [8], making the use of differentiated cells derived from iPSCs an option to be considered.
5. Possibility for iPSC-Derived Cell-Based FILD Therapy
Despite the problems associated with the clinical application of iPSCs themselves, these derived cells have advantages in cell-based therapy for the treatment of FILD. As mentioned previously, the stockpiling of various HLAs has begun in Japan, and the use of matching HLAs has the potential to complement not only the anti-inflammatory but also the functional effect of stem cell-based therapy [12]. MSCs and AT2, which have shown anti-fibrotic effects, have already been reported in the methods to differentiate from iPSCs [98,99]. Therefore, it is now conceivable to target drug development with these iPSC-derived cells with matched HLA.
As mentioned above, the drug supply for current cell-based therapies is dependent on the number of donors, which is a potential risk for stable supply. The proliferative capacity of iPSCs is not expected to decrease with culture passaging, unlike MSCs, which have been more extensively studied for cell-based therapy in pulmonary fibrosis. The use of iPSC-derived cells may also reduce the risk of quality variation between donors for the derivation of other stem cells (e.g., MSCs or AT2) and allow for a more stable product supply. In addition, both the selection of the source cells that show the strongest anti-fibrotic effect in FILD and the selection of the differentiated cells in the generation from iPSCs can be considered. This, in turn, could lead to the creation of cell products with greater anti-fibrotic activity than existing MSCs and AT2 cell therapies.
In addition, the clinical applications of iPSC-derived cells have already been underway [5], and iPSC-derived cells will not be an ethical or technical limitation. The use of differentiated cells derived from iPSCs is expected to reduce the risk of tumor formation, which has been a concern with iPSCs.
In light of the above, although there are difficulties in applying iPSCs themselves in cell therapy, if cells with high anti-fibrotic activity can be generated by further studying the origin of iPSCs and differentiated cells, such cells may be found to have an anti-fibrotic effect that is superior to that of the existing cell therapies (Figure 2).
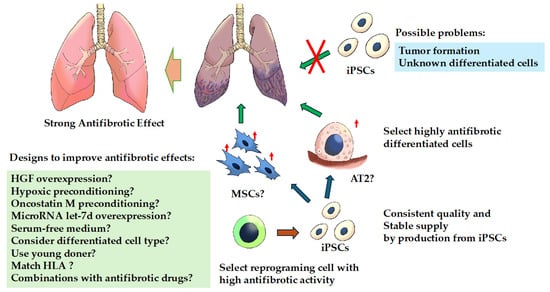
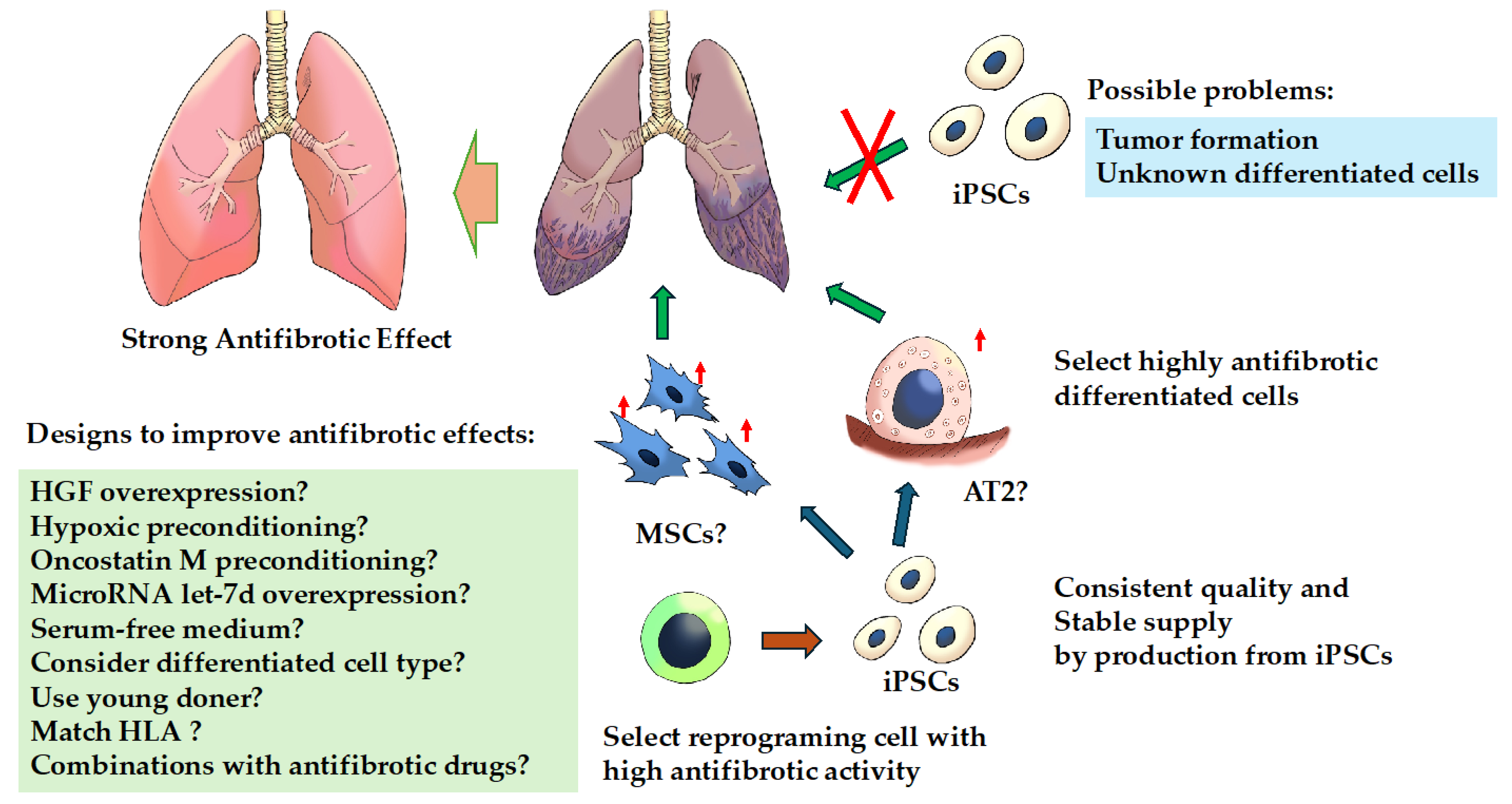
Figure 2.
Treatment strategies for fibrosing interstitial lung diseases with induced pluripotent stem cells. Cell-based therapy with induced pluripotent stem cells (iPSCs) per se is likely to be difficult for several reasons. There is a possibility that effective cell-based therapy can be achieved by inducing iPSCs from cells with efficient anti-fibrotic activity and differentiating them into cells with high anti-fibrotic activity because iPSCs may retain the characteristics of the parent cells from which they were generated. Abbreviations: AT2, alveolar epithelial type II; HGF, hepatocyte growth factor; iPSCs, induced pluripotent stem cells; MSCs, mesenchymal stem cells. ↑: indicates enhanced.
6. Proposal for iPSC-Derived Cell Type for FILD Treatment
The properties of iPSCs may differ depending on their origin, and in particular, low-passage-generated iPSCs have been reported to exhibit DNA methylation features similar to those of the somatic cells from which they were derived [100]. For example, such changes may facilitate differentiation into the derived cells and have indeed been used in animal experiments [101]. This means that when iPSCs are reprogrammed, the epigenetic changes are similar to the cells from which they are derived, and they are more likely to differentiate into their original cells. It is therefore desirable, in terms of differentiation efficiency, that the post-differentiated cells and the cells to be reprogrammed are identical.
MSCs have been reported to differ in gene expression depending on the tissue or organ from which they are harvested [102] in a study comparing umbilical cord blood-derived neonatal unrestricted somatic stem cells (USSCs), BM-MSCs, and AD-MSCs. USSCs showed gene expression related to neurogenesis, BM-MSCs showed a tendency to differentiate into mesoderm and ectoderm, and AD-MSCs were highly enriched in immune-related genes [102]. Because of these differences, the reprogramming cell source should also be taken into account when MSCs are to be considered as differentiated cells. If immunomodulatory effects are expected, it may be possible to produce cell products with high anti-fibrotic effects by using AD-MSCs as reprogramming cells, then generating iPSCs and differentiating them into AD-MSC-like cells. Other studies have reported that AD-MSCs secrete more HGF (responsible for the restoration of alveolar epithelial cells) and VEGF in the low serum culture system, and these may have a higher regenerative capacity and immunoregulatory function than BM-MSCs [103,104]. While many previous reports have used BM-MSCs, AD-MSCs may be more suitable. (↑; indicates enhanced.)
There are reports that both AD-MSCs and BM-MSCs are similar in terms of phenotype and differentiation [105], and it would be difficult to classify the molecular biology of the two. For example, it has been reported that the cytokine production and the gene expression involved in initiation vary with passaging. This further complicates the distinction between the two MScs. However, one report mentioned that the production of IL-6 by BM-MSCs is low in early passages, whereas AD-MSCs produce high levels, which is a possibility to be specific. It may be a result of being influenced by the location of the adipose tissue harvested, and further studies are needed to differentiate them. Despite these differences, both AD-MSCs and BM-MSCs are effective iPSC-derived cell options because they exert anti-fibrotic effects through similar mechanisms. AT2 has also been considered as a target for the differentiation of iPSCs. Although both MSCs and AT2 have been studied in clinical trials [92,95,96], there are no comparative studies between them, including basic studies, and future studies are needed.
At present, MSCs are most likely to be considered, given the abundance of previous reports and clinical trials on FILDs, and AD-MSCs, which have been reported to secrete high levels of HGF, are recommended as differentiated cells.
7. Limitations for iPSC-Derived Cell
The exact mechanism by which the differentiated cells exert their anti-inflammatory effects is not fully understood. Many studies have shown that these cells accumulate at the sites of inflammation and exert their anti-inflammatory effects, but there are some points to consider.
The first concern is the difficulty in assessing cell differentiation after the cell products have been administered. MSCs have “multipotency” to differentiate into multiple cell types, which is different from the “pluripotency” characteristic of iPSCs. However, for example, several reports have shown that administrated BM-MSCs were differentiated into AT2-like cells [51,58]. As these results have been scarce in recent years, the frequency of differentiation into AT2-like cells may not be high or strongly involved in anti-fibrotic effects. However, the fact that administered cells differentiate is certain, and assessing the distribution of all administered cells is difficult.
Another concern is that it is not fully understood whether the cells are eliminated or whether they differentiate and survive after administration at the site of inflammation. When transplanting human-derived MSCs into mice, engraftment varies depending on the immunodeficient condition of the mouse, thus rejection must be considered [52]. MSCs generally do not express MHC class II and therefore have low antigenicity and are not eliminated in the short term (48 h of INF-γ exposure upregulated MHC class II in MSC, but no lymphocyte alloreactivity was seen) [106]. On the other hand, long-term course and MSC-derived cells may be eliminated in the case of MHC-mismatched cell transplantation [107,108]. However, it does not suggest that the anti-fibrotic effect is lost with MSC rejection after a long period of administration. Human BM-MSCs showed anti-fibrotic effects even when transplanted into normal immunocompetent C57BL/6J BLM mouse models [57]. The MSCs are unlikely to be viable in the long term, and if they are thought to act in the short term then they may be eliminated. In other words, it is thought that an anti-inflammatory effect could be expected in the lung if it acts even for short periods during high levels of inflammation. As another example, bone marrow-derived “allogeneic” multipotent adult progenitor cells (invimestrocel) have been reported to potentially improve 180-day survival in acute respiratory distress syndrome (ARDS) [109]. Whether the transplanted cells engraft is an important question, but the initial anti-inflammatory effects of stem/progenitor cells may lead to a better long-term outcome, even if they are rejected. Although the degree of antigenicity of this cell is unknown and whether it is eliminated or engrafted cannot be determined, an assessment of the health risk of long-term engraftment is also necessary.
8. Conclusions
At present, the clinical use of undifferentiated iPSCs is likely to be difficult for a number of reasons that need to be addressed before iPSCs themselves can be introduced as therapeutic agents in cell-based therapy for FILD. However, regenerative medicine products generated from differentiated cells derived from iPSCs appear to hold promise as innovative cell-based therapies in the future.
Author Contributions
Y.N. designed and wrote the manuscript. Y.S. and S.N. reviewed, edited and supervised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Japan Society for the Promotion of Science (JSPS) KAKENHI Grant (19K17644) to Y.N., GlaxoSmithKline, GSK Japan Research Grant 2021 awarded to Y.N. (AS2021A000200866), Dokkyo Medical University Investigator-Initiated Research Grant awarded to Y.N. (No. 2023-04), and The Japanese Respiratory Society Boehringer Ingelheim research program to Y.S. (grant number none, 2023).
Data Availability Statement
Not applicable.
Conflicts of Interest
Y.N. has received research grants from GlaxoSmithKline (2021).
References
- Lederer, D.J.; Martinez, F.J. Idiopathic Pulmonary Fibrosis. N. Engl. J. Med. 2018, 378, 1811–1823. [Google Scholar] [CrossRef] [PubMed]
- Natsuizaka, M.; Chiba, H.; Kuronuma, K.; Otsuka, M.; Kudo, K.; Mori, M.; Bando, M.; Sugiyama, Y.; Takahashi, H. Epidemiologic survey of Japanese patients with idiopathic pulmonary fibrosis and investigation of ethnic differences. Am. J. Respir. Crit. Care Med. 2014, 190, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Wijsenbeek, M.; Cottin, V. Spectrum of Fibrotic Lung Diseases. N. Engl. J. Med. 2020, 383, 958–968. [Google Scholar] [CrossRef] [PubMed]
- Laporta Hernandez, R.; Aguilar Perez, M.; Lázaro Carrasco, M.T.; Ussetti Gil, P. Lung Transplantation in Idiopathic Pulmonary Fibrosis. Med. Sci. 2018, 6, 68. [Google Scholar] [CrossRef] [PubMed]
- Mandai, M.; Watanabe, A.; Kurimoto, Y.; Hirami, Y.; Morinaga, C.; Daimon, T.; Fujihara, M.; Akimaru, H.; Sakai, N.; Shibata, Y.; et al. Autologous Induced Stem-Cell-Derived Retinal Cells for Macular Degeneration. N. Engl. J. Med. 2017, 376, 1038–1046. [Google Scholar] [CrossRef] [PubMed]
- Bayati, P.; Taherian, M.; Soleimani, M.; Farajifard, H.; Mojtabavi, N. Induced pluripotent stem cells modulate the Wnt pathway in the bleomycin-induced model of idiopathic pulmonary fibrosis. Stem Cell Res. Ther. 2023, 14, 343. [Google Scholar] [CrossRef] [PubMed]
- How, C.K.; Chien, Y.; Yang, K.Y.; Shih, H.C.; Juan, C.C.; Yang, Y.P.; Chiou, G.Y.; Huang, P.I.; Chang, Y.L.; Chen, L.K.; et al. Induced pluripotent stem cells mediate the release of interferon gamma-induced protein 10 and alleviate bleomycin-induced lung inflammation and fibrosis. Shock 2013, 39, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Ye, X.; Sun, R.; Matsumoto, Y.; Moriyama, M.; Asano, Y.; Ajioka, Y.; Saijo, Y. Differentiation of mouse induced pluripotent stem cells into alveolar epithelial cells in vitro for use in vivo. Stem Cells Transl. Med. 2014, 3, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; He, Z.; Gao, Y.; Zheng, R.; Zhang, X.; Zhao, L.; Tan, M. Induced Pluripotent Stem Cells Inhibit Bleomycin-Induced Pulmonary Fibrosis in Mice through Suppressing TGF-β1/Smad-Mediated Epithelial to Mesenchymal Transition. Front. Pharmacol. 2016, 7, 430. [Google Scholar] [CrossRef]
- Hosokawa, M.; Mikawa, R.; Hagiwara, A.; Okuno, Y.; Awaya, T.; Yamamoto, Y.; Takahashi, S.; Yamaki, H.; Osawa, M.; Setoguchi, Y.; et al. Cryptotanshinone is a candidate therapeutic agent for interstitial lung disease associated with a BRICHOS-domain mutation of SFTPC. iScience 2023, 26, 107731. [Google Scholar] [CrossRef]
- Saito, M.K.; Osawa, M.; Tsuchida, N.; Shiraishi, K.; Niwa, A.; Woltjen, K.; Asaka, I.; Ogata, K.; Ito, S.; Kobayashi, S.; et al. A disease-specific iPS cell resource for studying rare and intractable diseases. Inflamm. Regen. 2023, 43, 43. [Google Scholar] [CrossRef]
- Center for iPS Cell Research and Application K.U.C. iPS Cell Stock Project, HLA-Homozygous iPS Cells. Available online: https://www.cira-foundation.or.jp/e/research-institution/ips-stock-project/homozygous.html (accessed on 1 April 2024).
- Wijsenbeek, M.; Kreuter, M.; Olson, A.; Fischer, A.; Bendstrup, E.; Wells, C.D.; Denton, C.P.; Mounir, B.; Zouad-Lejour, L.; Quaresma, M.; et al. Progressive fibrosing interstitial lung diseases: Current practice in diagnosis and management. Curr. Med. Res. Opin. 2019, 35, 2015–2024. [Google Scholar] [CrossRef] [PubMed]
- Wells, A.U.; Flaherty, K.R.; Brown, K.K.; Inoue, Y.; Devaraj, A.; Richeldi, L.; Moua, T.; Crestani, B.; Wuyts, W.A.; Stowasser, S.; et al. Nintedanib in patients with progressive fibrosing interstitial lung diseases-subgroup analyses by interstitial lung disease diagnosis in the INBUILD trial: A randomised, double-blind, placebo-controlled, parallel-group trial. Lancet Respir. Med. 2020, 8, 453–460. [Google Scholar] [CrossRef]
- Brown, K.K.; Martinez, F.J.; Walsh, S.L.F.; Thannickal, V.J.; Prasse, A.; Schlenker-Herceg, R.; Goeldner, R.G.; Clerisme-Beaty, E.; Tetzlaff, K.; Cottin, V.; et al. The natural history of progressive fibrosing interstitial lung diseases. Eur. Respir. J. 2020, 55, 2000085. [Google Scholar] [CrossRef]
- Distler, J.H.W.; Györfi, A.H.; Ramanujam, M.; Whitfield, M.L.; Königshoff, M.; Lafyatis, R. Shared and distinct mechanisms of fibrosis. Nat. Rev. Rheumatol. 2019, 15, 705–730. [Google Scholar] [CrossRef]
- Winters, N.I.; Burman, A.; Kropski, J.A.; Blackwell, T.S. Epithelial Injury and Dysfunction in the Pathogenesis of Idiopathic PulmonaryFibrosis. Am. J. Med. Sci. 2019, 357, 374–378. [Google Scholar] [CrossRef]
- Wells, R.G. Tissue mechanics and fibrosis. Biochim. Biophys. Acta 2013, 1832, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Lokmic, Z.; Musyoka, J.; Hewitson, T.D.; Darby, I.A. Hypoxia and hypoxia signaling in tissue repair and fibrosis. Int. Rev. Cell Mol. Biol. 2012, 296, 139–185. [Google Scholar] [PubMed]
- Parker, M.W.; Rossi, D.; Peterson, M.; Smith, K.; Sikström, K.; White, E.S.; Connett, J.E.; Henke, C.A.; Larsson, O.; Bitterman, P.B. Fibrotic extracellular matrix activates a profibrotic positive feedback loop. J. Clin. Investig. 2014, 124, 1622–1635. [Google Scholar] [CrossRef]
- Nakamura, Y.; Shimizu, Y.; Fujimaki-Shiraishi, M.; Uchida, N.; Takemasa, A.; Niho, S. A Protective Effect of Pirfenidone in Lung Fibroblast-Endothelial Cell Network via Inhibition of Rho-Kinase Activity. Biomedicines 2023, 11, 2259. [Google Scholar] [CrossRef]
- Kelly, M.; Kolb, M.; Bonniaud, P.; Gauldie, J. Re-evaluation of fibrogenic cytokines in lung fibrosis. Curr. Pharm. Des. 2003, 9, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Vaillant, P.; Menard, O.; Vignaud, J.M.; Martinet, N.; Martinet, Y. The role of cytokines in human lung fibrosis. Monaldi Arch. Chest Dis. 1996, 51, 145–152. [Google Scholar]
- Fernandez, I.E.; Eickelberg, O. The impact of TGF-β on lung fibrosis: From targeting to biomarkers. Proc. Am. Thorac. Soc. 2012, 9, 111–116. [Google Scholar] [CrossRef]
- Nakamura, Y.; Shimizu, Y. Cellular and Molecular Control of Lipid Metabolism in Idiopathic Pulmonary Fibrosis: Clinical Application of the Lysophosphatidic Acid Pathway. Cells 2023, 12, 548. [Google Scholar] [CrossRef]
- Uchida, N.; Shimizu, Y.; Fujimaki, M.; Horibata, Y.; Nakamura, Y.; Horigane, Y.; Chibana, K.; Takemasa, A.; Sugimoto, H.; Niho, S. Metabolic changes induced by TGF-β1 via reduced expression of phosphatidylserine decarboxylase during myofibroblast transition. J. Clin. Biochem. Nutr. 2022, 70, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Johannson, K.A.; Chaudhuri, N.; Adegunsoye, A.; Wolters, P.J. Treatment of fibrotic interstitial lung disease: Current approaches and future directions. Lancet 2021, 398, 1450–1460. [Google Scholar] [CrossRef] [PubMed]
- Richeldi, L.; Davies, H.R.; Ferrara, G.; Franco, F. Corticosteroids for idiopathic pulmonary fibrosis. Cochrane Database Syst. Rev. 2003, 2003, cd002880. [Google Scholar] [CrossRef]
- Raghu, G.; Collard, H.R.; Egan, J.J.; Martinez, F.J.; Behr, J.; Brown, K.K.; Colby, T.V.; Cordier, J.F.; Flaherty, K.R.; Lasky, J.A.; et al. An official ATS/ERS/JRS/ALAT statement: Idiopathic pulmonary fibrosis: Evidence-based guidelines for diagnosis and management. Am. J. Respir. Crit. Care Med. 2011, 183, 788–824. [Google Scholar] [CrossRef]
- Flaherty, K.R.; Wells, A.U.; Cottin, V.; Devaraj, A.; Walsh, S.L.F.; Inoue, Y.; Richeldi, L.; Kolb, M.; Tetzlaff, K.; Stowasser, S.; et al. Nintedanib in Progressive Fibrosing Interstitial Lung Diseases. N. Engl. J. Med. 2019, 381, 1718–1727. [Google Scholar] [CrossRef]
- Behr, J.; Prasse, A.; Kreuter, M.; Johow, J.; Rabe, K.F.; Bonella, F.; Bonnet, R.; Grohe, C.; Held, M.; Wilkens, H.; et al. Pirfenidone in patients with progressive fibrotic interstitial lung diseases other than idiopathic pulmonary fibrosis (RELIEF): A double-blind, randomised, placebo-controlled, phase 2b trial. Lancet Respir. Med. 2021, 9, 476–486. [Google Scholar] [CrossRef]
- Maher, T.M.; Corte, T.J.; Fischer, A.; Kreuter, M.; Lederer, D.J.; Molina-Molina, M.; Axmann, J.; Kirchgaessler, K.U.; Samara, K.; Gilberg, F.; et al. Pirfenidone in patients with unclassifiable progressive fibrosing interstitial lung disease: A double-blind, randomised, placebo-controlled, phase 2 trial. Lancet Respir. Med. 2020, 8, 147–157. [Google Scholar] [CrossRef]
- Leong, S.W.; Bos, S.; Lordan, J.L.; Nair, A.; Fisher, A.J.; Meachery, G. Lung transplantation for interstitial lung disease: Evolution over three decades. BMJ Open Respir. Res. 2023, 10, e001387. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Zhang, Z.N.; Rong, Z.; Xu, Y. Immunogenicity of induced pluripotent stem cells. Nature 2011, 474, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Okita, K.; Nagata, N.; Yamanaka, S. Immunogenicity of induced pluripotent stem cells. Circ. Res. 2011, 109, 720–721. [Google Scholar] [CrossRef]
- Masuda, S. Risk of teratoma formation after transplantation of induced pluripotent stem cells. Chest 2012, 141, 1120–1121. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Kawai, H.; Tian, F.; Ohta, Y.; Abe, K. Tumorigenic development of induced pluripotent stem cells in ischemic mouse brain. Cell Transplant. 2011, 20, 883–891. [Google Scholar] [CrossRef]
- Nakagawa, M.; Takizawa, N.; Narita, M.; Ichisaka, T.; Yamanaka, S. Promotion of direct reprogramming by transformation-deficient Myc. Proc. Natl. Acad. Sci. USA 2010, 107, 14152–14157. [Google Scholar] [CrossRef]
- Okita, K.; Nakagawa, M.; Hyenjong, H.; Ichisaka, T.; Yamanaka, S. Generation of mouse induced pluripotent stem cells without viral vectors. Science 2008, 322, 949–953. [Google Scholar] [CrossRef]
- Stadtfeld, M.; Nagaya, M.; Utikal, J.; Weir, G.; Hochedlinger, K. Induced pluripotent stem cells generated without viral integration. Science 2008, 322, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, J. iPS cell-based therapy for Parkinson’s disease: A Kyoto trial. Regen. Ther. 2020, 13, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Morizane, A.; Doi, D.; Magotani, H.; Onoe, H.; Hayashi, T.; Mizuma, H.; Takara, S.; Takahashi, R.; Inoue, H.; et al. Human iPS cell-derived dopaminergic neurons function in a primate Parkinson’s disease model. Nature 2017, 548, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, N.; Eto, K. Platelet production from induced pluripotent stem cells. J. Thromb. Haemost. 2017, 15, 1717–1727. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, N.; Kanda, J.; Nakamura, S.; Kitano, T.; Hishizawa, M.; Kondo, T.; Shimizu, S.; Shigemasa, A.; Hirai, H.; Arai, Y.; et al. iPLAT1: The first-in-human clinical trial of iPSC-derived platelets as a phase 1 autologous transfusion study. Blood 2022, 140, 2398–2402. [Google Scholar] [CrossRef] [PubMed]
- A Prospective Observational Study of Induced Pluripoteint Stem Cell-Derived Cardiac Spheres Transplantation Extended Follow-up. UMIN-CTR Clinical Trial. 2022. Available online: https://center6.umin.ac.jp/cgi-open-bin/ctr_e/ctr_view.cgi?recptno=R000053954 (accessed on 1 April 2024).
- Samura, T.; Miyagawa, S.; Kawamura, T.; Fukushima, S.; Yokoyama, J.Y.; Takeda, M.; Harada, A.; Ohashi, F.; Sato-Nishiuchi, R.; Toyofuku, T.; et al. Laminin-221 Enhances Therapeutic Effects of Human-Induced Pluripotent Stem Cell-Derived 3-Dimensional Engineered Cardiac Tissue Transplantation in a Rat Ischemic Cardiomyopathy Model. J. Am. Heart Assoc. 2020, 9, e015841. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Hamazaki, N.; Hamada, N.; Nagamatsu, G.; Okamoto, I.; Ohta, H.; Nosaka, Y.; Ishikura, Y.; Kitajima, T.S.; Semba, Y.; et al. Generation of functional oocytes from male mice in vitro. Nature 2023, 615, 900–906. [Google Scholar] [CrossRef]
- Avior, Y.; Sagi, I.; Benvenisty, N. Pluripotent stem cells in disease modelling and drug discovery. Nat. Rev. Mol. Cell Biol. 2016, 17, 170–182. [Google Scholar] [CrossRef]
- Ortiz, L.A.; Gambelli, F.; McBride, C.; Gaupp, D.; Baddoo, M.; Kaminski, N.; Phinney, D.G. Mesenchymal stem cell engraftment in lung is enhanced in response to bleomycin exposure and ameliorates its fibrotic effects. Proc. Natl. Acad. Sci. USA 2003, 100, 8407–8411. [Google Scholar] [CrossRef] [PubMed]
- Liebler, J.M.; Lutzko, C.; Banfalvi, A.; Senadheera, D.; Aghamohammadi, N.; Crandall, E.D.; Borok, Z. Retention of human bone marrow-derived cells in murine lungs following bleomycin-induced lung injury. Am. J. Physiol. Lung Cell Mol. Physiol. 2008, 295, L285–L292. [Google Scholar] [CrossRef]
- Gazdhar, A.; Susuri, N.; Hostettler, K.; Gugger, M.; Knudsen, L.; Roth, M.; Ochs, M.; Geiser, T. HGF Expressing Stem Cells in Usual Interstitial Pneumonia Originate from the Bone Marrow and Are Antifibrotic. PLoS ONE 2013, 8, e65453. [Google Scholar] [CrossRef]
- Lee, S.H.; Jang, A.S.; Kim, Y.E.; Cha, J.Y.; Kim, T.H.; Jung, S.; Park, S.K.; Lee, Y.K.; Won, J.H.; Kim, Y.H.; et al. Modulation of cytokine and nitric oxide by mesenchymal stem cell transfer in lung injury/fibrosis. Respir. Res. 2010, 11, 16. [Google Scholar] [CrossRef]
- Rojas, M.; Xu, J.; Woods, C.R.; Mora, A.L.; Spears, W.; Roman, J.; Brigham, K.L. Bone marrow-derived mesenchymal stem cells in repair of the injured lung. Am. J. Respir. Cell Mol. Biol. 2005, 33, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, L.A.; Dutreil, M.; Fattman, C.; Pandey, A.C.; Torres, G.; Go, K.; Phinney, D.G. Interleukin 1 receptor antagonist mediates the antiinflammatory and antifibrotic effect of mesenchymal stem cells during lung injury. Proc. Natl. Acad. Sci. USA 2007, 104, 11002–11007. [Google Scholar] [CrossRef]
- Ono, M.; Ohkouchi, S.; Kanehira, M.; Tode, N.; Kobayashi, M.; Ebina, M.; Nukiwa, T.; Irokawa, T.; Ogawa, H.; Akaike, T.; et al. Mesenchymal stem cells correct inappropriate epithelial-mesenchyme relation in pulmonary fibrosis using stanniocalcin-1. Mol. Ther. 2015, 23, 549–560. [Google Scholar] [CrossRef]
- Huang, K.; Kang, X.; Wang, X.; Wu, S.; Xiao, J.; Li, Z.; Wu, X.; Zhang, W. Conversion of bone marrow mesenchymal stem cells into type II alveolar epithelial cells reduces pulmonary fibrosis by decreasing oxidative stress in rats. Mol. Med. Rep. 2015, 11, 1685–1692. [Google Scholar] [CrossRef]
- Lan, Y.W.; Choo, K.B.; Chen, C.M.; Hung, T.H.; Chen, Y.B.; Hsieh, C.H.; Kuo, H.P.; Chong, K.Y. Hypoxia-preconditioned mesenchymal stem cells attenuate bleomycin-induced pulmonary fibrosis. Stem Cell Res. Ther. 2015, 6, 97. [Google Scholar] [CrossRef]
- Cahill, E.F.; Kennelly, H.; Carty, F.; Mahon, B.P.; English, K. Hepatocyte Growth Factor Is Required for Mesenchymal Stromal Cell Protection Against Bleomycin-Induced Pulmonary Fibrosis. Stem Cells Transl. Med. 2016, 5, 1307–1318. [Google Scholar] [CrossRef] [PubMed]
- Lan, Y.W.; Theng, S.M.; Huang, T.T.; Choo, K.B.; Chen, C.M.; Kuo, H.P.; Chong, K.Y. Oncostatin M-Preconditioned Mesenchymal Stem Cells Alleviate Bleomycin-Induced Pulmonary Fibrosis Through Paracrine Effects of the Hepatocyte Growth Factor. Stem Cells Transl. Med. 2017, 6, 1006–1017. [Google Scholar] [CrossRef] [PubMed]
- Huleihel, L.; Sellares, J.; Cardenes, N.; Álvarez, D.; Faner, R.; Sakamoto, K.; Yu, G.; Kapetanaki, M.G.; Kaminski, N.; Rojas, M. Modified mesenchymal stem cells using miRNA transduction alter lung injury in a bleomycin model. Am. J. Physiol. Lung Cell Mol. Physiol. 2017, 313, L92–L103. [Google Scholar] [CrossRef]
- Takao, S.; Nakashima, T.; Masuda, T.; Namba, M.; Sakamoto, S.; Yamaguchi, K.; Horimasu, Y.; Miyamoto, S.; Iwamoto, H.; Fujitaka, K.; et al. Human bone marrow-derived mesenchymal stromal cells cultured in serum-free media demonstrate enhanced antifibrotic abilities via prolonged survival and robust regulatory T cell induction in murine bleomycin-induced pulmonary fibrosis. Stem Cell Res. Ther. 2021, 12, 506. [Google Scholar] [CrossRef] [PubMed]
- Gad, E.S.; Salama, A.A.A.; El-Shafie, M.F.; Arafa, H.M.M.; Abdelsalam, R.M.; Khattab, M. The Anti-fibrotic and Anti-inflammatory Potential of Bone Marrow-Derived Mesenchymal Stem Cells and Nintedanib in Bleomycin-Induced Lung Fibrosis in Rats. Inflammation 2020, 43, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, E.R.; Laflamme, M.A.; Papayannopoulou, T.; Kahn, M.; Murry, C.E.; Henderson, W.R., Jr. Human embryonic stem cells differentiated to lung lineage-specific cells ameliorate pulmonary fibrosis in a xenograft transplant mouse model. PLoS ONE 2012, 7, e33165. [Google Scholar] [CrossRef] [PubMed]
- Jun, D.; Garat, C.; West, J.; Thorn, N.; Chow, K.; Cleaver, T.; Sullivan, T.; Torchia, E.C.; Childs, C.; Shade, T.; et al. The pathology of bleomycin-induced fibrosis is associated with loss of resident lung mesenchymal stem cells that regulate effector T-cell proliferation. Stem Cells 2011, 29, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Cargnoni, A.; Gibelli, L.; Tosini, A.; Signoroni, P.B.; Nassuato, C.; Arienti, D.; Lombardi, G.; Albertini, A.; Wengler, G.S.; Parolini, O. Transplantation of allogeneic and xenogeneic placenta-derived cells reduces bleomycin-induced lung fibrosis. Cell Transplant. 2009, 18, 405–422. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Han, F.; Li, H.; Zhang, J.; Qiao, X.; Shi, J.; Yang, L.; Dong, J.; Luo, M.; Wei, J.; et al. Human placental mesenchymal stem cells of fetal origins-alleviated inflammation and fibrosis by attenuating MyD88 signaling in bleomycin-induced pulmonary fibrosis mice. Mol. Immunol. 2017, 90, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Moodley, Y.; Vaghjiani, V.; Chan, J.; Baltic, S.; Ryan, M.; Tchongue, J.; Samuel, C.S.; Murthi, P.; Parolini, O.; Manuelpillai, U. Anti-inflammatory effects of adult stem cells in sustained lung injury: A comparative study. PLoS ONE 2013, 8, e69299. [Google Scholar] [CrossRef] [PubMed]
- Moodley, Y.; Atienza, D.; Manuelpillai, U.; Samuel, C.S.; Tchongue, J.; Ilancheran, S.; Boyd, R.; Trounson, A. Human umbilical cord mesenchymal stem cells reduce fibrosis of bleomycin-induced lung injury. Am. J. Pathol. 2009, 175, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Garcia, O.; Carraro, G.; Turcatel, G.; Hall, M.; Sedrakyan, S.; Roche, T.; Buckley, S.; Driscoll, B.; Perin, L.; Warburton, D. Amniotic fluid stem cells inhibit the progression of bleomycin-induced pulmonary fibrosis via CCL2 modulation in bronchoalveolar lavage. PLoS ONE 2013, 8, e71679. [Google Scholar] [CrossRef]
- Kotani, T.; Masutani, R.; Suzuka, T.; Oda, K.; Makino, S.; Ii, M. Anti-inflammatory and anti-fibrotic effects of intravenous adipose-derived stem cell transplantation in a mouse model of bleomycin-induced interstitial pneumonia. Sci. Rep. 2017, 7, 14608. [Google Scholar] [CrossRef]
- Tashiro, J.; Elliot, S.J.; Gerth, D.J.; Xia, X.; Pereira-Simon, S.; Choi, R.; Catanuto, P.; Shahzeidi, S.; Toonkel, R.L.; Shah, R.H.; et al. Therapeutic benefits of young, but not old, adipose-derived mesenchymal stem cells in a chronic mouse model of bleomycin-induced pulmonary fibrosis. Transl. Res. 2015, 166, 554–567. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Lee, E.J.; Lee, S.Y.; Kim, J.H.; Shim, J.J.; Shin, C.; In, K.H.; Kang, K.H.; Uhm, C.S.; Kim, H.K.; et al. The effect of adipose stem cell therapy on pulmonary fibrosis induced by repetitive intratracheal bleomycin in mice. Exp. Lung Res. 2014, 40, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wu, J.; Yuan, R.; Li, Y.; Yang, Q.; Wu, B.; Zhai, X.; Wang, J.; Magalon, J.; Sabatier, F.; et al. Adipose-derived mesenchymal stem cell therapy for reverse bleomycin-induced experimental pulmonary fibrosis. Sci. Rep. 2023, 13, 13183. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, S.; Scotton, C.J.; McNulty, K.; Nye, E.; Stamp, G.; Laurent, G.; Bonnet, D.; Janes, S.M. Bone marrow stem cells expressing keratinocyte growth factor via an inducible lentivirus protects against bleomycin-induced pulmonary fibrosis. PLoS ONE 2009, 4, e8013. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Mollar, A.; Nacher, M.; Gay-Jordi, G.; Closa, D.; Xaubet, A.; Bulbena, O. Intratracheal transplantation of alveolar type II cells reverses bleomycin-induced lung fibrosis. Am. J. Respir. Crit. Care Med. 2007, 176, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Guillamat-Prats, R.; Gay-Jordi, G.; Xaubet, A.; Peinado, V.I.; Serrano-Mollar, A. Alveolar type II cell transplantation restores pulmonary surfactant protein levels in lung fibrosis. J. Heart Lung Transplant. 2014, 33, 758–765. [Google Scholar] [CrossRef] [PubMed]
- Cores, J.; Dinh, P.C.; Hensley, T.; Adler, K.B.; Lobo, L.J.; Cheng, K. A pre-investigational new drug study of lung spheroid cell therapy for treating pulmonary fibrosis. Stem Cells Transl. Med. 2020, 9, 786–798. [Google Scholar] [CrossRef]
- Germano, D.; Blyszczuk, P.; Valaperti, A.; Kania, G.; Dirnhofer, S.; Landmesser, U.; Lüscher, T.F.; Hunziker, L.; Zulewski, H.; Eriksson, U. Prominin-1/CD133+ lung epithelial progenitors protect from bleomycin-induced pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2009, 179, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.A.; Yeh, C.C.; Hsu, C.H.; Hsu, C.W.; Kuo, F.H.; Tsai, P.J.; Fu, Y.S. Reversal of Pulmonary Fibrosis: Human Umbilical Mesenchymal Stem Cells from Wharton’s Jelly versus Human-Adipose-Derived Mesenchymal Stem Cells. Int. J. Mol. Sci. 2023, 24, 6948. [Google Scholar] [CrossRef]
- Lee, H.; Jeong, O.Y.; Park, H.J.; Lee, S.L.; Bok, E.Y.; Kim, M.; Suh, Y.S.; Cheon, Y.H.; Kim, H.O.; Kim, S.; et al. Promising Therapeutic Effects of Embryonic Stem Cells-Origin Mesenchymal Stem Cells in Experimental Pulmonary Fibrosis Models: Immunomodulatory and Anti-Apoptotic Mechanisms. Immune Netw. 2023, 23, e45. [Google Scholar] [CrossRef]
- Li, H.Y.; Chien, Y.; Chen, Y.J.; Chen, S.F.; Chang, Y.L.; Chiang, C.H.; Jeng, S.Y.; Chang, C.M.; Wang, M.L.; Chen, L.K.; et al. Reprogramming induced pluripotent stem cells in the absence of c-Myc for differentiation into hepatocyte-like cells. Biomaterials 2011, 32, 5994–6005. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Li, L.F.; Yang, C.T.; Lu, K.H.; Huang, C.C.; Kao, K.C.; Chiou, S.H. Suppressing NF-κB and NKRF Pathways by Induced Pluripotent Stem Cell Therapy in Mice with Ventilator-Induced Lung Injury. PLoS ONE 2013, 8, e66760. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Li, L.F.; Fu, J.Y.; Kao, K.C.; Huang, C.C.; Chien, Y.; Liao, Y.W.; Chiou, S.H.; Chang, Y.L. Induced pluripotent stem cell therapy ameliorates hyperoxia-augmented ventilator-induced lung injury through suppressing the Src pathway. PLoS ONE 2014, 9, e109953. [Google Scholar] [CrossRef]
- Li, Y.; Wang, S.; Liu, J.; Li, X.; Lu, M.; Wang, X.; Ren, Y.; Li, X.; Xiang, M. Induced Pluripotent Stem Cells Attenuate Acute Lung Injury Induced by Ischemia Reperfusion via Suppressing the High Mobility Group Box-1. Dose Response 2020, 18, 1559325820969340. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, A.; Wanczyk, H.; Jensen, T.; Finck, C. Human induced pluripotent stem cells ameliorate hyperoxia-induced lung injury in a mouse model. Am. J. Transl. Res. 2020, 12, 292–307. [Google Scholar]
- Yang, K.Y.; Shih, H.C.; How, C.K.; Chen, C.Y.; Hsu, H.S.; Yang, C.W.; Lee, Y.C.; Perng, R.P.; Peng, C.H.; Li, H.Y.; et al. IV delivery of induced pluripotent stem cells attenuates endotoxin-induced acute lung injury in mice. Chest 2011, 140, 1243–1253. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Mollar, A.; Gay-Jordi, G.; Guillamat-Prats, R.; Closa, D.; Hernandez-Gonzalez, F.; Marin, P.; Burgos, F.; Martorell, J.; Sánchez, M.; Arguis, P.; et al. Safety and Tolerability of Alveolar Type II Cell Transplantation in Idiopathic Pulmonary Fibrosis. Chest 2016, 150, 533–543. [Google Scholar] [CrossRef]
- Karyana, M.; Djaharuddin, I.; Rif’ati, L.; Arif, M.; Choi, M.K.; Angginy, N.; Yoon, A.; Han, J.; Josh, F.; Arlinda, D.; et al. Safety of DW-MSC infusion in patients with low clinical risk COVID-19 infection: A randomized, double-blind, placebo-controlled trial. Stem Cell Res. Ther. 2022, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Tzouvelekis, A.; Paspaliaris, V.; Koliakos, G.; Ntolios, P.; Bouros, E.; Oikonomou, A.; Zissimopoulos, A.; Boussios, N.; Dardzinski, B.; Gritzalis, D.; et al. A prospective, non-randomized, no placebo-controlled, phase Ib clinical trial to study the safety of the adipose derived stromal cells-stromal vascular fraction in idiopathic pulmonary fibrosis. J. Transl. Med. 2013, 11, 171. [Google Scholar] [CrossRef]
- Ntolios, P.; Manoloudi, E.; Tzouvelekis, A.; Bouros, E.; Steiropoulos, P.; Anevlavis, S.; Bouros, D.; Froudarakis, M.E. Longitudinal outcomes of patients enrolled in a phase Ib clinical trial of the adipose-derived stromal cells-stromal vascular fraction in idiopathic pulmonary fibrosis. Clin. Respir. J. 2018, 12, 2084–2089. [Google Scholar] [CrossRef]
- Chambers, D.C.; Enever, D.; Ilic, N.; Sparks, L.; Whitelaw, K.; Ayres, J.; Yerkovich, S.T.; Khalil, D.; Atkinson, K.M.; Hopkins, P.M. A phase 1b study of placenta-derived mesenchymal stromal cells in patients with idiopathic pulmonary fibrosis. Respirology 2014, 19, 1013–1018. [Google Scholar] [CrossRef] [PubMed]
- Glassberg, M.K.; Minkiewicz, J.; Toonkel, R.L.; Simonet, E.S.; Rubio, G.A.; DiFede, D.; Shafazand, S.; Khan, A.; Pujol, M.V.; LaRussa, V.F.; et al. Allogeneic Human Mesenchymal Stem Cells in Patients With Idiopathic Pulmonary Fibrosis via Intravenous Delivery (AETHER): A Phase I Safety Clinical Trial. Chest 2017, 151, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Fishman, J.E.; Kim, G.J.; Kyeong, N.Y.; Goldin, J.G.; Glassberg, M.K. Intravenous stem cell dose and changes in quantitative lung fibrosis and DLCO in the AETHER trial: A pilot study. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 7568–7572. [Google Scholar] [PubMed]
- Averyanov, A.; Koroleva, I.; Konoplyannikov, M.; Revkova, V.; Lesnyak, V.; Kalsin, V.; Danilevskaya, O.; Nikitin, A.; Sotnikova, A.; Kotova, S.; et al. First-in-human high-cumulative-dose stem cell therapy in idiopathic pulmonary fibrosis with rapid lung function decline. Stem Cells Transl. Med. 2020, 9, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, D.; Takenaka-Ninagawa, N.; Motoike, S.; Kajiya, M.; Akaboshi, T.; Zhao, C.; Shibata, M.; Senda, S.; Toyooka, Y.; Sakurai, H.; et al. Induction of functional xeno-free MSCs from human iPSCs via a neural crest cell lineage. NPJ Regen. Med. 2022, 7, 47. [Google Scholar] [CrossRef]
- Gotoh, S.; Ito, I.; Nagasaki, T.; Yamamoto, Y.; Konishi, S.; Korogi, Y.; Matsumoto, H.; Muro, S.; Hirai, T.; Funato, M.; et al. Generation of alveolar epithelial spheroids via isolated progenitor cells from human pluripotent stem cells. Stem Cell Rep. 2014, 3, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Doi, A.; Wen, B.; Ng, K.; Zhao, R.; Cahan, P.; Kim, J.; Aryee, M.J.; Ji, H.; Ehrlich, L.I.; et al. Epigenetic memory in induced pluripotent stem cells. Nature 2010, 467, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, C.; Murakami, Y.; Ishii, E.; Fujita, H.; Wakao, H. Reprogramming and redifferentiation of mucosal-associated invariant T cells reveal tumor inhibitory activity. eLife 2022, 11, e70848. [Google Scholar] [CrossRef] [PubMed]
- Jansen, B.J.; Gilissen, C.; Roelofs, H.; Schaap-Oziemlak, A.; Veltman, J.A.; Raymakers, R.A.; Jansen, J.H.; Kögler, G.; Figdor, C.G.; Torensma, R.; et al. Functional differences between mesenchymal stem cell populations are reflected by their transcriptome. Stem Cells Dev. 2010, 19, 481–490. [Google Scholar] [CrossRef]
- Iwashima, S.; Ozaki, T.; Maruyama, S.; Saka, Y.; Kobori, M.; Omae, K.; Yamaguchi, H.; Niimi, T.; Toriyama, K.; Kamei, Y.; et al. Novel culture system of mesenchymal stromal cells from human subcutaneous adipose tissue. Stem Cells Dev. 2009, 18, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Furuhashi, K.; Tsuboi, N.; Shimizu, A.; Katsuno, T.; Kim, H.; Saka, Y.; Ozaki, T.; Sado, Y.; Imai, E.; Matsuo, S.; et al. Serum-starved adipose-derived stromal cells ameliorate crescentic GN by promoting immunoregulatory macrophages. J. Am. Soc. Nephrol. 2013, 24, 587–603. [Google Scholar] [CrossRef] [PubMed]
- Kozlowska, U.; Krawczenko, A.; Futoma, K.; Jurek, T.; Rorat, M.; Patrzalek, D.; Klimczak, A. Similarities and differences between mesenchymal stem/progenitor cells derived from various human tissues. World J. Stem Cells 2019, 11, 347–374. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Tammik, C.; Rosendahl, K.; Zetterberg, E.; Ringdén, O. HLA expression and immunologic properties of differentiated and undifferentiated mesenchymal stem cells. Exp. Hematol. 2003, 31, 890–896. [Google Scholar] [CrossRef] [PubMed]
- Halm, D.; Leibig, N.; Martens, J.; Stark, G.B.; Groß, T.; Zimmermann, S.; Finkenzeller, G.; Lampert, F. Direct comparison of the immunogenicity of major histocompatibility complex-I and -II deficient mesenchymal stem cells in vivo. Biol. Chem. 2021, 402, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Eliopoulos, N.; Stagg, J.; Lejeune, L.; Pommey, S.; Galipeau, J. Allogeneic marrow stromal cells are immune rejected by MHC class I- and class II-mismatched recipient mice. Blood 2005, 106, 4057–4065. [Google Scholar] [CrossRef]
- Ichikado, K.; Kotani, T.; Kondoh, Y.; Imanaka, H.; Johkoh, T.; Fujimoto, K.; Nunomiya, S.; Kawayama, T.; Sawada, M.; Jenkins, E.; et al. Clinical efficacy and safety of multipotent adult progenitor cells (invimestrocel) for acute respiratory distress syndrome (ARDS) caused by pneumonia: A randomized, open-label, standard therapy-controlled, phase 2 multicenter study (ONE-BRIDGE). Stem Cell Res. Ther. 2023, 14, 217. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).