


Transcription Factor-Mediated Generation of Dopaminergic Neurons from Human iPSCs—A Comparison of Methods
 and
and {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. iPSC-Derived Dopaminergic Neuron Generation following Theka et al.
2.2.1. Overexpression of Transcription Factors Using Recombinant Lentiviruses
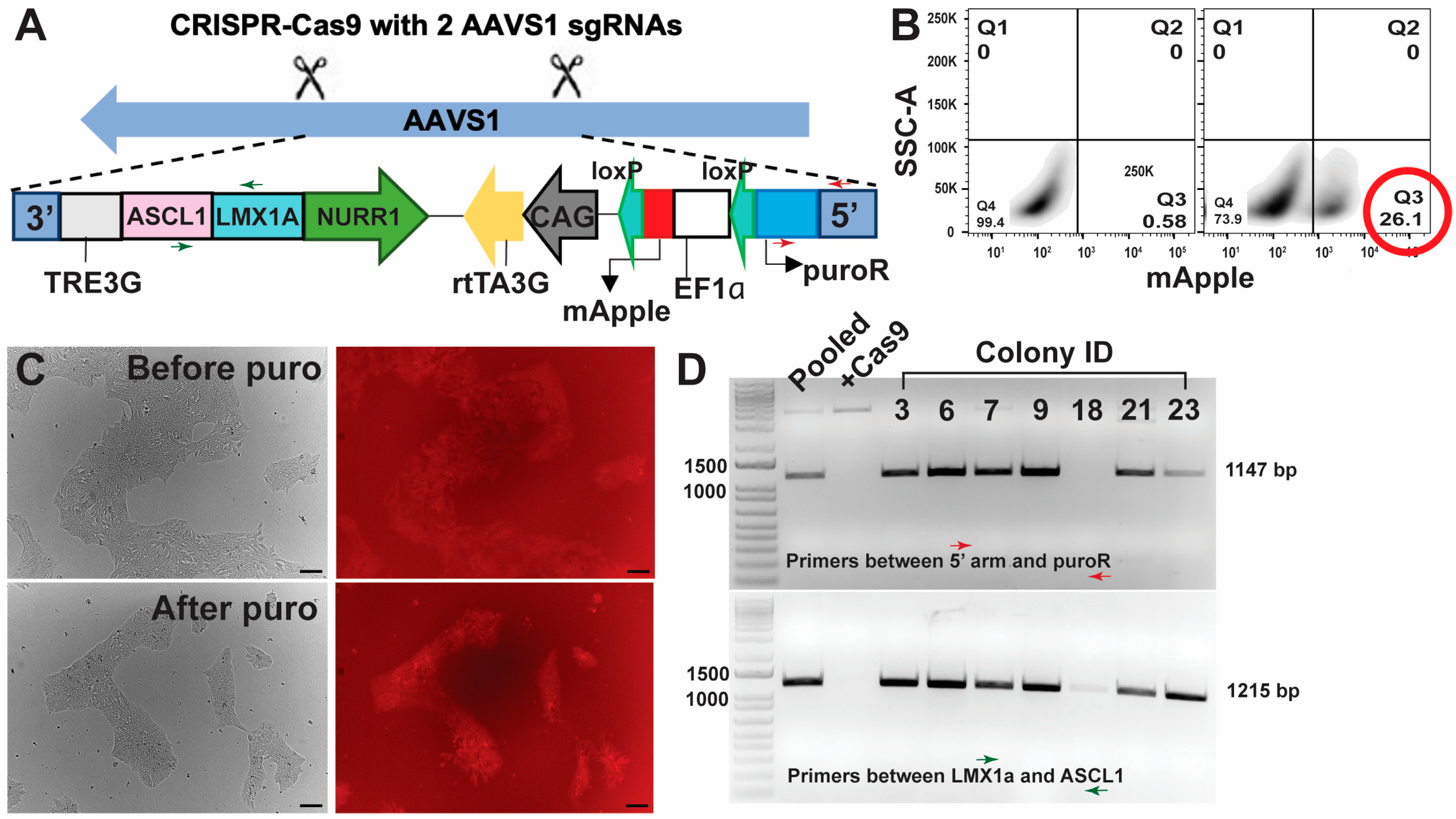
2.2.2. Integration of Transcription Factors Using the CRISPR-Cas9 Approach
2.2.3. Genotyping of iPSCs with Integrated hALAN
2.2.4. Differentiation of iPSCs into iDAs
2.3. iPSC-Derived Dopaminergic Neuron Generation following Nishimura et al.
2.4. iPSC-Derived Dopaminergic Neuron Generation following Sheta et al.
2.5. Immunocytochemistry and Calcium Signalling on iPSC-Derived iDAs
2.6. Dopamine Release Assay on iPSC-Derived iDAs
2.7. Image Analysis and Statistical Analysis
3. Results
3.1. Targeted and Stable Integration in iPSCs Maintained Transcription Factor Expression Better than Lentiviral Transduction
3.2. Differentiation following Theka et al. and Nishimura et al.
3.3. Differentiation following Sheta et al.
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schondorf, D.C.; Aureli, M.; McAllister, F.E.; Hindley, C.J.; Mayer, F.; Schmid, B.; Sardi, S.P.; Valsecchi, M.; Hoffmann, S.; Schwarz, L.K.; et al. iPSC-derived neurons from GBA1-associated Parkinson’s disease patients show autophagic defects and impaired calcium homeostasis. Nat. Commun. 2014, 5, 4028. [Google Scholar] [CrossRef] [PubMed]
- Tcw, J. Human iPSC application in Alzheimer’s disease and Tau-related neurodegenerative diseases. Neurosci. Lett. 2019, 699, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Moretti, A.; Laugwitz, K.L.; Dorn, T.; Sinnecker, D.; Mummery, C. Pluripotent stem cell models of human heart disease. Cold Spring Harb. Perspect. Med. 2013, 3, a014027. [Google Scholar] [CrossRef] [PubMed]
- Imberti, B.; Tomasoni, S.; Ciampi, O.; Pezzotta, A.; Derosas, M.; Xinaris, C.; Rizzo, P.; Papadimou, E.; Novelli, R.; Benigni, A.; et al. Renal progenitors derived from human iPSCs engraft and restore function in a mouse model of acute kidney injury. Sci. Rep. 2015, 5, 8826. [Google Scholar] [CrossRef] [PubMed]
- Theka, I.; Caiazzo, M.; Dvoretskova, E.; Leo, D.; Ungaro, F.; Curreli, S.; Manago, F.; Dell’Anno, M.T.; Pezzoli, G.; Gainetdinov, R.R.; et al. Rapid generation of functional dopaminergic neurons from human induced pluripotent stem cells through a single-step procedure using cell lineage transcription factors. Stem Cells Transl. Med. 2013, 2, 473–479. [Google Scholar] [CrossRef]
- Kriks, S.; Shim, J.W.; Piao, J.; Ganat, Y.M.; Wakeman, D.R.; Xie, Z.; Carrillo-Reid, L.; Auyeung, G.; Antonacci, C.; Buch, A.; et al. Dopamine neurons derived from human ES cells efficiently engraft in animal models of Parkinson’s disease. Nature 2011, 480, 547–551. [Google Scholar] [PubMed]
- Sagal, J.; Zhan, X.; Xu, J.; Tilghman, J.; Karuppagounder, S.S.; Chen, L.; Dawson, V.L.; Dawson, T.M.; Laterra, J.; Ying, M. Proneural transcription factor Atoh1 drives highly efficient differentiation of human pluripotent stem cells into dopaminergic neurons. Stem Cells Transl. Med. 2014, 3, 888–898. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, K.; Nitta, T.; Doi, K.; Takata, K. Rapid Conversion of Human Induced Pluripotent Stem Cells into Dopaminergic Neurons by Inducible Expression of Two Transcription Factors. Stem Cells Dev. 2022, 31, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Sheta, R.; Teixeira, M.; Idi, W.; Pierre, M.; de Rus Jacquet, A.; Emond, V.; Zorca, C.E.; Vanderperre, B.; Durcan, T.M.; Fon, E.A.; et al. Combining NGN2 programming and dopaminergic patterning for a rapid and efficient generation of hiPSC-derived midbrain neurons. Sci. Rep. 2022, 12, 17176. [Google Scholar] [CrossRef]
- Cardo, L.F.; Monzon-Sandoval, J.; Li, Z.; Webber, C.; Li, M. Single-Cell Transcriptomics and In Vitro Lineage Tracing Reveals Differential Susceptibility of Human iPSC-Derived Midbrain Dopaminergic Neurons in a Cellular Model of Parkinson’s Disease. Cells 2023, 12, 2860. [Google Scholar] [CrossRef]
- Mahajani, S.; Raina, A.; Fokken, C.; Kugler, S.; Bahr, M. Homogenous generation of dopaminergic neurons from multiple hiPSC lines by transient expression of transcription factors. Cell Death Dis. 2019, 10, 898. [Google Scholar] [CrossRef]
- Chen, Y.; Kuang, J.; Niu, Y.; Zhu, H.; Chen, X.; So, K.F.; Xu, A.; Shi, L. Multiple factors to assist human-derived induced pluripotent stem cells to efficiently differentiate into midbrain dopaminergic neurons. Neural Regen. Res. 2024, 19, 908–914. [Google Scholar] [CrossRef]
- Zheng, Y.Y.; Xu, H.; Wang, Y.S. Progress in direct reprogramming of dopaminergic cell replacement therapy. Neurol. Sci. 2023, 45, 873–881. [Google Scholar] [CrossRef]
- Caiazzo, M.; Dell’Anno, M.T.; Dvoretskova, E.; Lazarevic, D.; Taverna, S.; Leo, D.; Sotnikova, T.D.; Menegon, A.; Roncaglia, P.; Colciago, G.; et al. Direct generation of functional dopaminergic neurons from mouse and human fibroblasts. Nature 2011, 476, 224–227. [Google Scholar] [CrossRef]
- Nakamura, R.; Nonaka, R.; Oyama, G.; Jo, T.; Kamo, H.; Nuermaimaiti, M.; Akamatsu, W.; Ishikawa, K.I.; Hattori, N. A defined method for differentiating human iPSCs into midbrain dopaminergic progenitors that safely restore motor deficits in Parkinson’s disease. Front. Neurosci. 2023, 17, 1202027. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Xia, N.; Reijo Pera, R.A. Directed dopaminergic neuron differentiation from human pluripotent stem cells. J. Vis. Exp. 2014, 91, 51737. [Google Scholar] [CrossRef]
- Stathakos, P.; Jimenez-Moreno, N.; Crompton, L.A.; Nistor, P.A.; Badger, J.L.; Barbuti, P.A.; Kerrigan, T.L.; Randall, A.D.; Caldwell, M.A.; Lane, J.D. A monolayer hiPSC culture system for autophagy/mitophagy studies in human dopaminergic neurons. Autophagy 2021, 17, 855–871. [Google Scholar] [CrossRef]
- Raposo, A.; Vasconcelos, F.F.; Drechsel, D.; Marie, C.; Johnston, C.; Dolle, D.; Bithell, A.; Gillotin, S.; van den Berg, D.L.C.; Ettwiller, L.; et al. Ascl1 Coordinately Regulates Gene Expression and the Chromatin Landscape during Neurogenesis. Cell Rep. 2015, 10, 1544–1556. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, F.M.; van der Linden, A.J.; Wang, Y.; von Oerthel, L.; Sul, H.S.; Burbach, J.P.; Smidt, M.P. Identification of Dlk1, Ptpru and Klhl1 as novel Nurr1 target genes in meso-diencephalic dopamine neurons. Development 2009, 136, 2363–2373. [Google Scholar] [CrossRef]
- Andersson, E.; Tryggvason, U.; Deng, Q.; Friling, S.; Alekseenko, Z.; Robert, B.; Perlmann, T.; Ericson, J. Identification of intrinsic determinants of midbrain dopamine neurons. Cell 2006, 124, 393–405. [Google Scholar] [CrossRef]
- Zhang, Y.; Pak, C.; Han, Y.; Ahlenius, H.; Zhang, Z.; Chanda, S.; Marro, S.; Patzke, C.; Acuna, C.; Covy, J.; et al. Rapid single-step induction of functional neurons from human pluripotent stem cells. Neuron 2013, 78, 785–798. [Google Scholar] [CrossRef] [PubMed]
- Fernandopulle, M.S.; Prestil, R.; Grunseich, C.; Wang, C.; Gan, L.; Ward, M.E. Transcription Factor-Mediated Differentiation of Human iPSCs into Neurons. Curr. Protoc. Cell Biol. 2018, 79, e51. [Google Scholar] [CrossRef] [PubMed]
- Ihry, R.J.; Worringer, K.A.; Salick, M.R.; Frias, E.; Ho, D.; Theriault, K.; Kommineni, S.; Chen, J.; Sondey, M.; Ye, C.; et al. p53 inhibits CRISPR-Cas9 engineering in human pluripotent stem cells. Nat. Med. 2018, 24, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.M.; Hope, K.M.; Xu, J.B.; Mitchell, N.L.; Palmer, D.N. Inhibition of storage pathology in prenatal CLN5-deficient sheep neural cultures by lentiviral gene therapy. Neurobiol. Dis. 2014, 62, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Basak, I.; Bhatlekar, S.; Manne, B.K.; Stoller, M.; Hugo, S.; Kong, X.; Ma, L.; Rondina, M.T.; Weyrich, A.S.; Edelstein, L.C.; et al. miR-15a-5p regulates expression of multiple proteins in the megakaryocyte GPVI signaling pathway. J. Thromb. Haemost. 2019, 17, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Pedersen, O.Z.; Peng, J.; Couture, L.A.; Rao, M.S.; Zeng, X. Optimizing dopaminergic differentiation of pluripotent stem cells for the manufacture of dopaminergic neurons for transplantation. Cytotherapy 2013, 15, 999–1010. [Google Scholar] [CrossRef]
- Basak, I.; Hansen, R.A.; Ward, M.E.; Hughes, S.M. Deficiency of the Lysosomal Protein CLN5 Alters Lysosomal Function and Movement. Biomolecules 2021, 11, 1412. [Google Scholar] [CrossRef] [PubMed]
- Muto, A.; Ohkura, M.; Abe, G.; Nakai, J.; Kawakami, K. Real-time visualization of neuronal activity during perception. Curr. Biol. 2013, 23, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Ren, Y.; Yuen, E.Y.; Zhong, P.; Ghaedi, M.; Hu, Z.; Azabdaftari, G.; Nakaso, K.; Yan, Z.; Feng, J. Parkin controls dopamine utilization in human midbrain dopaminergic neurons derived from induced pluripotent stem cells. Nat. Commun. 2012, 3, 668. [Google Scholar] [CrossRef]
- Reyes, S.; Fu, Y.; Double, K.; Thompson, L.; Kirik, D.; Paxinos, G.; Halliday, G.M. GIRK2 expression in dopamine neurons of the substantia nigra and ventral tegmental area. J. Comp. Neurol. 2012, 520, 2591–2607. [Google Scholar] [CrossRef]
- Tian, R.; Gachechiladze, M.A.; Ludwig, C.H.; Laurie, M.T.; Hong, J.Y.; Nathaniel, D.; Prabhu, A.V.; Fernandopulle, M.S.; Patel, R.; Abshari, M.; et al. CRISPR Interference-Based Platform for Multimodal Genetic Screens in Human iPSC-Derived Neurons. Neuron 2019, 104, 239–255.e12. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.C.; Ulane, C.M.; Burke, R.E. Clinical progression in Parkinson disease and the neurobiology of axons. Ann. Neurol. 2010, 67, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Sheta, R.; Teixeira, M.; Idi, W.; Oueslati, A. Optimized protocol for the generation of functional human induced-pluripotent-stem-cell-derived dopaminergic neurons. STAR Protoc. 2023, 4, 102486. [Google Scholar] [CrossRef] [PubMed]
- Powell, S.K.; O’Shea, C.; Townsley, K.; Prytkova, I.; Dobrindt, K.; Elahi, R.; Iskhakova, M.; Lambert, T.; Valada, A.; Liao, W.; et al. Induction of dopaminergic neurons for neuronal subtype-specific modeling of psychiatric disease risk. Mol. Psychiatry 2023, 28, 1970–1982. [Google Scholar] [CrossRef] [PubMed]
- Alekseenko, Z.; Dias, J.M.; Adler, A.F.; Kozhevnikova, M.; van Lunteren, J.A.; Nolbrant, S.; Jeggari, A.; Vasylovska, S.; Yoshitake, T.; Kehr, J.; et al. Robust derivation of transplantable dopamine neurons from human pluripotent stem cells by timed retinoic acid delivery. Nat. Commun. 2022, 13, 3046. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Jiang, H.; Li, H.; Li, L.; Yan, Z.; Feng, J. Generation of human A9 dopaminergic pacemakers from induced pluripotent stem cells. Mol. Psychiatry 2022, 27, 4407–4418. [Google Scholar] [CrossRef]
- Otero, M.G.; Bell, S.; Laperle, A.H.; Lawless, G.; Myers, Z.; Castro, M.A.; Villalba, J.M.; Svendsen, C.N. Organ-Chips Enhance the Maturation of Human iPSC-Derived Dopamine Neurons. Int. J. Mol. Sci. 2023, 24, 14227. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McDonald, K.O.; Lyons, N.M.A.; Gray, L.K.C.; Xu, J.B.; Schoderboeck, L.; Hughes, S.M.; Basak, I. Transcription Factor-Mediated Generation of Dopaminergic Neurons from Human iPSCs—A Comparison of Methods. Cells 2024, 13, 1016. https://doi.org/10.3390/cells13121016
McDonald KO, Lyons NMA, Gray LKC, Xu JB, Schoderboeck L, Hughes SM, Basak I. Transcription Factor-Mediated Generation of Dopaminergic Neurons from Human iPSCs—A Comparison of Methods. Cells. 2024; 13(12):1016. https://doi.org/10.3390/cells13121016
Chicago/Turabian StyleMcDonald, Kirstin O., Nikita M. A. Lyons, Luca K. C. Gray, Janet B. Xu, Lucia Schoderboeck, Stephanie M. Hughes, and Indranil Basak. 2024. "Transcription Factor-Mediated Generation of Dopaminergic Neurons from Human iPSCs—A Comparison of Methods" Cells 13, no. 12: 1016. https://doi.org/10.3390/cells13121016
APA StyleMcDonald, K. O., Lyons, N. M. A., Gray, L. K. C., Xu, J. B., Schoderboeck, L., Hughes, S. M., & Basak, I. (2024). Transcription Factor-Mediated Generation of Dopaminergic Neurons from Human iPSCs—A Comparison of Methods. Cells, 13(12), 1016. https://doi.org/10.3390/cells13121016