Abstract
The reciprocal modulation between the CXCL12/CXCR4/ACKR3 axis and the STAT3 signaling pathway plays a crucial role in the progression of various diseases and neoplasms. Activation of the CXCL12/CXCR4/ACKR3 axis triggers the STAT3 pathway through multiple mechanisms, while the STAT3 pathway also regulates the expression of CXCL12. This review offers a thorough and systematic analysis of the reciprocal regulatory mechanisms between the CXCL12/CXCR4/ACKR3 signaling axis and the STAT3 signaling pathway in the context of diseases, particularly tumors. It explores the potential clinical applications in tumor treatment, highlighting possible therapeutic targets and novel strategies for targeted tumor therapy.
1. Introduction
Crosstalk between the CXCL12/CXCR4/ACKR3 axis and the STAT3 signaling pathway has been hypothesized on the basis of findings that both the CXCL12/CXCR4/ACKR3 axis and STAT3 signaling pathway are involved in the progression of several cancers. In addition, it has been suggested that crosstalk between the CXCL12/CXCR4/ACKR3 axis and the STAT3 signaling pathway plays a role in other systems.
In the following sections, we will introduce the CXCL12/CXCR4/ACKR3 Axis and the STAT3 signaling pathway, followed by a summary of suggested mechanisms leading to observed CXCL12/CXCR4/ACKR3 Axis and STAT3 pathway crosstalk. We will discuss targeting opportunities for simultaneous regulation of the CXCL12/CXCR4/ACKR3 Axis and STAT3 pathway in therapeutic development.
2. Background
2.1. The CXCL12/CXCR4/ACKR3 Axis
CXCL12, also referred to as stromal cell-derived factor-1 (SDF-1) or pre-B cell growth-stimulating factor (PBSF), is a member of the CXC chemokine subfamily. Current research indicates that CXCL12 possesses two specific receptors, namely, CXCR4 and ACKR3. CXCL12 is the only recognized CXC chemokine that undergoes differential mRNA splicing [1]. As a result of its selective gene splicing, seven isoforms of CXCL12 have been identified: α, β, γ, δ, ε, θ, and iso7. Among these isoforms, α and β are the most prevalent and have been extensively studied [2]. The seven isoforms are encoded by the same CXCL12 gene and share the first three exons. Their distinction lies in the fourth exon, which determines the length of the splice variants. In fact, all CXCL12 isoforms share the initial 67 amino acids, albeit varying in length [3]. CXCL12α, composed of 68 amino acids, is expressed in various tissues, with a significant enrichment in stromal cells and endothelial cells. It participates in a wide range of physiological and pathological processes. In contrast, CXCL12β, which contains an additional four amino acids compared to CXCL12α, exhibits similar functional properties. However, the β isoform possesses enhanced angiogenic potential and is predominantly enriched in vascular-rich organs such as the kidney, liver, and spleen [4,5]. The remaining splice variants of CXCL12 are distinguished by differences in their fourth exon [3]. CXCL12γ is primarily found in metabolically active organs with low vascularization, such as the brain and heart. Notably, the γ isoform exhibits the highest affinity for its receptor, CXCR4, and exerts the longest-lasting downstream effects [6].
The C-X-C chemokine receptor 4 (CXCR4), also known as CD184, is a member of the seven-transmembrane G protein-coupled receptor family. CXCR4 is a chemokine receptor akin to rhodopsin, characterized by seven transmembrane domains and typically classified as a G protein-coupled receptor (GPCR) [7]. CXCL12 binds to the extracellular domains of CXCR4, leading to the conversion of GDP-bound G proteins to their GTP-bound forms. This conversion results in the dissociation of heterotrimeric G proteins into Gα subunits and Gβγ subunits. The released Gα and Gβγ subunits subsequently activate downstream molecules, including protein kinase B (AKT), c-Jun N-terminal kinase (JNK), mitogen-activated protein kinase (MEK), and extracellular signal-regulated kinase 1/2 (ERK1/2), thereby modulating transcription [8]. In addition, the CXCL12/CXCR4 signaling axis can activate the Janus kinases/signal transducers and activators of transcription (JAK/STAT) pathways [9], the Wnt/β-catenin signaling pathway [10], and the Ca2+/calmodulin-dependent protein kinase II/AMP response element-binding protein (CaMKII/CREB) signaling pathway [11]. However, the specific G protein subunit through which CXCR4 activates these pathways remains an area of active investigation. Based on the coupling Gα subunits, diverse GPCR signaling pathways can be classified into four families: Gαs, Gαi, Gαq, and Gα12 [12]. Initially, CXCR4 was categorized as a Gαi protein-coupled receptor [13]. Upon the binding of CXCL12 to CXCR4, the G protein-coupled receptor (GPCR) is activated. The activation of the Gαi subunit inhibits adenylyl cyclase, which catalyzes the conversion of 5′-triphosphate adenosine (ATP) to cyclic adenosine monophosphate (cAMP). The cAMP molecules bind to the two cAMP-binding domains in the regulatory subunit of protein kinase A (PKA), leading to the release of PKA’s two catalytic subunits. Consequently, downstream target proteins are activated and phosphorylated [14,15].
ACKR3, also known as CXCR7 or RDC1, is a member of the seven-transmembrane G protein-coupled receptor (GPCR) family, akin to CXCR4. Unlike CXCR4, ACKR3 is an atypical G protein-coupled receptor. Despite ACKR3 harboring two amino acid substitutions within the prototypical DRYLAIV motif for G protein interaction, the reconstitution of this motif does not enable ACKR3 to signal via G proteins. Consequently, ACKR3 demonstrates a reduced affinity for G proteins, while it exhibits an elevated affinity for the scaffold protein β-arrestin2 (ARRB2) [16,17]. As a result, upon binding to its ligand CXCL12, ACKR3 does not dissociate from G proteins. Instead, it activates the G protein-coupled receptor kinase 2 (GRK2) and undergoes receptor phosphorylation, leading to conformational changes. Subsequently, ARRB2 is recruited to activate downstream signaling pathways. Notably, studies have shown that in a mouse model with CXCR4 knockout, ACKR3-positive neural progenitor cells exhibit migration along the CXCL12 gradient in vitro [18]. Moreover, the inhibition of ACKR3 suppresses CXCL12-induced migration of cervical cancer cells, whereas CXCR4 does not participate in these processes [19]. Therefore, the signaling pathways mediated by ACKR3 may diverge from the canonical chemokine pathways governed by CXCR4. By binding to CXCR4/ACKR3 through autocrine or paracrine mechanisms, CXCL12 activates downstream signaling pathways, including STAT3, NFκB, and MAPK. These pathways are instrumental in various biological processes, such as development, homeostasis maintenance, the regulation of inflammatory responses, and the facilitation of malignant tumor progression [20,21,22,23].
2.2. The STAT3 Signaling Pathway
The Signal Transducer and Activator of Transcription (STAT) protein family consists of seven members: STAT1, STAT2, STAT3, STAT4, STAT5A, STAT5B, and STAT6 [24]. These proteins can transmit signals from both receptor-associated and non-receptor-associated kinases to the cell nucleus [25]. Among the seven members of the STAT protein family, STAT3 emerges as the most potent promoter of tumor growth and immune evasion [26,27]. Notably, it is the only family member whose genetic deletion results in embryonic lethality [28,29].
STAT3 is a protein composed of 770 amino acids and is characterized by six functionally conserved domains. In its inactive state, STAT3 exists as a monomer within the cytoplasm. Studies have indicated that the presence of STAT3 in the cytoplasm is crucial for maintaining the pluripotency of embryonic stem cells and regulating immune responses [29,30]. The Src homology 2 (SH2) domain is the most conserved structural motif within STAT proteins. Upon tyrosine phosphorylation by Janus kinase (JAK)-mediated growth factor receptors, such as gp130, the SH2 domain of STAT3 recognizes and binds to specific phosphotyrosine motifs, playing a critical role in signal transduction [31]. For instance, phosphorylated STAT3 induces the upregulation of regulatory factors involved in cell proliferation (e.g., Cyclin D1 and MYC) and survival (e.g., BCL-xL and survivin), as well as the expression of proangiogenic factors (e.g., VEGF) and immune-suppressive cytokines (e.g., IL-6) [26,32].
The canonical STAT3 pathway is triggered when cytokines such as interleukin-6 (IL-6) bind to their respective receptors (e.g., gp130/IL-6Rα). This ligand–receptor interaction induces the phosphorylation of receptor-associated kinases, known as Janus kinases (JAKs). The subsequent phosphorylation of JAKs leads to the tyrosine phosphorylation of STAT3 at residue Tyr705, thereby activating the STAT3 pathway. Once activated, STAT3 forms homodimers and dissociates from the receptor, translocating into the cell nucleus. Within the nucleus, it binds to specific promoter DNA sequences, driving gene transcription. The activation of the STAT3 pathway, whether through intrinsic or extrinsic mechanisms, exerts a profound influence on the tumor microenvironment. It promotes tumor proliferation, angiogenesis, and immune evasion and contributes to tumor metastasis through various mechanisms, including epithelial–mesenchymal transition and the upregulation of matrix metalloproteinases [33,34,35,36,37]. Intrinsic mechanisms include JAK2/STAT3 signaling transduction and intracellular STAT3’s direct transcriptional regulation of genes associated with tumor progression, such as those promoting the differentiation and cytotoxic functions of tumor-specific T cells [38]. Extrinsic mechanisms encompass the activation of the STAT3 pathway with ionizing radiation [39] or with cytokines produced by inflammatory cells within the tumor microenvironment [40], thereby facilitating tumor development.
The activation of the STAT3 pathway leads to the upregulation of VEGF and specific matrix metalloproteinases (MMPs). These factors play distinct roles: VEGF fosters angiogenesis, while MMPs contribute to invasive processes that propel tumor metastasis [41]. Additionally, STAT3 forms a positive feedback loop by binding to the IL-6 promoter, resulting in an increase in IL-6 expression [42]. Both VEGF and IL-6 possess immunosuppressive properties, facilitating immune evasion by tumor cells with excessive STAT3 activation [43,44]. Furthermore, the protumorigenic effects of STAT3 may involve the induction of miR-21 and miR-181b-1, which, respectively, inhibit the expression of tumor suppressors PTEN and ubiquitin carboxyl-terminal hydrolase CYLD [45]. Notably, apoptosis-related proteins B-cell lymphoma-2 (Bcl-2) and Bcl-2-associated protein X (Bax) serve as transcriptional targets of the STAT3 pathway [46,47]. Upon STAT3 activation, the inhibition of Bcl-2 and Bax promotes cell survival.
The STAT3 pathway can be activated by several kinases, including the epidermal growth factor receptor (EGFR), vascular endothelial growth factor receptor 2 (VEGFR2), hepatocyte growth factor receptor (HGFR), platelet-derived growth factor receptor (PDGFR), SRC, and ABL. Among these, EGFR, VEGFR2, HGFR, and PDGFR are classified as receptor tyrosine kinases (RTKs) with intrinsic tyrosine kinase activities. These RTKs phosphorylate JAK, thereby facilitating the activation of the STAT3 pathway. Additionally, non-receptor tyrosine kinases (nRTKs) such as Src or ABL can also activate STAT3. At the molecular level, Src or ABL induces tyrosine phosphorylation, leading to STAT3 activation and subsequent translocation into the cell nucleus, where it exerts its regulatory functions on gene transcription [48]. Furthermore, in various tumors, including lung cancer, breast cancer, and colorectal cancer, the activation of the CXCL12/CXCR4/ACKR3 signaling axis has been associated with STAT3 pathway activation [49,50,51,52].
The STAT3 pathway is subject to negative regulation by various cellular factors, including suppressor of cytokine signaling (SOCS), protein inhibitor of activated STAT (PIAS), and protein tyrosine phosphatases (PTPs). Among these, SOCS is a protein family characterized by a central Src homology 2 (SH2) domain. SOCS3 is a member of the SOCS family and acts as a negative feedback loop for the STAT3 signaling pathway. Specifically, SOCS3 interacts with the JAK2 domain (or the intracellular portion of the receptor) to exert its effects [53]. In particular, the kinase inhibitory region of SOCS3 blocks the substrate-binding groove on JAK2, thereby inhibiting its kinase activity and promoting the ubiquitination and degradation of receptor proteins, which in turn suppresses STAT3 phosphorylation [53,54]. Additionally, SOCS can interact with receptor tyrosine kinases (RTKs), thereby disrupting STAT3 recruitment [55,56]. Notably, SOCS does not inhibit the non-receptor tyrosine kinase SRC-mediated activation of the STAT3 signaling pathway [57]. The deletion of SOCS3 leads to STAT3 pathway activation, promoting cell proliferation and increasing the incidence of hepatitis-induced liver cancer [58]. Importantly, the upregulation of SOCS3 inhibits tumor growth and metastasis. Exogenous SOCS3 suppresses the growth of human non-small cell lung cancer cells and enhances their sensitivity to radiotherapy [59].
The protein inhibitor of activated STAT (PIAS) functions as an inhibitor of STAT transcriptional activation. PIAS proteins can inhibit STAT transcription through various mechanisms, including blocking the DNA-binding activity of transcription factors, recruiting transcriptional repressors, and facilitating protein SUMOylation modifications (analogous to ubiquitination) [60]. Among these, PIAS3 specifically interacts with activated STAT3 dimers, preventing their binding to DNA and thereby inhibiting STAT3 transcriptional activity [61]. In glioblastoma tissues, an elevated expression of PIAS3 promotes cell proliferation [62,63]. Furthermore, the upregulation of PIAS3 suppresses cell proliferation and enhances chemosensitivity in various tumors. For instance, curcumin, by upregulating PIAS3 in ovarian and endometrial cancer cells, inhibits STAT3 activation and restrains tumor cell growth [64]. Additionally, the overexpression of PIAS3 inhibits lung cancer cell growth and restores sensitivity to chemotherapy [65,66].
Protein tyrosine phosphatases (PTPs) play a pivotal role in negatively regulating STAT3 activity. Seven types of PTPs have been identified as targeting STAT3: PTPRD, PTPRT, PTPRK, SHP-1, SHP-2, PTPN9, and TC-PTP. PTPRD, PTPRT, and PTPRK directly interact with STAT3, dephosphorylating STAT3 at the Tyr705 site to inhibit STAT3 signal transduction [67,68,69]. SHP-1 and SHP-2 possess a phosphatase domain and two SH2 domains. Through their SH2 domains, they interact with target proteins (such as JAK/STAT3, Src family kinases, and growth factor receptors), dephosphorylating substrates via their phosphatase activity [70,71]. In colorectal cancer, the p53R248Q mutation competitively binds to SHP2, leading to STAT3 activation [72]. In 2012, PTPN9 was first reported to induce STAT3 dephosphorylation, suppressing breast cancer growth [73]. Additionally, STAT3 is one of the substrates for TC-PTP. Deficiency of TC-PTP in triple-negative primary breast cancer cells enhances cell proliferation through STAT3 signaling activation and increased Src family kinase activity [74]. Overexpression of TC-PTP significantly inhibits in vitro cell proliferation and suppresses xenograft growth in human breast cancer cell lines [74].
The interaction between the CXCL12/CXCR4/ACKR3 axis and the STAT3 signaling pathway is crucial in the development and progression of various diseases, including cancer. In breast cancer, the CXCL12/CXCR4/ACKR3 axis promoted breast cancer metastasis through the activation of the STAT3 pathway [52,75]. In skin-related disorders, CXCL12 activated the STAT3 pathway, inhibiting hair growth [76]. Additionally, the STAT3 pathway regulated the expression of CXCL12. Understanding the relationship between the CXCL12/CXCR4/ACKR3 axis and the STAT3 pathway could lead to new therapeutic targets for innovative cancer treatments.
3. Crosstalk between CXCL12/CXCR4/ACKR3 Signaling Axis and STAT3 Pathway
The CXCL12/CXCR4/ACKR3 signaling axis and the STAT3 pathway are intimately linked with immune regulation and the malignant progression of a variety of diseases, encompassing skin disorders, brain injuries, myocardial damage, and tumors [24,77]. Within this context, the CXCL12/CXCR4/ACKR3 axis exerts both positive and negative effects through the activation of STAT3. Moreover, the activated STAT3 pathway can augment CXCL12 expression, playing a pivotal role in the progression of these diseases. Understanding the relationship between the CXCL12/CXCR4/ACKR3 axis and the STAT3 pathway could lead to new therapeutic targets for innovative cancer treatments
3.1. CXCL12/CXCR4/ACKR3 and STAT3 Pathway Crosstalk in Cancer
Research findings indicate that the CXCL12 and STAT3 proteins are significantly upregulated in various malignancies, including breast cancer, lung cancer, hepatocellular carcinoma, esophageal cancer, bladder cancer, and leukemia [78,79,80]. Notably, tumor cells positive for CXCR4 migrate along the CXCL12 concentration gradient to distant organs exhibiting high CXCL12 expression, thereby facilitating tumor metastasis [81]. In bladder cancer tissues, a positive correlation exists between elevated CXCR4 expression and positive p-STAT3 expression [82]. These findings suggest an activating interplay between the CXCL12/CXCR4 signaling axis and the STAT3 pathway.
The CXCL12/CXCR4/ACKR3 signaling axis activates the STAT3 pathway, playing a central role in tumor proliferation, metastasis, anti-apoptosis, maintenance of tumor stemness, and immune escape. In breast cancer tissues, dual positivity for CXCR4 and phosphorylated STAT3 (p-STAT3) correlates positively with tumor size, lymph node metastasis, and histological grade [52]. Further investigations reveal that the CXCL12/CXCR4 axis phosphorylates JAK2, thereby activating the STAT3 signaling pathway to promote breast cancer proliferation and metastasis. Inhibition of either CXCR4 or JAK2 suppresses STAT3 phosphorylation, reversing the proliferative and metastatic phenotypes of breast cancer [52]. Using a mouse orthotopic tumor model constructed with 4T1.2 breast cancer cells, it was observed that 4T1.2 cells can metastasize to the lungs. Cell experiments demonstrated that CXCL12 upregulates STAT3 phosphorylation, while knocking down ACKR3 attenuates the upregulation of p-STAT3Y705. These findings suggest that the CXCL12/ACKR3 axis activates the STAT3 signaling pathway. Immunohistochemical analysis of the in situ tumors revealed concurrent elevation of p-STAT3 and the vascular marker CD31, as well as the proliferation marker Ki67, promoting tumor metastasis to the lungs [75].
Epithelial–mesenchymal transition (EMT) plays a pivotal role in tumor metastasis [83]. Furthermore, the enhancement of a tumor’s mesenchymal phenotype is associated with chemotherapy resistance and unfavorable outcomes [84]. In esophageal cancer, in vitro experiments have demonstrated that the CXCL12/ACKR3 axis activates the STAT3 pathway. This activation leads to the downregulation of the epithelial cell marker protein E-Cadherin and the upregulation of mesenchymal cell markers N-Cadherin, Vimentin, Snail, and Slug. Consequently, this process promotes EMT and accelerates the metastasis of esophageal cancer. In vivo tumor transplantation experiments have also revealed that the upregulation of ACKR3 enhances the expression of Ki67 in tumor tissues, thereby promoting the growth and metastasis of esophageal cancer [49]. Similarly, in gastric cancer, activation of the CXCL12/CXCR4 axis via the STAT3 pathway induces downregulation of E-Cadherin and upregulation of N-Cadherin, Vimentin, and Snail, enhancing EMT, migration, and the invasiveness of gastric cancer cells [85]. Additionally, Yang et al. found that downregulation of ACKR3 in acute T-cell lymphoblastic leukemia inhibits STAT3 pathway activation, suppressing tumor growth and lung metastasis in vivo [86]. The silencing of ACKR3 also leads to decreased matrix metalloproteinase-2 (MMP2) and vascular endothelial growth factor (VEGF) expression, inhibiting migration and invasion of tumor endothelial cells in hepatocellular carcinoma [87]. Furthermore, in gastric cancer, ACKR3 has been implicated in promoting tumor growth and metastasis via the STAT3/c-Myc pathway [88].
Tumor stem cells (CSCs) constitute a critical cellular subset in the heterogeneous development of tumors, playing pivotal roles in tumor metastasis, recurrence, and drug resistance [89]. Hong et al. demonstrated that activation of the CXCL12/CXCR4 axis induces immortalized embryonic kidney cells to transform into cells resembling tumor stem cells, potentially contributing to carcinogenesis [90]. In hepatocellular carcinoma, the activation of the CXCL12/CXCR4 axis via the STAT3 signaling pathway promotes protein expression of CSC markers SOX2 and CD133, enhancing the spheroid-forming capacity and invasiveness of liver cancer cells [91].
Additionally, resistance to apoptosis in tumors promotes proliferation and metastasis. Ahn et al. proposed that 3-formylchromone downregulates p-STAT3, attenuating the CXCL12-driven migration and invasiveness of liver cancer cells. Simultaneously, 3-formylchromone inhibits the expression of Bcl-2, Bcl-xL, Survivin, and Mcl-1 in liver cancer cells, suggesting that 3-formylchromone promotes apoptosis in liver cancer cells by suppressing the STAT3 pathway and weakening CXCL12-driven metastasis [70]. In non-small cell lung cancer, in vitro experiments demonstrate that CXCL12 binding to CXCR4, via activation of the JAK2/STAT3 signaling pathway, reduces the proportion of lung cancer cell apoptosis induced by the chemotherapeutic drug cisplatin. Clinical lung cancer tissue samples also reveal a positive correlation between elevated CXCL12 and positive p-STAT3 expression and the malignant progression of lung cancer [50].
Autophagy is a fundamental cellular process that maintains homeostasis by degrading cellular components through lysosomal machinery. It serves as a critical mechanism for supplying cells with essential biomolecules and energy during periods of stress. Selective autophagy is mediated by autophagic receptors, with p62 being one of the earliest identified autophagic receptor proteins. Notably, p62 is closely associated with tumor development [92]. In the context of bladder cancer, cancer-associated fibroblasts (CAFs) secrete CXCL12, which activates the JAK2/STAT3 signaling pathway, leading to the accumulation of p62. This, in turn, promotes the migration and invasion of bladder cancer cells and upregulates the expression of programmed cell death ligand 1 (PDL-1) via non-degradative pathways [93]. Further investigations reveal that CXCL12 secreted by CAFs, upon binding to its specific receptor CXCR4, activates JAK2/STAT3 signaling, resulting in increased expression of cylindromatosis (CYLD), a deubiquitinase. CYLD deubiquitinates p62, leading to its accumulation and the subsequent inhibition of PDL-1 degradation via autophagy pathways [93]. Elevated PD-L1 expression can facilitate immune escape by tumor cells [94,95].
Tumor immune escape, which encompasses both immune suppression and immune resistance, represents a key malignant phenotype of tumor cells. CXCL12, secreted by cancer-associated fibroblasts (CAFs), activates the STAT3 pathway, inducing the generation and activation of myeloid-derived suppressor cells, resulting in tumor immune escape and further promoting tumor development [96]. Multiple studies have demonstrated that within leukemia, the CXCL12/CXCR4 axis plays a pivotal role in maintaining the interactions between leukemia cells and the bone marrow niche. This axis facilitates the adhesion of leukemia cells to the bone marrow, enhancing cell survival and reducing sensitivity to signal transduction inhibitors and chemotherapeutic agents [97,98,99,100]. Additionally, Madrazo E et al. discovered that CXCL12 promotes H3K9 methylation in primary T-cell acute lymphoblastic leukemia (T-ALL) cells, a process occurring within minutes. This suggests that CXCL12-mediated H3K9 methylation alters the overall chromatin conformation of T-ALL cells, enhancing their nuclear deformation and migratory capabilities [101]. The CXCL12/CXCR4/STAT3 pathway can promote the expression of IL-10 in chronic lymphocytic leukemia (CLL) cells, inducing immunosuppression. Treatment with lenalidomide, an immunomodulatory anticancer drug, inhibits STAT3Y705 phosphorylation induced with IL-10 in healthy T cells, reversing the T-cell dysfunction induced with CLL [102], indicating the significance of CXCL12/CXCR4 in leukemia treatment. Concurrently, research by Sison EA et al. has shown that chemotherapy for acute lymphoblastic leukemia (ALL) upregulates CXCR4 expression in all cells. Co-culturing ALL cells with bone marrow stromal cells (capable of producing CXCL12) generates drug resistance and protects ALL cells from chemotherapy-induced apoptosis, leading to further exacerbation of leukemia [103]. In summary, various studies suggest that CXCR4 inhibition may be beneficial for the treatment of ALL, yet further research is required to fully understand the comprehensive effects of targeting the leukemia microenvironment.
3.2. CXCL12/CXCR4/ACKR3 and STAT3 Pathway Crosstalk in Other Diseases
The CXCL12/CXCR4/ACKR3 signaling axis and the STAT3 pathway play crucial roles in inflammation and autoimmune diseases. In various diseases, the CXCL12/STAT3 signaling cascade exhibits diverse functions, both beneficial and malignant. For instance, in Toxoplasma-induced brain injury, elevated mRNA expression of CXCL12 and STAT3, in conjunction with other molecules, orchestrates innate immunity and T cell-mediated protective responses, effectively controlling cerebral Toxoplasma infection [104]. Additionally, cardiac stem cells, through paracrine secretion of CXCL12 and activation of the STAT3 pathway, ameliorate acute myocardial function following ischemia [105]. These stem cells contribute to post-ischemic myocardial recovery by mobilizing to injured tissue and enhancing local vascular regeneration after myocardial infarction, thereby mediating cardiac protection [106]. In cardiac myocytes, the miR-135a-5p/CXCL12/JAK2-STAT signaling axis promotes post-myocardial infarction inflammation and apoptosis, ultimately mitigating the extent of myocardial injury [107]. Furthermore, in the context of local vascular injury following arterial damage, the JAK/STAT signaling pathway induces the upregulation of vascular CXCL12 expression, promoting proliferative responses during vascular remodeling [108].
CXCL12 is also highly expressed in dermal cells, and during the process of wound healing, overexpressed IL-10 activates the STAT3 pathway, promoting fibroblast-specific hyaluronic acid synthesis and facilitating tissue regeneration. Furthermore, IL-10 enhances the expression of CXCL12 and VEGF in fibroblasts [109,110]. In studies related to skin-associated diseases, the activation of CXCR4 by CXCL12 was found to promote the phosphorylation of STAT3 and STAT5 in the dermal papilla layer and outer root sheath cells, thereby inhibiting hair growth [76].
Zhuang et al. discovered that in Helicobacter pylori-associated gastritis, the E-box basic helix–loop–helix protein 40 (BHLHE40) translocates to the cell nucleus and interacts with p-STAT3 (Tyr705), which is activated by the cagA protein. This interaction results in an upregulation of CXCL12 expression. The CXCL12, released by gastric epithelial cells, promotes CD4+ T cell infiltration in the gastric mucosa [111]. BHLHE40, a member of the basic helix–loop–helix transcription factor family, has been reported to play a crucial role in inflammatory diseases. Notably, in H. pylori-infected vascular endothelial cells, BHLHE40 expression is induced via the CagA/ERK pathway [111].
In a study conducted in 2020 on renal fibrosis, it was found that the binding of CXCL12α to CXCR4 instigates the phosphorylation of JAK2 and JAK3. This event triggers the activation of STAT3 and STAT6. The activation of these proteins subsequently inhibits the transcription of GSK3β and facilitates the assembly of the β-catenin degradation complex. As a result, β-catenin is liberated from the cytoplasmic degradation complex, leading to its accumulation in the nucleus. The nuclear β-catenin then mediates renal tubular cell fibrosis by interacting with the transcription factor TCF/LEF [112]. In a related study, Wang et al. provided evidence that in the context of diabetic kidney injury, the CXCL12α/CXCR4/STAT3 signaling pathway mediates the upregulation of DPP4-induced mitochondrial dysfunction in renal tubular cells. This finding underscores the critical role of this signaling pathway in renal pathology [113].
3.3. Mechanism of STAT3 Pathway Activation by CXCL12/CXCR4/ACKR3 Axis
In 1999, it was initially observed that the dimerization of CXCR4 is induced upon its binding to its ligand CXCL12, which in turn activates the JAK/STAT signaling pathway [114]. Further investigations have unveiled that the activation of JAK2 is a result of CXCL12 binding to CXCR4, which subsequently leads to the activation of the STAT3 signaling pathway [9,115] (Figure 1). Ahr et al. provided compelling evidence that two distinct structural domains of CXCR4 are involved in JAK2/STAT3 signal transduction. More specifically, the N-terminal portion of the third intracellular loop (ICL3) of CXCR4 is responsible for the activation of JAK2, which results in the phosphorylation of tyrosine at residue 157 (Tyr157) within the C-terminal portion of the second intracellular loop (ICL2). This event ultimately leads to the activation of the STAT3 pathway [116]. This observation underscores the intricate molecular mechanisms involved in signal transduction pathways.
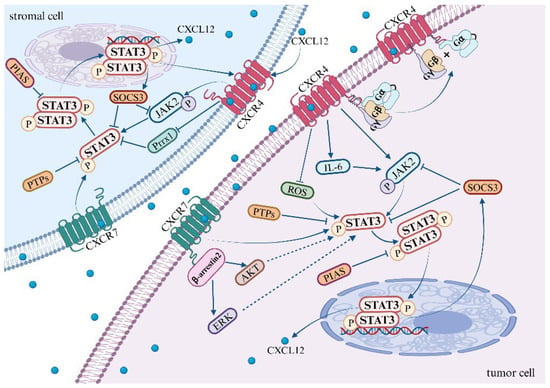
Figure 1.
Mechanisms of the STAT3 pathway activation by CXCL12/CXCR4/ACKR3 axis.
CXCR4 activates the JAK2/STAT3 pathway through the association of heterotrimeric G proteins with GPCRs [117]. The precise mechanistic underpinnings of this association remain to be elucidated. However, Soriano SF et al. discovered that the binding of heterotrimeric G proteins to G protein-coupled receptors (GPCRs) is contingent upon JAK activation. This process involves the upregulation of the suppressor of cytokine signaling (SOCS3), which, by binding to CXCR4, impedes the JAK/STAT and Gi pathways without disrupting the expression of chemokine receptors on the cell surface [115]. Existing research suggests that the activation of CXCL12/CXCR4 leads to an upregulation in the expression of SOCS3 within the STAT3 pathway. Subsequently, SOCS3 binds to CXCR4, providing a negative feedback mechanism that inhibits the STAT3 pathway [55,56,115,118]. It is noteworthy that the JAK2 inhibitor AG490 mitigates the upregulation of SOCS3, whereas the G protein inhibitor pertussis toxin (PTX) does not exert any influence on the expression of SOCS3.
Kim et al. uncovered that the upregulation of CXCR4, when stimulated by AKT and JNK, fosters the expression and secretion of IL-6. Following this, IL-6 activates the JAK/STAT3/Snail signaling axis, which leads to the depletion of the nuclear protein UHRF1, thereby inducing EMT in liver cancer cells [119]. Additionally, CXCR4 may activate the STAT3 pathway by inhibiting the production of reactive oxygen species (ROS). Under hypoxic conditions, ROS are known to mediate the activation of the STAT3 pathway. Notably, in tumor-targeted therapy, both curcumin and a novel dimanganese complex (FMSP) have been observed to downregulate CXCR4, thereby promoting the generation of ROS and inhibiting the activation of the STAT3 pathway [120,121]. Furthermore, the vascular endothelial growth factor (VEGF) may mediate the activation of CXCR4 in the STAT3 pathway. Clinical samples from lung cancer patients have demonstrated the co-expression of CXCR4, p-STAT3, and VEGF-A, suggesting a potential role in tumor progression and angiogenesis in non-small cell lung cancer (NSCLC) [122].
In addition to the aforementioned pathways, several other signaling pathways have been reported to activate the STAT3 pathway. The binding of vascular endothelial growth factor (VEGF) and epidermal growth factor (EGF) to their respective receptors instigates receptor dimerization. The phosphorylation of these dimerized receptors occurs reciprocally, with VEGFR inducing JAK2 phosphorylation and EGFR inducing c-Src kinase phosphorylation, thereby facilitating the activation of the STAT3 pathway [123]. Moreover, mTORC1 has also been found to activate the STAT3 pathway, thereby promoting angiogenesis [124]. Apart from IL-6, other members of the interleukin family, such as IL-10, IL-17, IL-19, and IL-22, are capable of activating the STAT3 pathway [125,126,127,128]. Although there is currently no definitive research demonstrating whether CXCR4 activates STAT3 through these pathways, further investigation may elucidate the specific mechanisms underlying CXCR4-mediated STAT3 activation.
ACKR3, identified in 2005, functions as an additional specific receptor for CXCL12 [129]. Current research has shown that the activation of CXCL12/ACKR3 contributes to the progression of certain diseases and tumors by facilitating STAT3 signaling. However, the exact mechanism through which ACKR3 activates the STAT3 pathway remains to be comprehensively elucidated. In studies pertaining to ACKR3, it has been found that the binding of CXCL12 to ACKR3 recruits β-arrestin2, which subsequently activates the AKT and ERK signaling pathways [8,130]. Notably, the activation of either the AKT or ERK pathway has been linked with enhanced STAT3 transcriptional activity [90,131], providing valuable insights for future investigations into the mechanisms underlying STAT3 pathway activation.
In addition to its interaction with its cognate receptor, CXCL12 has the ability to modulate transcriptional co-activators, thereby promoting the transcriptional activation of STAT3. In the pre-metastatic microenvironment of hepatocellular carcinoma, the expression of CXCL12 can downregulate the transcription factor Paired Related Homeobox 1 (Prrx1), thereby inducing an increase in the protein and mRNA expression of CXCR4 in hepatocellular carcinoma cells via the STAT3 pathway [91]. Specifically, the binding of CXCL12 to CXCR4, under the mediation of Prrx1, promotes the nuclear translocation and phosphorylation level of STAT3, activates the STAT3 signaling pathway, and upregulates the expression of CXCR4. In the pre-metastatic niche, CXCL12 recruits circulating tumor cells with high CXCR4 and low Prrx1 expression from the bloodstream, ultimately promoting the metastatic colonization of distant organs [91].
3.4. Mechanism of STAT3 Signaling Pathway Upregulation of CXCL12 Expression
The expression of CXCL12 is also modulated by the STAT3 signaling pathway. Activated STAT3, by upregulating CXCL12, promotes tumor cell migration. Immunosuppressive cytokines (such as IL-10, PD-L1, and TGF-β) activate the STAT3 signaling pathway, leading to tumor immune suppression and epithelial–mesenchymal transition. Consequently, CXCL12 expression is enhanced in primary tumors [132]. In related studies involving fibroblasts, both aging and p53 mutations activate the STAT3 pathway in fibroblasts, leading to increased secretion of CXCL12. This induction of CXCL12 contributes to the acquisition of cancer-associated fibroblast (CAF) characteristics, ultimately promoting tumor growth and metastasis [133,134].
In bone marrow stromal cells, increased expression of heme oxygenase-1 (HO-1) facilitates JAK2/STAT3 phosphorylation, subsequently promoting the secretion of CXCL12 and activation of the CXCL12/CXCR4 signaling pathway [135]. This molecular cascade is implicated in various signal transduction pathways related to cell survival and proliferation. Although several studies suggest that the STAT3 pathway upregulates CXCL12 expression, the precise regulatory mechanisms remain elusive.
In the context of other diseases, it has been found that p-STAT3 binds to the CXCL12 promoter to enhance its expression. Li ZY and colleagues discovered that p-STAT3 can directly bind to a 163-bp fragment of the CXCL12 promoter from −1608 to −1770, promoting the expression of CXCL12 in the dorsal root ganglia (DRG), leading to chronic pain after oxaliplatin treatment [136]. Research by Xin WJ and others showed that long-term morphine treatment increased the binding of p-STAT3 to the specific binding site (−1667/−1685) of the CXCL12 promoter [137] and that p21 inhibited the binding of STAT3 to the CXCL12 promoter [108]. At the same time, p-STAT3 regulates histone modifications in the CXCL12 promoter region. It enhances the interaction between the acetyltransferase p300 and STAT3, inducing histone H4 acetylation in the CXCL12 promoter region, which promotes the expression of CXCL12 [137].
The STAT3 pathway has the capacity to modulate other transcription factors, thereby enhancing CXCL12 expression. Kim et al. revealed that in normal fibroblasts, the activation of the IL-6/STAT3 pathway leads to an upregulation of Twist1. This upregulation subsequently augments CXCL12 expression, thereby facilitating the transformation of normal fibroblasts into cancer-associated fibroblasts (CAFs) [138]. Furthermore, it has been observed that activated STAT3 can induce a phenotypic transition in tumor-associated macrophages (TAMs) from M1 to M2 via NF-κB signaling within the tumor microenvironment (TME). This transition results in immune suppression and the induction of EMT in colorectal cancer, thereby promoting the expression of CXCL12 in primary tumors [132].
4. Interplay between the CXCL12/CXCR4/ACKR3 Axis and the STAT3 Pathway in Coordination with Other Signaling Pathways
Tumor growth and metastasis represent a multifaceted process encompassing multiple stages that involve the regulation of a variety of growth factors and signaling pathways [139]. Upon interaction with its specific receptors, CXCL12, secreted by either stromal cells or tumor cells, has the potential to activate distinct signaling pathways in various environments. These pathways often demonstrate parallel or intersecting relationships (Figure 2). Notably, research on prostate cancer has identified that CXCL12 secreted by stromal cells can stimulate the invasive characteristics of prostate cancer cells, potentially playing a significant role in tumor progression associated with obesity [140]. Further mechanistic investigations suggest that the binding of CXCL12 to its receptors concurrently activates the STAT3, NFκB, and MAPK signaling pathways [140]. However, the reciprocal regulatory interactions among STAT3, NFκB, and MAPK induced by CXCL12 are yet to be fully elucidated. Jin et al. have documented the regulatory effects of the MAPK/STAT3/NFκB signaling cascade in the treatment of gastric cancer cells [141,142].
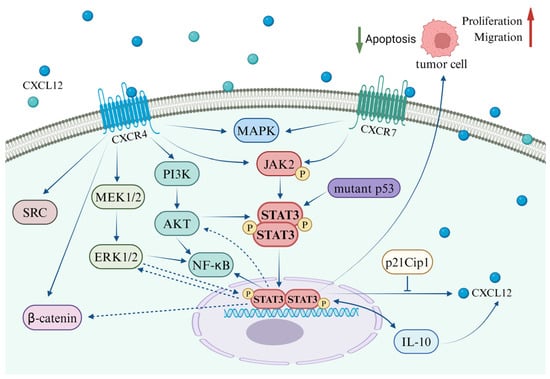
Figure 2.
Interplay between the CXCL12/CXCR4/ACKR3 axis and the STAT3 pathway in coordination with other signaling pathways.
In a variety of tumor cells, including gastric cancer, glioblastoma, and breast cancer, the activation of the CXCL12/CXCR4 axis has been observed to stimulate the SRC, ERK, and STAT3 pathways. This stimulation promotes tumor cell migration and imparts anti-apoptotic capabilities [143]. In a similar vein, the CXCL12/STAT3 axis has been implicated in research related to brain injury. In a rat model of middle cerebral artery occlusion (MCAO), the administration of oncostatin M (OSM) was found to significantly upregulate CXCL12 in bone marrow-derived mesenchymal stem cells (BMSCs) via the STAT3 and ERK pathways. Moreover, OSM notably enhanced the expression of VEGF and MMP-2. OSM also facilitated the secretion of CXCL12 in astrocytes through the STAT3 and ERK pathways, thereby augmenting BMSC migration [144]. Liang et al. demonstrated that IL-6 activates the Wnt/β-catenin pathway via STAT3/ERK signaling, thereby promoting EMT in colorectal cancer cells [145]. These findings offer valuable insights into the interplay between the CXCL12-mediated STAT3 pathway and the ERK and Wnt/β-catenin pathways, thereby presenting a promising direction for future research.
Cancer stem cells (CSCs) are predominantly responsible for chemotherapy resistance and tumor recurrence. Research has demonstrated that when co-cultured in vitro with human bone marrow-derived mesenchymal stem cells, CD15-induced CSCs (iCSCs) isolated from 293FT cells exhibit characteristics akin to tumor stem cells and proliferate effectively. Subsequent investigations have discovered that iCSCs express the chemokine CXCL12 and its receptor CXCR4, which activate the Fut4 gene via the CXCR4/ERK/ELK-1 signaling pathway, thereby maintaining iCSCs in an undifferentiated state through the CXCR4/AKT/STAT3 signal [90]. Mesenchymal stem cells (MSCs) possess therapeutic potential for the repair of damaged tissues. In studies pertaining to skin wound healing, it has been observed that MSC exosomes activate the AKT, ERK, and STAT3 signaling pathways and induce the expression of CXCL12 [146]. Bone marrow mesenchymal stem cells (BMSCs) demonstrate potential regenerative effects on brain damage, and it has been established that oncostatin M (OSM) influences the proliferation and migration of mesenchymal stem cells.
5. Targeted Therapy for the CXCL12/CXCR4/ACKR3 and STAT3 Pathways
Given the deleterious role that the CXCL12/STAT3 axis plays in a variety of tumors and diseases, it is of utmost importance to target this axis for therapeutic intervention in tumors. Pharmacological intervention serves as the cornerstone in the treatment of cancer. Notably, within the realm of tumor metastasis, treatment with vitamin D has been demonstrated to selectively inhibit the expression of p-STAT3, zinc finger E-box binding homeobox 1 (Zeb1), and Vimentin while enhancing the expression of E-Cadherin. Intriguingly, in vivo studies further substantiate that a deficiency in vitamin D results in an upregulation of Zeb1 and p-STAT3 expression in primary breast tumor cells, thereby augmenting the expression of CXCL12 within the pulmonary stroma [147]. Furthermore, in the context of lung metastasis, a deficiency in vitamin D facilitates the co-localization of CXCL12 and its receptor CXCR4 [147].
Pancreatic ductal adenocarcinoma (PDAC), one of the most lethal forms of cancer, is distinguished by the presence of highly tumorigenic and metastatic cancer stem cells (CSCs). Treatment with the antimalarial drug chloroquine in vitro has been observed to significantly diminish CSCs, thereby reducing in vivo tumorigenicity and invasiveness across a wide range of pancreatic cancers [148]. The combination of gemcitabine with in vivo therapy results in more effective tumor eradication and an improved overall survival rate. From a mechanistic perspective, the inhibitory effects of chloroquine are attributed to the suppression of the CXCL12/CXCR4 signaling pathway, leading to a reduction in the phosphorylation of ERK and STAT3 [148]. Furthermore, SKLB-850, a novel SYK inhibitor, has been found to markedly suppress proliferation and induce apoptosis in B-cell lymphomas (BCLs). Further mechanistic studies reveal that SKLB-850 effectively inhibits the activation of the Syk/ERK, Src/FAK, and JAK2/STAT3 pathways, while significantly reducing the secretion of chemokines CCL3, CCL4, and CXCL12 [149].
Traditional Chinese medicine (TCM) plays a pivotal role in modulating the tumor microenvironment (TME), which includes reshaping the immunosuppressive microenvironment, hypoxic conditions, angiogenesis/lymphangiogenesis, and the extracellular matrix [150]. The Qingre Huoxue (QRHX) formula, a 1:1 mixture (w/w) of Paeonia lactiflora (Shaoyao) and Scutellaria baicalensis (Huangqin), has garnered increasing recognition for its anticancer and anti-inflammatory properties [151]. Notably, QRHX inhibits inflammation and the CXCL12/CXCR4/JAK2/STAT3 signaling pathway, thereby modulating tumor-associated macrophages in murine models and suppressing tumor growth [152]. Additionally, thymoquinone (TQ), the major active component of Nigella glandulifera (Huixiangye Heizhongcao), inhibits the growth of multiple myeloma cell lines. Mechanistically, it suppresses CXCL12-mediated actin polymerization and cell proliferation, significantly reduces STAT3 phosphorylation in multiple myeloma cells, and downregulates Bcl-2 and Bcl-XL expression [153]. Vitexin, a glycosylated flavonoid found in various medicinal plants, exhibits diverse pharmacological activities [154]. It downregulates DNA-binding capacity, reduces nuclear translocation of STAT3, and attenuates epidermal growth factor (EGF)-driven STAT3 gene expression. Furthermore, it inhibits the CXCL12-induced invasion of liver cancer cells [155]. Sinomenine, an alkaloid isolated from traditional Chinese herbs, has been successfully used for centuries in treating rheumatoid arthritis [156]. Notably, sinomenine has gained attention for its antitumor potential. It has been demonstrated to inhibit proliferation and induce apoptosis in various human tumor cells [157,158,159,160]. Ye et al. found that sinomenine induces S-phase arrest, inhibits invasion and metastasis in osteosarcoma cells by suppressing the CXCR4-STAT3 pathway, and mitigates bone destruction and angiogenesis mediated by osteoclastogenesis in osteosarcoma [161]. Turmeric (Curcuma longa), a perennial herb of the ginger family, has shown anticancer potential in a 7,12-dimethylbenz[a]anthracene (DMBA)-induced carcinogenesis hamster model [162]. Aggarwal et al. discovered that turmeric downregulates CXCR4 expression and inhibits tumor cell proliferation by suppressing the STAT3 pathway [121].
Chimeric antigen receptor T cells (CAR-T) specific to Claudin182 (CLDN18.2) have demonstrated therapeutic efficacy in CLDN18.2-positive pancreatic ductal adenocarcinoma (PDAC). The presence of CXCR4 has been observed to enhance the infiltration of CAR-T cells, thereby enabling CXCR4 CAR-T cells to more abundantly infiltrate tumor sites. Through the STAT3/NF-κB/CXCL12α axis, these cells inhibit the migration of myeloid-derived suppressor cells, thus achieving enhanced therapeutic effects in the treatment of CLDN18.2-positive pancreatic cancer [163]. Synthetic triterpenoids of the oleanane type, including 2-cyano-3,12-dioxooleana-1,9(11)-dien-28-oic acid methyl ester, are emerging as promising pharmaceuticals for the prevention and treatment of breast cancer. These compounds have been shown to inhibit the proliferation of estrogen receptor (ER)-positive breast cancer cells both in vitro and in vivo [164,165,166]. Synthetic triterpenoids have been found to inhibit the infiltration of tumor-associated macrophages into the mammary glands of PyMT mice, reduce the levels of chemokines CXCL12 and CCL2 in primary PyMT mammary tumor cells, and inhibit the phosphorylation of STAT3, thereby suppressing cell proliferation [167].
The CXCL12/CXCR4/ACKR3 axis plays a pivotal role in regulating the migration, dissemination, and homing of leukemia cells. ACKR3 is markedly overexpressed in ALL cells compared to hematopoietic stem cells or normal hematopoietic progenitor cells [168]. MicroRNA-101 (miR-101) has emerged as a novel T-ALL suppressor targeting ACKR3. Exogenous overexpression of miR-101 in leukemia cells inhibited T-ALL cell proliferation and invasion in vitro, and xenograft mouse models demonstrated that miR-101 overexpression suppressed tumor growth and lung metastasis in vivo. Mechanistically, miR-101 suppresses T-ALL tumor development by targeting the CXCL12/ACKR3/STAT3 signaling pathway [86]. T140 is a CXCR4-specific inhibitor. Burger et al. reported that T140 and its analogs inhibited the activity, chemotaxis, and migration of CLL cells in the bone marrow stroma, with the mechanism involving the disruption of CXCL12-induced STAT3 and MAPK phosphorylation [169] (Table 1).

Table 1.
Compendium of therapeutic agents targeting the CXCL12/STAT3 pathway.
At present, several CXCR4 inhibitors have received FDA approval for clinical use. These include Plerixafor (also known as AMD-3100) [170], Motixafortide [171], and Mavorixafor [172]. Research findings indicate that Plerixafor is capable of inhibiting the activation of the STAT3 signaling pathway, thereby suppressing the development of breast cancer and prostate cancer [52,140]. Furthermore, when Plerixafor is used in combination with triptolide, it reduces the growth of osteosarcoma and inhibits lung metastasis. Notably, the phosphorylation levels of STAT3 decrease in osteosarcoma cells treated with this combination therapy [173]. Additionally, in the context of chronic kidney disease, the CXCR4 inhibitor Plerixafor shows promise as a targeted therapy for renal fibrosis by inhibiting the STAT3 pathway [112]. However, the relationship between Motixafortide and Mavorixafor and the STAT3 pathway remains to be elucidated.
6. Discussion
The chemokine CXCL12 and STAT3 signaling pathways play pivotal roles in the development and metastasis of tumors. Therapies targeting tumors, driven by the activation of the STAT3 signaling pathway by CXCL12, warrant considerable attention. A multitude of studies have demonstrated that CXCL12 fosters tumor growth and metastasis, and pharmacological interventions targeting the STAT3 pathway have proven efficacious for various cancers. This review endeavors to elucidate the reciprocal regulatory interaction between CXCL12 and STAT3, thereby addressing the deleterious effects engendered by various tumors.
The CXCL12/CXCR4/ACKR3 axis exhibits an activating interaction with the STAT3 pathway, which contributes to the malignant progression of tumors. However, no study has elucidated the specific molecular mechanisms by which the CXCL12/CXCR4/ACKR3 axis activates the JAK2/STAT3 pathway, and responses to the CXCL12/STAT3 signal transduction may vary across different tumor types and cell lines. The interaction between CXCL12 and its receptors, CXCR4 or ACKR3, plays a crucial role in hematopoietic stem cell proliferation and leukemogenesis. Reviews have highlighted that the CXCL12/CXCR4/ACKR3 axis can activate the STAT3 pathway, leading to leukemia development. Although several drugs (such as Plerixafor, BKT140, and NOX-A12.) targeting CXCL12/CXCR4 have been developed for leukemia treatment, it is essential to recognize that any aberrations (such as intrinsic genetic variations or external factors disrupting homeostasis) may contribute to leukemia initiation. Beyond conferring survival advantages to malignant cells, the activated CXCL12/CXCR4/ACKR3 axis also enhances the invasiveness of hematological malignancies by regulating cell migration and conferring resistance to chemotherapy. Additionally, the potential for leveraging the upregulation of CXCL12 and the subsequent activation of the STAT3 pathway to prevent tumor metastasis warrants further investigation. The present research is largely focused on elucidating the role of CXCL12/STAT3 signaling, yet there is a lack of clear therapeutic strategies. A pivotal issue is how to intervene in the CXCL12/CXCR4/STAT3 signaling pathway to achieve therapeutic effects.
The genesis and progression of tumors are influenced by a myriad of factors, with diverse cytokines and signaling pathways interacting within the tumor. The crosstalk mechanism between CXCL12 and STAT3 varies in different contexts. Initially, CXCL12 was identified as binding with its receptor CXCR4 to influence the STAT3 pathway. However, subsequent to the confirmation of ACKR3 as a receptor of CXCL12, a plethora of studies have verified that the binding of CXCL12 and ACKR3 can also activate the STAT3 pathway, thereby fostering tumor development. Moreover, an abundance of research has also substantiated that the activated STAT3 signaling pathway can promote the secretion of CXCL12 in both stromal cells and tumor cells. Regrettably, no study has yet demonstrated a mutual feedback activation mechanism between CXCL12 and STAT3 in the same cellular context, posing a formidable challenge for future research. Nevertheless, several drug treatments have proven effective against the CXCL12/STAT3 pathway; traditional Chinese medicine, in particular, offers promising prospects. Furthermore, specific targeted intervention of the related proteins in the crosstalk signal axis of CXCL12/STAT3 can pave new avenues for cancer treatment.
Funding
This research was funded by NSFC (82360548), the Basic Research Program of the Science and Technology Department of Yunnan Province (202301ATO70466 and KUST-PE2022019Y).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Murphy, P.M.; Heusinkveld, L. Multisystem multitasking by CXCL12 and its receptors CXCR4 and ACKR3. Cytokine 2018, 109, 2–10. [Google Scholar]
- Righetti, A.; Giulietti, M.; Šabanović, B.; Occhipinti, G.; Principato, G.; Piva, F. CXCL12 and Its Isoforms: Different Roles in Pancreatic Cancer? J. Oncol. 2019, 2019, 9681698. [Google Scholar] [CrossRef]
- Yu, L.; Cecil, J.; Peng, S.B.; Schrementi, J.; Kovacevic, S.; Paul, D.; Su, E.W.; Wang, J. Identification and expression of novel isoforms of human stromal cell-derived factor 1. Gene 2006, 374, 174–179. [Google Scholar] [CrossRef] [PubMed]
- Dettin, M.; Pasquato, A.; Scarinci, C.; Zanchetta, M.; De Rossi, A.; Di Bello, C. Anti-HIV activity and conformational studies of peptides derived from the C-terminal sequence of SDF-1. J. Med. Chem. 2004, 47, 3058–3064. [Google Scholar] [CrossRef] [PubMed]
- Janowski, M. Functional diversity of SDF-1 splicing variants. Cell Adhes. Migr. 2009, 3, 243–249. [Google Scholar] [CrossRef]
- Laguri, C.; Sadir, R.; Rueda, P.; Baleux, F.; Gans, P.; Arenzana-Seisdedos, F.; Lortat-Jacob, H. The novel CXCL12gamma isoform encodes an unstructured cationic domain which regulates bioactivity and interaction with both glycosaminoglycans and CXCR4. PLoS ONE 2007, 2, e1110. [Google Scholar] [CrossRef]
- Loetscher, M.; Geiser, T.; O’reilly, T.; Zwahlen, R.; Baggiolini, M.; Moser, B. Cloning of a human seven-transmembrane domain receptor, LESTR, that is highly expressed in leukocytes. J. Biol. Chem. 1994, 269, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, E.L.; Lee, W.; Lu, J.; Lowy, A.M.; Kim, J. Chemokine CXCL12 activates dual CXCR4 and CXCR7-mediated signaling pathways in pancreatic cancer cells. J. Transl. Med. 2012, 10, 68. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.F.; Wang, J.F.; Matczak, E.; Proper, J.A.; Groopman, J.E. Janus kinase 2 is involved in stromal cell-derived factor-1alpha-induced tyrosine phosphorylation of focal adhesion proteins and migration of hematopoietic progenitor cells. Blood 2001, 97, 3342–3348. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.Y.; Gao, Z.H.; Chu, J.H.; Han, X.Z.; Qu, X.J. Downregulation of the CXCR4/CXCL12 axis blocks the activation of the Wnt/beta-catenin pathway in human colon cancer cells. Biomed. Pharmacother. 2015, 71, 46–52. [Google Scholar] [CrossRef]
- Hu, X.M.; Zhang, H.; Xu, H.; Zhang, H.L.; Chen, L.P.; Cui, W.Q.; Yang, W.; Shen, W. Chemokine receptor CXCR4 regulates CaMKII/CREB pathway in spinal neurons that underlies cancer-induced bone pain. Sci. Rep. 2017, 7, 4005. [Google Scholar] [CrossRef]
- Goldsmith, Z.G.; Dhanasekaran, D.N. G protein regulation of MAPK networks. Oncogene 2007, 26, 3122–3142. [Google Scholar]
- Gupta, S.K.; Lysko, P.G.; Pillarisetti, K.; Ohlstein, E.; Stadel, J.M. Chemokine receptors in human endothelial cells. Functional expression of CXCR4 and its transcriptional regulation by inflammatory cytokines. J. Biol. Chem. 1998, 273, 4282–4287. [Google Scholar] [CrossRef]
- Gerits, N.; Kostenko, S.; Shiryaev, A.; Johannessen, M.; Moens, U. Relations between the mitogen-activated protein kinase and the cAMP-dependent protein kinase pathways: Comradeship and hostility. Cell. Signal. 2008, 20, 1592–1607. [Google Scholar]
- Atkins, M.; Wurmser, M.; Darmon, M.; Roche, F.; Nicol, X.; Metin, C. CXCL12 targets the primary cilium cAMP/cGMP ratio to regulate cell polarity during migration. Nat. Commun. 2023, 14, 8003. [Google Scholar] [CrossRef]
- Naumann, U.; Cameroni, E.; Pruenster, M.; Mahabaleshwar, H.; Raz, E.; Zerwes, H.G.; Rot, A.; Thelen, M. CXCR7 functions as a scavenger for CXCL12 and CXCL11. PLoS ONE 2010, 5, e9175. [Google Scholar] [CrossRef]
- Rajagopal, S.; Kim, J.; Ahn, S.; Craig, S.; Lam, C.M.; Gerard, N.P.; Gerard, C.; Lefkowitz, R.J. Beta-arrestin- but not G protein-mediated signaling by the “decoy” receptor CXCR7. Proc. Natl. Acad. Sci. USA 2010, 107, 628–632. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, M.; Li, Y.; Xu, D.; Wang, Y.; Song, A.; Zhu, B.; Huang, Y.; Zheng, J.C. CXCR7 Mediates Neural Progenitor Cells Migration to CXCL12 Independent of CXCR4. Stem Cells 2015, 33, 2574–2585. [Google Scholar] [CrossRef]
- Xu, L.; Li, C.; Hua, F.; Liu, X. The CXCL12/CXCR7 signalling axis promotes proliferation and metastasis in cervical cancer. Med. Oncol. 2021, 38, 58. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.H.; Lloyd, A.R. Chemokines: Leucocyte recruitment and activation cytokines. Lancet 1997, 349, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.; Saha, A.; Clark, R.; Kolonin, M.G.; Digiovanni, J. CXCR4 and CXCR7 signaling promotes tumor progression and obesity-associated epithelial-mesenchymal transition in prostate cancer cells. Oncogene 2022, 41, 4633–4644. [Google Scholar]
- Antonello, P.; Pizzagalli, D.U.; Foglierini, M.; Melgrati, S.; Radice, E.; Thelen, S.; Thelen, M. ACKR3 promotes CXCL12/CXCR4-mediated cell-to-cell-induced lymphoma migration through LTB4 production. Front. Immunol. 2022, 13, 1067885. [Google Scholar] [CrossRef]
- Lu, X.; Wu, Y.; Cao, R.; Yu, X.; Gong, J. CXCL12 secreted by pancreatic stellate cells accelerates gemcitabine resistance of pancreatic cancer by enhancing glycolytic reprogramming. Anim. Cells Syst. 2022, 26, 148–157. [Google Scholar] [CrossRef]
- Fang, X.; Hong, Y.; Dai, L.; Qian, Y.; Zhu, C.; Wu, B.; Li, S. CRH promotes human colon cancer cell proliferation via IL-6/JAK2/STAT3 signaling pathway and VEGF-induced tumor angiogenesis. Mol. Carcinog. 2017, 56, 2434–2445. [Google Scholar] [CrossRef]
- Yu, H.; Kortylewski, M.; Pardoll, D. Crosstalk between cancer and immune cells: Role of STAT3 in the tumour microenvironment. Nat. Rev. Immunol. 2007, 7, 41–51. [Google Scholar] [CrossRef]
- Yu, H.; Pardoll, D.; Jove, R. STATs in cancer inflammation and immunity: A leading role for STAT3. Nat. Rev. Cancer 2009, 9, 798–809. [Google Scholar]
- Bromberg, J. Stat proteins and oncogenesis. J. Clin. Investig. 2002, 109, 1139–1142. [Google Scholar] [CrossRef]
- Dorritie, K.A.; Mccubrey, J.A.; Johnson, D.E. STAT transcription factors in hematopoiesis and leukemogenesis: Opportunities for therapeutic intervention. Leukemia 2014, 28, 248–257. [Google Scholar]
- Takeda, K.; Noguchi, K.; Shi, W.; Tanaka, T.; Matsumoto, M.; Yoshida, N.; Kishimoto, T.; Akira, S. Targeted disruption of the mouse Stat3 gene leads to early embryonic lethality. Proc. Natl. Acad. Sci. USA 1997, 94, 3801–3804. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.P.; Ghoreschi, K.; Steward-Tharp, S.M.; Rodriguez-Canales, J.; Zhu, J.; Grainger, J.R.; Hirahara, K.; Sun, H.W.; Wei, L.; Vahedi, G.; et al. Opposing regulation of the locus encoding IL-17 through direct, reciprocal actions of STAT3 and STAT5. Nat. Immunol. 2011, 12, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Sgrignani, J.; Garofalo, M.; Matkovic, M.; Merulla, J.; Catapano, C.V.; Cavalli, A. Structural Biology of STAT3 and Its Implications for Anticancer Therapies Development. Int. J. Mol. Sci. 2018, 19, 1591. [Google Scholar] [CrossRef]
- Bournazou, E.; Bromberg, J. Targeting the tumor microenvironment: JAK-STAT3 signaling. JAK-STAT 2013, 2, e23828. [Google Scholar] [CrossRef]
- Okitsu, K.; Kanda, T.; Imazeki, F.; Yonemitsu, Y.; Ray, R.B.; Chang, C.; Yokosuka, O. Involvement of interleukin-6 and androgen receptor signaling in pancreatic cancer. Genes Cancer 2010, 1, 859–867. [Google Scholar] [CrossRef]
- Thoennissen, N.H.; Iwanski, G.B.; Doan, N.B.; Okamoto, R.; Lin, P.; Abbassi, S.; Song, J.H.; Yin, D.; Toh, M.; Xie, W.D.; et al. Cucurbitacin B induces apoptosis by inhibition of the JAK/STAT pathway and potentiates antiproliferative effects of gemcitabine on pancreatic cancer cells. Cancer Res. 2009, 69, 5876–5884. [Google Scholar] [CrossRef]
- Zimmers, T.A.; Fishel, M.L.; Bonetto, A. STAT3 in the systemic inflammation of cancer cachexia. Semin. Cell Dev. Biol. 2016, 54, 28–41. [Google Scholar]
- Shen, H.; Guo, M.; Wang, L.; Cui, X. MUC16 facilitates cervical cancer progression via JAK2/STAT3 phosphorylation-mediated cyclooxygenase-2 expression. Genes Genom. 2020, 42, 127–133. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, Y.; Wang, S.; Shen, Q.; Zhou, X. The role of STAT3 in leading the crosstalk between human cancers and the immune system. Cancer Lett. 2018, 415, 117–128. [Google Scholar]
- Sun, Q.; Zhao, X.; Li, R.; Liu, D.; Pan, B.; Xie, B.; Chi, X.; Cai, D.; Wei, P.; Xu, W.; et al. STAT3 regulates CD8+ T cell differentiation and functions in cancer and acute infection. J. Exp. Med. 2023, 220, e20220686. [Google Scholar]
- Li, F.; Gao, L.; Jiang, Q.; Wang, Z.; Dong, B.; Yan, T.; Chen, X. Radiation enhances the invasion abilities of pulmonary adenocarcinoma cells via STAT3. Mol. Med. Rep. 2013, 7, 1883–1888. [Google Scholar] [CrossRef]
- Hashimoto, S.; Hashimoto, A.; Muromoto, R.; Kitai, Y.; Oritani, K.; Matsuda, T. Central Roles of STAT3-Mediated Signals in Onset and Development of Cancers: Tumorigenesis and Immunosurveillance. Cells 2022, 11, 2618. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Jove, R. The STATs of cancer--new molecular targets come of age. Nat. Rev. Cancer 2004, 4, 97–105. [Google Scholar]
- Chang, Q.; Bournazou, E.; Sansone, P.; Berishaj, M.; Gao, S.P.; Daly, L.; Wels, J.; Theilen, T.; Granitto, S.; Zhang, X.; et al. The IL-6/JAK/Stat3 feed-forward loop drives tumorigenesis and metastasis. Neoplasia 2013, 15, 848–862. [Google Scholar] [CrossRef]
- Wang, T.; Niu, G.; Kortylewski, M.; Burdelya, L.; Shain, K.; Zhang, S.; Bhattacharya, R.; Gabrilovich, D.; Heller, R.; Coppola, D.; et al. Regulation of the innate and adaptive immune responses by Stat-3 signaling in tumor cells. Nat. Med. 2004, 10, 48–54. [Google Scholar] [CrossRef]
- Wang, Y.S.; Chen, C.; Zhang, S.Y.; Li, Y.; Jin, Y.H. (20S) Ginsenoside Rh2 Inhibits STAT3/VEGF Signaling by Targeting Annexin A2. Int. J. Mol. Sci. 2021, 22, 9289. [Google Scholar] [CrossRef]
- Iliopoulos, D.; Jaeger, S.A.; Hirsch, H.A.; Bulyk, M.L.; Struhl, K. STAT3 activation of miR-21 and miR-181b-1 via PTEN and CYLD are part of the epigenetic switch linking inflammation to cancer. Mol. Cell 2010, 39, 493–506. [Google Scholar] [CrossRef]
- Gozgit, J.M.; Bebernitz, G.; Patil, P.; Ye, M.; Parmentier, J.; Wu, J.; Su, N.; Wang, T.; Ioannidis, S.; Davies, A.; et al. Effects of the JAK2 inhibitor, AZ960, on Pim/BAD/BCL-xL survival signaling in the human JAK2 V617F cell line SET-2. J. Biol. Chem. 2008, 283, 32334–32343. [Google Scholar] [CrossRef]
- Bai, J.; Sui, J.; Demirjian, A.; Vollmer, C.M., Jr.; Marasco, W.; Callery, M.P. Predominant Bcl-XL knockdown disables antiapoptotic mechanisms: Tumor necrosis factor-related apoptosis-inducing ligand-based triple chemotherapy overcomes chemoresistance in pancreatic cancer cells in vitro. Cancer Res. 2005, 65, 2344–2352. [Google Scholar] [CrossRef]
- Zou, S.; Tong, Q.; Liu, B.; Huang, W.; Tian, Y.; Fu, X. Targeting STAT3 in Cancer Immunotherapy. Mol. Cancer 2020, 19, 145. [Google Scholar]
- Guo, J.; Tong, C.Y.; Shi, J.G.; Li, X.J. C-X-C motif chemokine ligand 12 (CXCL12)/C-X-C motif chemokine receptor 7(CXCR7) regulates epithelial-mesenchymal transition process and promotes the metastasis of esophageal cancer by activating signal transducer and activator of transcription 3 (STAT3) pathway. Bioengineered 2022, 13, 7425–7438. [Google Scholar] [PubMed]
- Wang, M.; Lin, T.; Wang, Y.; Gao, S.; Yang, Z.; Hong, X.; Chen, G. CXCL12 suppresses cisplatin-induced apoptosis through activation of JAK2/STAT3 signaling in human non-small-cell lung cancer cells. OncoTargets Ther. 2017, 10, 3215–3224. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, X.; Song, Y.; Si, M.; Sun, Y.; Liu, X.; Cui, S.; Qu, X.; Yu, X. Exosomal miR-146a-5p and miR-155-5p promote CXCL12/CXCR7-induced metastasis of colorectal cancer by crosstalk with cancer-associated fibroblasts. Cell Death Dis. 2022, 13, 380. [Google Scholar] [CrossRef]
- Liu, X.; Xiao, Q.; Bai, X.; Yu, Z.; Sun, M.; Zhao, H.; Mi, X.; Wang, E.; Yao, W.; Jin, F.; et al. Activation of STAT3 is involved in malignancy mediated by CXCL12-CXCR4 signaling in human breast cancer. Oncol. Rep. 2014, 32, 2760–2768. [Google Scholar] [CrossRef]
- Kershaw, N.J.; Murphy, J.M.; Liau, N.P.; Varghese, L.N.; Laktyushin, A.; Whitlock, E.L.; Lucet, I.S.; Nicola, N.A.; Babon, J.J. SOCS3 binds specific receptor-JAK complexes to control cytokine signaling by direct kinase inhibition. Nat. Struct. Mol. Biol. 2013, 20, 469–476. [Google Scholar] [CrossRef]
- Carow, B.; Rottenberg, M.E. SOCS3, a Major Regulator of Infection and Inflammation. Front. Immunol. 2014, 5, 58. [Google Scholar] [CrossRef]
- Starr, R.; Willson, T.A.; Viney, E.M.; Murray, L.J.; Rayner, J.R.; Jenkins, B.J.; Gonda, T.J.; Alexander, W.S.; Metcalf, D.; Nicola, N.A.; et al. A family of cytokine-inducible inhibitors of signalling. Nature 1997, 387, 917–921. [Google Scholar] [CrossRef]
- Endo, T.A.; Masuhara, M.; Yokouchi, M.; Suzuki, R.; Sakamoto, H.; Mitsui, K.; Matsumoto, A.; Tanimura, S.; Ohtsubo, M.; Misawa, H.; et al. A new protein containing an SH2 domain that inhibits JAK kinases. Nature 1997, 387, 921–924. [Google Scholar] [CrossRef]
- Herrmann, A.; Vogt, M.; Monnigmann, M.; Clahsen, T.; Sommer, U.; Haan, S.; Poli, V.; Heinrich, P.C.; Muller-Newen, G. Nucleocytoplasmic shuttling of persistently activated STAT3. J. Cell Sci. 2007, 120 Pt 18, 3249–3261. [Google Scholar] [CrossRef]
- Ogata, H.; Kobayashi, T.; Chinen, T.; Takaki, H.; Sanada, T.; Minoda, Y.; Koga, K.; Takaesu, G.; Maehara, Y.; Iida, M.; et al. Deletion of the SOCS3 gene in liver parenchymal cells promotes hepatitis-induced hepatocarcinogenesis. Gastroenterology 2006, 131, 179–193. [Google Scholar] [CrossRef]
- Lin, Y.C.; Lin, C.K.; Tsai, Y.H.; Weng, H.H.; Li, Y.C.; You, L.; Chen, J.K.; Jablons, D.M.; Yang, C.T. Adenovirus-mediated SOCS3 gene transfer inhibits the growth and enhances the radiosensitivity of human non-small cell lung cancer cells. Oncol. Rep. 2010, 24, 1605–1612. [Google Scholar]
- Heppler, L.N.; Frank, D.A. Targeting Oncogenic Transcription Factors: Therapeutic Implications of Endogenous STAT Inhibitors. Trends Cancer 2017, 3, 816–827. [Google Scholar]
- Junicho, A.; Matsuda, T.; Yamamoto, T.; Kishi, H.; Korkmaz, K.; Saatcioglu, F.; Fuse, H.; Muraguchi, A. Protein inhibitor of activated STAT3 regulates androgen receptor signaling in prostate carcinoma cells. Biochem. Biophys. Res. Commun. 2000, 278, 9–13. [Google Scholar] [CrossRef]
- Zhang, C.; Mukherjee, S.; Tucker-Burden, C.; Ross, J.L.; Chau, M.J.; Kong, J.; Brat, D.J. TRIM8 regulates stemness in glioblastoma through PIAS3-STAT3. Mol. Oncol. 2017, 11, 280–294. [Google Scholar] [CrossRef]
- Brantley, E.C.; Nabors, L.B.; Gillespie, G.Y.; Choi, Y.H.; Palmer, C.A.; Harrison, K.; Roarty, K.; Benveniste, E.N. Loss of protein inhibitors of activated STAT-3 expression in glioblastoma multiforme tumors: Implications for STAT-3 activation and gene expression. Clin. Cancer Res. 2008, 14, 4694–4704. [Google Scholar] [CrossRef]
- Saydmohammed, M.; Joseph, D.; Syed, V. Curcumin suppresses constitutive activation of STAT-3 by up-regulating protein inhibitor of activated STAT-3 (PIAS-3) in ovarian and endometrial cancer cells. J. Cell Biochem. 2010, 110, 447–456. [Google Scholar] [CrossRef]
- Ogata, Y.; Osaki, T.; Naka, T.; Iwahori, K.; Furukawa, M.; Nagatomo, I.; Kijima, T.; Kumagai, T.; Yoshida, M.; Tachibana, I.; et al. Overexpression of PIAS3 suppresses cell growth and restores the drug sensitivity of human lung cancer cells in association with PI3-K/Akt inactivation. Neoplasia 2006, 8, 817–825. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, C.; Sethi, G.; Ahn, K.S. Brassinin inhibits STAT3 signaling pathway through modulation of PIAS-3 and SOCS-3 expression and sensitizes human lung cancer xenograft in nude mice to paclitaxel. Oncotarget 2015, 6, 6386–6405. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, X.; Guda, K.; Lawrence, E.; Sun, Q.; Watanabe, T.; Iwakura, Y.; Asano, M.; Wei, L.; Yang, Z.; et al. Identification functional characterization of paxillin as a target of protein tyrosine phosphatase receptor, T. Proc. Natl. Acad. Sci. USA 2010, 107, 2592–2597. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, A.; Yu, J.; Possemato, A.; Chen, Y.; Zheng, W.; Polakiewicz, R.D.; Kinzler, K.W.; Vogelstein, B.; Velculescu, V.E.; et al. Identification of STAT3 as a substrate of receptor protein tyrosine phosphatase T. Proc. Natl. Acad. Sci. USA 2007, 104, 4060–4064. [Google Scholar] [CrossRef]
- Chen, Y.W.; Guo, T.; Shen, L.; Wong, K.Y.; Tao, Q.; Choi, W.W.; Au-Yeung, R.K.; Chan, Y.P.; Wong, M.L.; Tang, J.C.; et al. Receptor-type tyrosine-protein phosphatase kappa directly targets STAT3 activation for tumor suppression in nasal NK/T-cell lymphoma. Blood 2015, 125, 1589–1600. [Google Scholar] [CrossRef]
- Mohan, C.D.; Yang, M.H.; Rangappa, S.; Chinnathambi, A.; Alharbi, S.A.; Alahmadi, T.A.; Deivasigamani, A.; Hui, K.M.; Sethi, G.; Rangappa, K.S.; et al. 3-Formylchromone Counteracts STAT3 Signaling Pathway by Elevating SHP-2 Expression in Hepatocellular Carcinoma. Biology 2021, 11, 29. [Google Scholar] [CrossRef]
- Tai, W.T.; Cheng, A.L.; Shiau, C.W.; Liu, C.Y.; Ko, C.H.; Lin, M.W.; Chen, P.J.; Chen, K.F. Dovitinib induces apoptosis and overcomes sorafenib resistance in hepatocellular carcinoma through SHP-1-mediated inhibition of STAT3. Mol. Cancer Ther. 2012, 11, 452–463. [Google Scholar] [CrossRef]
- Schulz-Heddergott, R.; Stark, N.; Edmunds, S.J.; Li, J.; Conradi, L.C.; Bohnenberger, H.; Ceteci, F.; Greten, F.R.; Dobbelstein, M.; Moll, U.M. Therapeutic Ablation of Gain-of-Function Mutant p53 in Colorectal Cancer Inhibits Stat3-Mediated Tumor Growth and Invasion. Cancer Cell 2018, 34, 298–314.e7. [Google Scholar] [CrossRef]
- Su, F.; Ren, F.; Rong, Y.; Wang, Y.; Geng, Y.; Wang, Y.; Feng, M.; Ju, Y.; Li, Y.; Zhao, Z.J.; et al. Protein tyrosine phosphatase Meg2 dephosphorylates signal transducer and activator of transcription 3 and suppresses tumor growth in breast cancer. Breast Cancer Res. 2012, 14, R38. [Google Scholar] [CrossRef]
- Shields, B.J.; Wiede, F.; Gurzov, E.N.; Wee, K.; Hauser, C.; Zhu, H.J.; Molloy, T.J.; O’toole, S.A.; Daly, R.J.; Sutherland, R.L.; et al. TCPTP regulates SFK and STAT3 signaling and is lost in triple-negative breast cancers. Mol. Cell Biol. 2013, 33, 557–570. [Google Scholar] [CrossRef]
- Wani, N.; Nasser, M.W.; Ahirwar, D.K.; Zhao, H.; Miao, Z.; Shilo, K.; Ganju, R.K. C-X-C motif chemokine 12/C-X-C chemokine receptor type 7 signaling regulates breast cancer growth and metastasis by modulating the tumor microenvironment. Breast Cancer Res. 2014, 16, R54. [Google Scholar]
- Zheng, M.; Oh, S.H.; Choi, N.; Choi, Y.J.; Kim, J.; Sung, J.H. CXCL12 inhibits hair growth through CXCR4. Biomed. Pharmacother. 2022, 150, 112996. [Google Scholar] [CrossRef]
- Janssens, R.; Struyf, S.; Proost, P. Pathological roles of the homeostatic chemokine CXCL12. Cytokine Growth Factor Rev. 2018, 44, 51–68. [Google Scholar]
- Wei, C.Y.; Zhu, M.X.; Lu, N.H.; Liu, J.Q.; Yang, Y.W.; Zhang, Y.; Shi, Y.D.; Feng, Z.H.; Li, J.X.; Qi, F.Z.; et al. Circular RNA circ_0020710 drives tumor progression and immune evasion by regulating the miR-370-3p/CXCL12 axis in melanoma. Mol. Cancer 2020, 19, 84. [Google Scholar] [CrossRef]
- Brooks, A.J.; Putoczki, T. JAK-STAT Signalling Pathway in Cancer. Cancers 2020, 12, 1971. [Google Scholar] [CrossRef]
- Giannoni, P.; Scaglione, S.; Quarto, R.; Narcisi, R.; Parodi, M.; Balleari, E.; Barbieri, F.; Pattarozzi, A.; Florio, T.; Ferrini, S.; et al. An interaction between hepatocyte growth factor and its receptor (c-MET) prolongs the survival of chronic lymphocytic leukemic cells through STAT3 phosphorylation: A potential role of mesenchymal cells in the disease. Haematologica 2011, 96, 1015–1023. [Google Scholar] [CrossRef]
- Domanska, U.M.; Kruizinga, R.C.; Nagengast, W.B.; Timmer-Bosscha, H.; Huls, G.; De Vries, E.G.; Walenkamp, A.M. A review on CXCR4/CXCL12 axis in oncology: No place to hide. Eur. J. Cancer 2013, 49, 219–230. [Google Scholar] [CrossRef]
- Shen, H.B.; Gu, Z.Q.; Jian, K.; Qi, J. CXCR4-mediated Stat3 activation is essential for CXCL12-induced cell invasion in bladder cancer. Tumour Biol. 2013, 34, 1839–1845. [Google Scholar] [CrossRef]
- Wu, X.; Tao, P.; Zhou, Q.; Li, J.; Yu, Z.; Wang, X.; Li, J.; Li, C.; Yan, M.; Zhu, Z.; et al. IL-6 secreted by cancer-associated fibroblasts promotes epithelial-mesenchymal transition and metastasis of gastric cancer via JAK2/STAT3 signaling pathway. Oncotarget 2017, 8, 20741–20750. [Google Scholar] [CrossRef]
- Zheng, X.; Carstens, J.L.; Kim, J.; Scheible, M.; Kaye, J.; Sugimoto, H.; Wu, C.C.; Lebleu, V.S.; Kalluri, R. Epithelial-to-mesenchymal transition is dispensable for metastasis but induces chemoresistance in pancreatic cancer. Nature 2015, 527, 525–530. [Google Scholar] [CrossRef]
- Xiang, Z.; Zhou, Z.J.; Xia, G.K.; Zhang, X.H.; Wei, Z.W.; Zhu, J.T.; Yu, J.; Chen, W.; He, Y.; Schwarz, R.E.; et al. A positive crosstalk between CXCR4 and CXCR2 promotes gastric cancer metastasis. Oncogene 2017, 36, 5122–5133. [Google Scholar] [CrossRef]
- Yang, X.Y.; Sheng, Y. miR-101 Represses T-Cell Acute Lymphoblastic Leukemia by Targeting CXCR7/STAT3 Axis. Oncol. Res. 2019, 27, 997–1006. [Google Scholar] [CrossRef]
- Wu, Y.; Tian, L.; Xu, Y.; Zhang, M.; Xiang, S.; Zhao, J.; Wang, Z. CXCR7 silencing inhibits the migration and invasion of human tumor endothelial cells derived from hepatocellular carcinoma by suppressing STAT3. Mol. Med. Rep. 2018, 18, 1644–1650. [Google Scholar] [CrossRef]
- Kim, S.; Kim, J.E.; Kim, N.; Joo, M.; Lee, M.W.; Jeon, H.J.; Ryu, H.; Song, I.C.; Song, G.Y.; Lee, H.J. Decursin inhibits tumor growth, migration, and invasion in gastric cancer by down-regulating CXCR7 expression. Am. J. Cancer Res. 2019, 9, 2007–2018. [Google Scholar]
- Eramo, A.; Lotti, F.; Sette, G.; Pilozzi, E.; Biffoni, M.; Di Virgilio, A.; Conticello, C.; Ruco, L.; Peschle, C.; De Maria, R. Identification and expansion of the tumorigenic lung cancer stem cell population. Cell Death Differ. 2008, 15, 504–514. [Google Scholar] [CrossRef]
- Oh, S.I.; Jeong, H.; Park, H.S.; Choi, K.A.; Hwang, I.; Lee, J.; Cho, J.; Hong, S. Activation of CXCL12-CXCR4 signalling induces conversion of immortalised embryonic kidney cells into cancer stem-like cells. Artif. Cells Nanomed. Biotechnol. 2020, 48, 1303–1313. [Google Scholar] [CrossRef]
- Tang, Y.; Lu, Y.; Chen, Y.; Luo, L.; Cai, L.; Peng, B.; Huang, W.; Liao, H.; Zhao, L.; Pan, M. Pre-metastatic niche triggers SDF-1/CXCR4 axis and promotes organ colonisation by hepatocellular circulating tumour cells via downregulation of Prrx1. J. Exp. Clin. Cancer Res. 2019, 38, 473. [Google Scholar] [CrossRef]
- Lamark, T.; Svenning, S.; Johansen, T. Regulation of selective autophagy: The p62/SQSTM1 paradigm. Essays Biochem. 2017, 61, 609–624. [Google Scholar]
- Zhang, Z.; Yu, Y.; Zhang, Z.; Li, D.; Liang, Z.; Wang, L.; Chen, Y.; Liang, Y.; Niu, H. Cancer-associated fibroblasts-derived CXCL12 enhances immune escape of bladder cancer through inhibiting P62-mediated autophagic degradation of PDL1. J. Exp. Clin. Cancer Res. 2023, 42, 316. [Google Scholar] [CrossRef]
- Feng, M.; Jiang, W.; Kim, B.Y.S.; Zhang, C.C.; Fu, Y.X.; Weissman, I.L. Phagocytosis checkpoints as new targets for cancer immunotherapy. Nat. Rev. Cancer 2019, 19, 568–586. [Google Scholar]
- Topalian, S.L.; Drake, C.G.; Pardoll, D.M. Immune checkpoint blockade: A common denominator approach to cancer therapy. Cancer Cell 2015, 27, 450–461. [Google Scholar] [CrossRef]
- Deng, Y.; Cheng, J.; Fu, B.; Liu, W.; Chen, G.; Zhang, Q.; Yang, Y. Hepatic carcinoma-associated fibroblasts enhance immune suppression by facilitating the generation of myeloid-derived suppressor cells. Oncogene 2017, 36, 1090–1101. [Google Scholar] [CrossRef]
- Pitt, L.A.; Tikhonova, A.N.; Hu, H.; Trimarchi, T.; King, B.; Gong, Y.; Sanchez-Martin, M.; Tsirigos, A.; Littman, D.R.; Ferrando, A.A.; et al. CXCL12-Producing Vascular Endothelial Niches Control Acute T Cell Leukemia Maintenance. Cancer Cell 2015, 27, 755–768. [Google Scholar] [CrossRef]
- Wang, Y.; Miao, H.; Li, W.; Yao, J.; Sun, Y.; Li, Z.; Zhao, L.; Guo, Q. CXCL12/CXCR4 axis confers adriamycin resistance to human chronic myelogenous leukemia and oroxylin A improves the sensitivity of K562/ADM cells. Biochem. Pharmacol. 2014, 90, 212–225. [Google Scholar] [CrossRef]
- Oberlin, E.; Amara, A.; Bachelerie, F.; Bessia, C.; Virelizier, J.L.; Arenzana-Seisdedos, F.; Schwartz, O.; Heard, J.M.; Clark-Lewis, I.; Legler, D.F.; et al. The CXC chemokine SDF-1 is the ligand for LESTR/fusin and prevents infection by T-cell-line-adapted HIV-1. Nature 1996, 382, 833–835. [Google Scholar] [CrossRef]
- Clark, B.R.; Keating, A. Biology of bone marrow stroma. Ann. N. Y. Acad. Sci. 1995, 770, 70–78. [Google Scholar] [CrossRef]
- Madrazo, E.; Gonzalez-Novo, R.; Ortiz-Placin, C.; Garcia De Lacoba, M.; Gonzalez-Murillo, A.; Ramirez, M.; Redondo-Munoz, J. Fast H3K9 methylation promoted by CXCL12 contributes to nuclear changes and invasiveness of T-acute lymphoblastic leukemia cells. Oncogene 2022, 41, 1324–1336. [Google Scholar] [CrossRef]
- Shaim, H.; Estrov, Z.; Harris, D.; Hernandez Sanabria, M.; Liu, Z.; Ruvolo, P.; Thompson, P.A.; Ferrajoli, A.; Daher, M.; Burger, J.; et al. The CXCR4-STAT3-IL-10 Pathway Controls the Immunoregulatory Function of Chronic Lymphocytic Leukemia and Is Modulated by Lenalidomide. Front. Immunol. 2017, 8, 1773. [Google Scholar] [CrossRef]
- Sison, E.A.; Magoon, D.; Li, L.; Annesley, C.E.; Rau, R.E.; Small, D.; Brown, P. Plerixafor as a chemosensitizing agent in pediatric acute lymphoblastic leukemia: Efficacy and potential mechanisms of resistance to CXCR4 inhibition. Oncotarget 2014, 5, 8947–8958. [Google Scholar] [CrossRef]
- Suzuki, Y.; Lutshumba, J.; Chen, K.C.; Abdelaziz, M.H.; Sa, Q.; Ochiai, E. IFN-gamma production by brain-resident cells activates cerebral mRNA expression of a wide spectrum of molecules critical for both innate and T cell-mediated protective immunity to control reactivation of chronic infection with Toxoplasma gondii. Front. Cell Infect. Microbiol. 2023, 13, 1110508. [Google Scholar] [CrossRef]
- Huang, C.; Gu, H.; Yu, Q.; Manukyan, M.C.; Poynter, J.A.; Wang, M. Sca-1+ cardiac stem cells mediate acute cardioprotection via paracrine factor SDF-1 following myocardial ischemia/reperfusion. PLoS ONE 2011, 6, e29246. [Google Scholar] [CrossRef]
- Huang, C.; Gu, H.; Zhang, W.; Manukyan, M.C.; Shou, W.; Wang, M. SDF-1/CXCR4 mediates acute protection of cardiac function through myocardial STAT3 signaling following global ischemia/reperfusion injury. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H1496–H1505. [Google Scholar] [CrossRef]
- Guo, X.Y.; Liu, Q.L.; Liu, W.; Cheng, J.X.; Li, Z.J. Effect and mechanism of miR-135a-5p/CXCL12/JAK-STAT axis on inflammatory response after myocardial infarction. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 12912–12928. [Google Scholar]
- Olive, M.; Mellad, J.A.; Beltran, L.E.; Ma, M.; Cimato, T.; Noguchi, A.C.; San, H.; Childs, R.; Kovacic, J.C.; Boehm, M. p21Cip1 modulates arterial wound repair through the stromal cell-derived factor-1/CXCR4 axis in mice. J. Clin. Investig. 2008, 118, 2050–2061. [Google Scholar] [CrossRef]
- Balaji, S.; Wang, X.; King, A.; Le, L.D.; Bhattacharya, S.S.; Moles, C.M.; Butte, M.J.; De Jesus Perez, V.A.; Liechty, K.W.; Wight, T.N.; et al. Interleukin-10-mediated regenerative postnatal tissue repair is dependent on regulation of hyaluronan metabolism via fibroblast-specific STAT3 signaling. FASEB J. 2017, 31, 868–881. [Google Scholar] [CrossRef]
- Short, W.D.; Steen, E.; Kaul, A.; Wang, X.; Olutoye, O.O., 2nd; Vangapandu, H.V.; Templeman, N.; Blum, A.J.; Moles, C.M.; Narmoneva, D.A.; et al. IL-10 promotes endothelial progenitor cell infiltration and wound healing via STAT3. FASEB J. 2022, 36, e22298. [Google Scholar] [CrossRef]
- Teng, Y.S.; Zhao, Y.L.; Li, M.S.; Liu, Y.G.; Cheng, P.; Lv, Y.P.; Mao, F.Y.; Chen, W.; Yang, S.M.; Hao, C.J.; et al. Upexpression of BHLHE40 in gastric epithelial cells increases CXCL12 production through interaction with p-STAT3 in Helicobacter pylori-associated gastritis. FASEB J. 2020, 34, 1169–1181. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Feng, Q.; Miao, J.; Wu, Q.; Zhou, S.; Shen, W.; Feng, Y.; Hou, F.F.; Liu, Y.; Zhou, L. C-X-C motif chemokine receptor 4 aggravates renal fibrosis through activating JAK/STAT/GSK3beta/beta-catenin pathway. J. Cell Mol. Med. 2020, 24, 3837–3855. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; He, L.; Dong, Y.; Fei, Y.; Wen, J.; Li, X.; Guan, J.; Liu, F.; Zhou, T.; Li, Z.; et al. Sitagliptin ameliorates renal tubular injury in diabetic kidney disease via STAT3-dependent mitochondrial homeostasis through SDF-1alpha/CXCR4 pathway. FASEB J. 2020, 34, 7500–7519. [Google Scholar] [CrossRef] [PubMed]
- Vila-Coro, A.J.; Rodriguez-Frade, J.M.; Martin De Ana, A.; Moreno-Ortiz, M.C.; Martinez, A.C.; Mellado, M. The chemokine SDF-1alpha triggers CXCR4 receptor dimerization and activates the JAK/STAT pathway. FASEB J. 1999, 13, 1699–1710. [Google Scholar] [CrossRef] [PubMed]
- Soriano, S.F.; Hernanz-Falcon, P.; Rodriguez-Frade, J.M.; De Ana, A.M.; Garzon, R.; Carvalho-Pinto, C.; Vila-Coro, A.J.; Zaballos, A.; Balomenos, D.; Martinez, A.C.; et al. Functional inactivation of CXC chemokine receptor 4-mediated responses through SOCS3 up-regulation. J. Exp. Med. 2002, 196, 311–321. [Google Scholar] [CrossRef]
- Ahr, B.; Denizot, M.; Robert-Hebmann, V.; Brelot, A.; Biard-Piechaczyk, M. Identification of the cytoplasmic domains of CXCR4 involved in Jak2 and STAT3 phosphorylation. J. Biol. Chem. 2005, 280, 6692–6700. [Google Scholar] [CrossRef]
- Ho, M.K.; Su, Y.; Yeung, W.W.; Wong, Y.H. Regulation of transcription factors by heterotrimeric G proteins. Curr. Mol. Pharmacol. 2009, 2, 19–31. [Google Scholar] [CrossRef]
- Mellado, M.; Rodriguez-Frade, J.M.; Aragay, A.; Del Real, G.; Martin, A.M.; Vila-Coro, A.J.; Serrano, A.; Mayor, F., Jr.; Martinez, A.C. The chemokine monocyte chemotactic protein 1 triggers Janus kinase 2 activation and tyrosine phosphorylation of the CCR2B receptor. J. Immunol. 1998, 161, 805–813. [Google Scholar] [CrossRef]
- Kim, J.H.; Shim, J.W.; Eum, D.Y.; Kim, S.D.; Choi, S.H.; Yang, K.; Heo, K.; Park, M.T. Downregulation of UHRF1 increases tumor malignancy by activating the CXCR4/AKT-JNK/IL-6/Snail signaling axis in hepatocellular carcinoma cells. Sci. Rep. 2017, 7, 2798. [Google Scholar] [CrossRef]
- Nourbakhsh, M.; Farzaneh, S.; Taghikhani, A.; Zarghi, A.; Noori, S. The Effect of a Newly Synthesized Ferrocene Derivative against MCF-7 Breast Cancer Cells and Spheroid Stem Cells through ROS Production and Inhibition of JAK2/STAT3 Signaling Pathway. Anticancer. Agents Med. Chem. 2020, 20, 875–886. [Google Scholar] [CrossRef]
- Kim, J.H.; Gupta, S.C.; Park, B.; Yadav, V.R.; Aggarwal, B.B. Turmeric (Curcuma longa) inhibits inflammatory nuclear factor (NF)-kappaB and NF-kappaB-regulated gene products and induces death receptors leading to suppressed proliferation, induced chemosensitization, and suppressed osteoclastogenesis. Mol. Nutr. Food Res. 2012, 56, 454–465. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Chen, G.Y.; Song, H.T.; Hong, X.; Yang, Z.Y.; Sui, G.J. Significance of CXCR4, phosphorylated STAT3 and VEGF-A expression in resected non-small cell lung cancer. Exp. Ther. Med. 2011, 2, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.E.; Darnell, J.E., Jr. Stats: Transcriptional control and biological impact. Nat. Rev. Mol. Cell Biol. 2002, 3, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Gargalionis, A.N.; Papavassiliou, K.A.; Papavassiliou, A.G. Targeting STAT3 Signaling Pathway in Colorectal Cancer. Biomedicines 2021, 9, 1016. [Google Scholar] [CrossRef] [PubMed]
- Kako, F.; Gabunia, K.; Ray, M.; Kelemen, S.E.; England, R.N.; Kako, B.; Scalia, R.G.; Autieri, M.V. Interleukin-19 induces angiogenesis in the absence of hypoxia by direct and indirect immune mechanisms. Am. J. Physiol. Cell Physiol. 2016, 310, C931–C941. [Google Scholar] [CrossRef] [PubMed]
- Koscso, B.; Csoka, B.; Kokai, E.; Nemeth, Z.H.; Pacher, P.; Virag, L.; Leibovich, S.J.; Hasko, G. Adenosine augments IL-10-induced STAT3 signaling in M2c macrophages. J. Leukoc. Biol. 2013, 94, 1309–1315. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Tan, Z.; Deng, L.; Chen, Y.; Xia, Y.; Gao, Y.; Wang, X.; Sun, B. Interleukin-22 promotes human hepatocellular carcinoma by activation of STAT3. Hepatology 2011, 54, 900–909. [Google Scholar] [CrossRef] [PubMed]
- Gu, F.M.; Li, Q.L.; Gao, Q.; Jiang, J.H.; Zhu, K.; Huang, X.Y.; Pan, J.F.; Yan, J.; Hu, J.H.; Wang, Z.; et al. IL-17 induces AKT-dependent IL-6/JAK2/STAT3 activation and tumor progression in hepatocellular carcinoma. Mol. Cancer 2011, 10, 150. [Google Scholar] [CrossRef]
- Balabanian, K.; Lagane, B.; Infantino, S.; Chow, K.Y.; Harriague, J.; Moepps, B.; Arenzana-Seisdedos, F.; Thelen, M.; Bachelerie, F. The chemokine SDF-1/CXCL12 binds to and signals through the orphan receptor RDC1 in T lymphocytes. J. Biol. Chem. 2005, 280, 35760–35766. [Google Scholar] [CrossRef]
- Uto-Konomi, A.; Mckibben, B.; Wirtz, J.; Sato, Y.; Takano, A.; Nanki, T.; Suzuki, S. CXCR7 agonists inhibit the function of CXCL12 by down-regulation of CXCR4. Biochem. Biophys. Res. Commun. 2013, 431, 772–776. [Google Scholar] [CrossRef]
- Mu, X.; Shi, W.; Xu, Y.; Xu, C.; Zhao, T.; Geng, B.; Yang, J.; Pan, J.; Hu, S.; Zhang, C.; et al. Tumor-derived lactate induces M2 macrophage polarization via the activation of the ERK/STAT3 signaling pathway in breast cancer. Cell Cycle 2018, 17, 428–438. [Google Scholar] [CrossRef] [PubMed]
- Junior RF, A.; Lira, G.A.; Schomann, T.; Cavalcante, R.S.; Vilar, N.F.; De Paula, R.C.M.; Gomes, R.F.; Chung, C.K.; Jorquera-Cordero, C.; Vepris, O.; et al. Retinoic acid-loaded PLGA nanocarriers targeting cell cholesterol potentialize the antitumour effect of PD-L1 antibody by preventing epithelial-mesenchymal transition mediated by M2-TAM in colorectal cancer. Transl. Oncol. 2023, 31, 101647. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Qiu, L.; Liu, Q.; Ma, Z.; Xie, X.; Luo, Y.; Wu, X. Senescent Fibroblasts Generate a CAF Phenotype through the Stat3 Pathway. Genes 2022, 13, 1579. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yu, B.; Tian, Y.; Dan, J.; Luo, Y.; Wu, X. P53 Mutant p53(N236S) Regulates Cancer-Associated Fibroblasts Properties Through Stat3 Pathway. OncoTargets Ther. 2020, 13, 1355–1363. [Google Scholar] [CrossRef]
- Huang, J.; Huang, L.Q.; He, H.S.; Yan, J.; Huang, C.; Wang, R.; Guan, Y.; Huang, D.P. Overexpression of heme oxygenase-1 in bone marrow stromal cells promotes multiple myeloma resistance through the JAK2/STAT3 pathway. Life Sci. 2020, 257, 118088. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Li, H.; Liu, Z.L.; Li, Q.; Qiu, H.W.; Zeng, L.J.; Yang, W.; Zhang, X.Z.; Li, Z.Y. Activation of STAT3-mediated CXCL12 up-regulation in the dorsal root ganglion contributes to oxaliplatin-induced chronic pain. Mol. Pain 2017, 13, 1744806917747425. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wei, J.; Liu, M.; Wu, S.; Ma, C.; Liu, C.; Huang, K.; Zhang, X.; Guo, R.; Zhang, K.; et al. Epigenetic upregulation of CXCL12 expression contributes to the acquisition and maintenance of morphine-induced conditioned place preference. Exp. Neurol. 2018, 306, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.W.; Yeo, S.Y.; Sung, C.O.; Kim, S.H. Twist1 is a key regulator of cancer-associated fibroblasts. Cancer Res. 2015, 75, 73–85. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Wang, Y.; Xia, R.; Wei, Y.; Wei, X. Role of the CCL2-CCR2 signalling axis in cancer: Mechanisms and therapeutic targeting. Cell Prolif. 2021, 54, e13115. [Google Scholar] [CrossRef]
- Saha, A.; Ahn, S.; Blando, J.; Su, F.; Kolonin, M.G.; Digiovanni, J. Proinflammatory CXCL12-CXCR4/CXCR7 Signaling Axis Drives Myc-Induced Prostate Cancer in Obese Mice. Cancer Res. 2017, 77, 5158–5168. [Google Scholar] [CrossRef]
- Zang, Y.Q.; Feng, Y.Y.; Luo, Y.H.; Zhai, Y.Q.; Ju, X.Y.; Feng, Y.C.; Wang, J.R.; Yu, C.Q.; Jin, C.H. Glycitein induces reactive oxygen species-dependent apoptosis and G0/G1 cell cycle arrest through the MAPK/STAT3/NF-kappaB pathway in human gastric cancer cells. Drug Dev. Res. 2019, 80, 573–584. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, G.Y.; Cao, J.L.; Li, Y.N.; Xue, H.; Wu, H.T.; Jin, C.H. Peimine-induced apoptosis and inhibition of migration by regulating reactive oxygen species-mediated MAPK/STAT3/NF-kappaB and Wnt/beta-catenin signaling pathways in gastric cancer MKN-45 cells. Drug Dev. Res. 2022, 83, 1683–1696. [Google Scholar] [CrossRef]
- Dent, P. HDACIs and the inhibition of invasive potential. Cancer Biol. Ther. 2013, 14, 776–777. [Google Scholar] [CrossRef]
- Han, J.; Feng, Z.; Xie, Y.; Li, F.; Lv, B.; Hua, T.; Zhang, Z.; Sun, C.; Su, D.; Ouyang, Q.; et al. Oncostatin M-induced upregulation of SDF-1 improves Bone marrow stromal cell migration in a rat middle cerebral artery occlusion stroke model. Exp. Neurol. 2019, 313, 49–59. [Google Scholar] [CrossRef]
- Gao, S.; Hu, J.; Wu, X.; Liang, Z. PMA treated THP-1-derived-IL-6 promotes EMT of SW48 through STAT3/ERK-dependent activation of Wnt/beta-catenin signaling pathway. Biomed. Pharmacother. 2018, 108, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Shabbir, A.; Cox, A.; Rodriguez-Menocal, L.; Salgado, M.; Van Badiavas, E. Mesenchymal Stem Cell Exosomes Induce Proliferation and Migration of Normal and Chronic Wound Fibroblasts, and Enhance Angiogenesis In Vitro. Stem Cells Dev. 2015, 24, 1635–1647. [Google Scholar] [CrossRef]
- Li, J.; Luco, A.L.; Camirand, A.; St-Arnaud, R.; Kremer, R. Vitamin D Regulates CXCL12/CXCR4 and Epithelial-to-Mesenchymal Transition in a Model of Breast Cancer Metastasis to Lung. Endocrinology 2021, 162, bqab049. [Google Scholar] [CrossRef]
- Balic, A.; Sorensen, M.D.; Trabulo, S.M.; Sainz, B., Jr.; Cioffi, M.; Vieira, C.R.; Miranda-Lorenzo, I.; Hidalgo, M.; Kleeff, J.; Erkan, M.; et al. Chloroquine targets pancreatic cancer stem cells via inhibition of CXCR4 and hedgehog signaling. Mol. Cancer Ther. 2014, 13, 1758–1771. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhang, G.; Liu, N.; Lin, W.; Ji, S.; Zheng, M.; Chen, K.; Liang, X.; Li, G.; Ma, Y.; et al. A novel orally available Syk/Src/Jak2 inhibitor, SKLB-850, showed potent anti-tumor activities in B cell lymphoma (BCL) models. Oncotarget 2017, 8, 111495–111507. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Song, Z.; Guo, Q.; Li, J. Synergistic Effect and Molecular Mechanisms of Traditional Chinese Medicine on Regulating Tumor Microenvironment and Cancer Cells. Biomed. Res. Int. 2016, 2016, 1490738. [Google Scholar] [CrossRef]
- Wu, J.; Xu, J.; Eksioglu, E.A.; Chen, X.; Zhou, J.; Fortenbery, N.; Wei, S.; Dong, J. Icariside II induces apoptosis of melanoma cells through the downregulation of survival pathways. Nutr. Cancer 2013, 65, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Cui, W.; Zhao, Z.; Gong, W.; Wei, Y.; Liu, J.; Li, M.; Li, Q.; Yan, C.; Qiu, J.; et al. Targeting Tumor Microenvironment: Effects of Chinese Herbal Formulae on Macrophage-Mediated Lung Cancer in Mice. Evid. Based Complement. Altern. Med. 2017, 2017, 7187168. [Google Scholar] [CrossRef] [PubMed]
- Badr, G.; Mohany, M.; Abu-Tarboush, F. Thymoquinone decreases F-actin polymerization and the proliferation of human multiple myeloma cells by suppressing STAT3 phosphorylation and Bcl2/Bcl-XL expression. Lipids Health Dis. 2011, 10, 236. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Min, J.W.; Kong, W.L.; He, X.H.; Li, J.X.; Peng, B.W. A review on the pharmacological effects of vitexin and isovitexin. Fitoterapia 2016, 115, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Mohan, C.D.; Shanmugam, M.K.; Rangappa, S.; Sethi, G.; Siveen, K.S.; Chinnathambi, A.; Alahmadi, T.A.; Alharbi, S.A.; Basappa, S.; et al. Vitexin abrogates invasion and survival of hepatocellular carcinoma cells through targeting STAT3 signaling pathway. Biochimie 2020, 175, 58–68. [Google Scholar] [CrossRef]
- Yamasaki, H. Pharmacology of sinomenine, an anti-rheumatic alkaloid from Sinomenium acutum. Acta Med. Okayama 1976, 30, 1–20. [Google Scholar] [PubMed]
- Zhou, L.; Luan, H.; Liu, Q.; Jiang, T.; Liang, H.; Dong, X.; Shang, H. Activation of PI3K/Akt and ERK signaling pathways antagonized sinomenine-induced lung cancer cell apoptosis. Mol. Med. Rep. 2012, 5, 1256–1260. [Google Scholar] [PubMed]
- Li, X.J.; Yue, P.Y.; Ha, W.Y.; Wong, D.Y.; Tin, M.M.; Wang, P.X.; Wong, R.N.; Liu, L. Effect of sinomenine on gene expression of the IL-1 beta-activated human synovial sarcoma. Life Sci. 2006, 79, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.L.; Zeng, J.; Chen, Y.L.; He, P.M.; Wen, M.X.; Ren, M.D.; Hu, Y.N.; Lu, G.F.; He, S. Sinomenine hydrochloride inhibits human hepatocellular carcinoma cell growth in vitro and in vivo: Involvement of cell cycle arrest and apoptosis induction. Int. J. Oncol. 2013, 42, 229–238. [Google Scholar] [CrossRef]
- Lv, Y.; Li, C.; Li, S.; Hao, Z. Sinomenine inhibits proliferation of SGC-7901 gastric adenocarcinoma cells via suppression of cyclooxygenase-2 expression. Oncol. Lett. 2011, 2, 741–745. [Google Scholar] [CrossRef]
- Xie, T.; Ren, H.Y.; Lin, H.Q.; Mao, J.P.; Zhu, T.; Wang, S.D.; Ye, Z.M. Sinomenine prevents metastasis of human osteosarcoma cells via S phase arrest and suppression of tumor-related neovascularization and osteolysis through the CXCR4-STAT3 pathway. Int. J. Oncol. 2016, 48, 2098–2112. [Google Scholar] [CrossRef]
- Garg, R.; Ingle, A.; Maru, G. Dietary turmeric modulates DMBA-induced p21ras, MAP kinases and AP-1/NF-kappaB pathway to alter cellular responses during hamster buccal pouch carcinogenesis. Toxicol. Appl. Pharmacol. 2008, 232, 428–439. [Google Scholar] [CrossRef]
- Sun, R.; Sun, Y.; Wu, C.; Liu, Y.; Zhou, M.; Dong, Y.; Du, G.; Luo, H.; Shi, B.; Jiang, H.; et al. CXCR4-modified CAR-T cells suppresses MDSCs recruitment via STAT3/NF-kappaB/SDF-1alpha axis to enhance efficacy against pancreatic cancer. Mol. Ther. 2023, 31, 3193–3209. [Google Scholar] [CrossRef]
- Lapillonne, H.; Konopleva, M.; Tsao, T.; Gold, D.; Mcqueen, T.; Sutherland, R.L.; Madden, T.; Andreeff, M. Activation of peroxisome proliferator-activated receptor gamma by a novel synthetic triterpenoid 2-cyano-3,12-dioxooleana-1,9-dien-28-oic acid induces growth arrest and apoptosis in breast cancer cells. Cancer Res. 2003, 63, 5926–5939. [Google Scholar]
- Konopleva, M.; Zhang, W.; Shi, Y.X.; Mcqueen, T.; Tsao, T.; Abdelrahim, M.; Munsell, M.F.; Johansen, M.; Yu, D.; Madden, T.; et al. Synthetic triterpenoid 2-cyano-3,12-dioxooleana-1,9-dien-28-oic acid induces growth arrest in HER2-overexpressing breast cancer cells. Mol. Cancer Ther. 2006, 5, 317–328. [Google Scholar] [CrossRef]
- Liby, K.; Risingsong, R.; Royce, D.B.; Williams, C.R.; Yore, M.M.; Honda, T.; Gribble, G.W.; Lamph, W.W.; Vannini, N.; Sogno, I.; et al. Prevention and treatment of experimental estrogen receptor-negative mammary carcinogenesis by the synthetic triterpenoid CDDO-methyl Ester and the rexinoid LG100268. Clin. Cancer Res. 2008, 14, 4556–4563. [Google Scholar] [CrossRef]
- Tran, K.; Risingsong, R.; Royce, D.; Williams, C.R.; Sporn, M.B.; Liby, K. The synthetic triterpenoid CDDO-methyl ester delays estrogen receptor-negative mammary carcinogenesis in polyoma middle T mice. Cancer Prev. Res. 2012, 5, 726–734. [Google Scholar] [CrossRef]
- Melo RC, C.; Longhini, A.L.; Bigarella, C.L.; Baratti, M.O.; Traina, F.; Favaro, P.; De Melo Campos, P.; Saad, S.T. CXCR7 is highly expressed in acute lymphoblastic leukemia and potentiates CXCR4 response to CXCL12. PLoS ONE 2014, 9, e85926. [Google Scholar] [CrossRef]
- Burger, M.; Hartmann, T.; Krome, M.; Rawluk, J.; Tamamura, H.; Fujii, N.; Kipps, T.J.; Burger, J.A. Small peptide inhibitors of the CXCR4 chemokine receptor (CD184) antagonize the activation, migration, and antiapoptotic responses of CXCL12 in chronic lymphocytic leukemia B cells. Blood 2005, 106, 1824–1830. [Google Scholar] [CrossRef]
- Dipersio, J.F.; Micallef, I.N.; Stiff, P.J.; Bolwell, B.J.; Maziarz, R.T.; Jacobsen, E.; Nademanee, A.; Mccarty, J.; Bridger, G.; Calandra, G.; et al. Phase III prospective randomized double-blind placebo-controlled trial of plerixafor plus granulocyte colony-stimulating factor compared with placebo plus granulocyte colony-stimulating factor for autologous stem-cell mobilization and transplantation for patients with non-Hodgkin’s lymphoma. J. Clin. Oncol. 2009, 27, 4767–4773. [Google Scholar]
- Hoy, S.M. Motixafortide: First Approval. Drugs 2023, 83, 1635–1643. [Google Scholar] [CrossRef] [PubMed]
- Mullard, A. CXCR4 chemokine antagonist scores a first FDA approval for WHIM syndrome. Nat. Rev. Drug Discov. 2024, 23, 411. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Fang, X.; Zhang, H.; Wang, X.; Li, M.; Jiang, W.; Tian, F.; Zhu, L.; Bian, Z. AMD3100 combined with triptolide inhibit proliferation, invasion and metastasis and induce apoptosis of human U2OS osteosarcoma cells. Biomed. Pharmacother. 2017, 86, 677–685. [Google Scholar] [CrossRef]
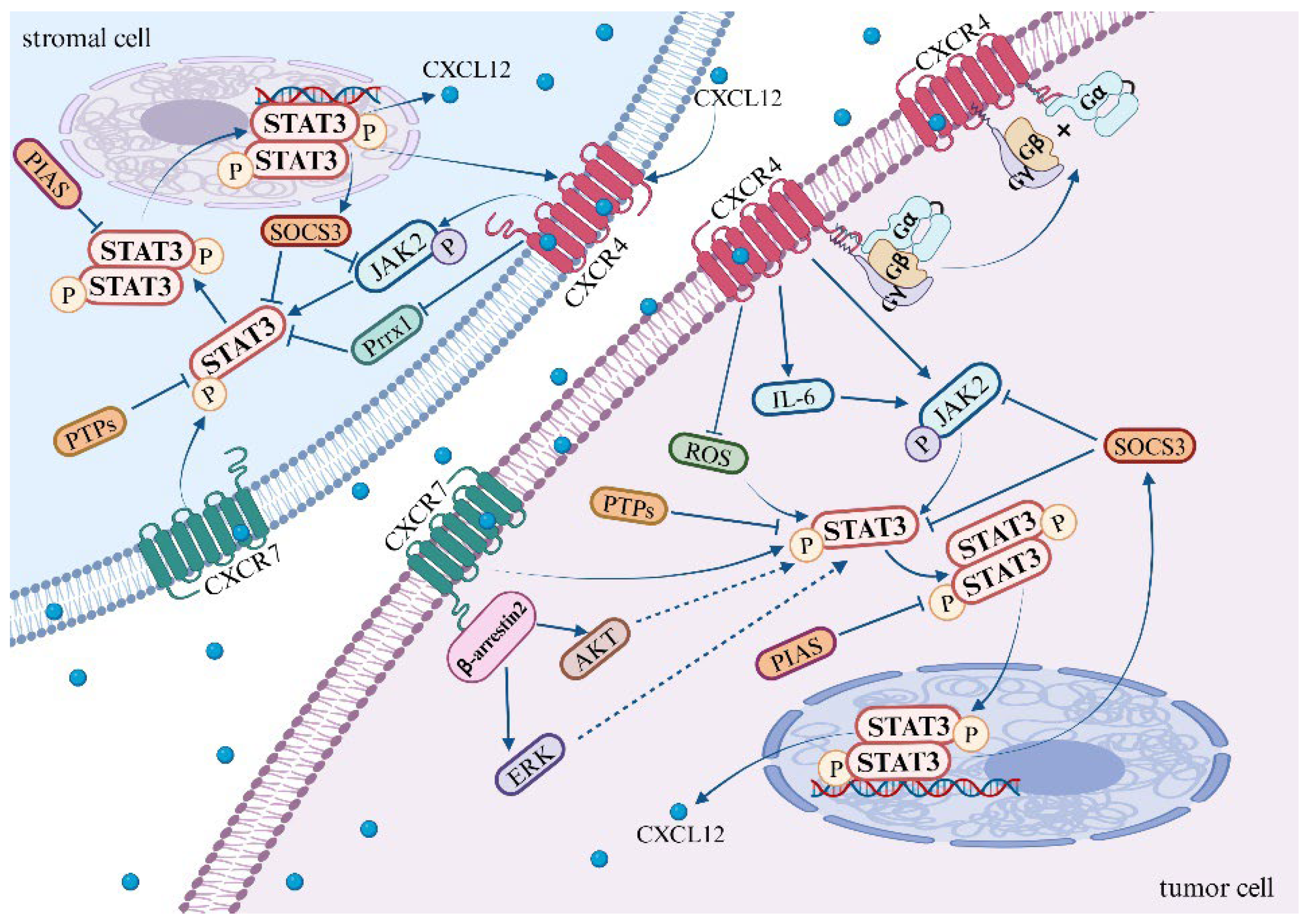
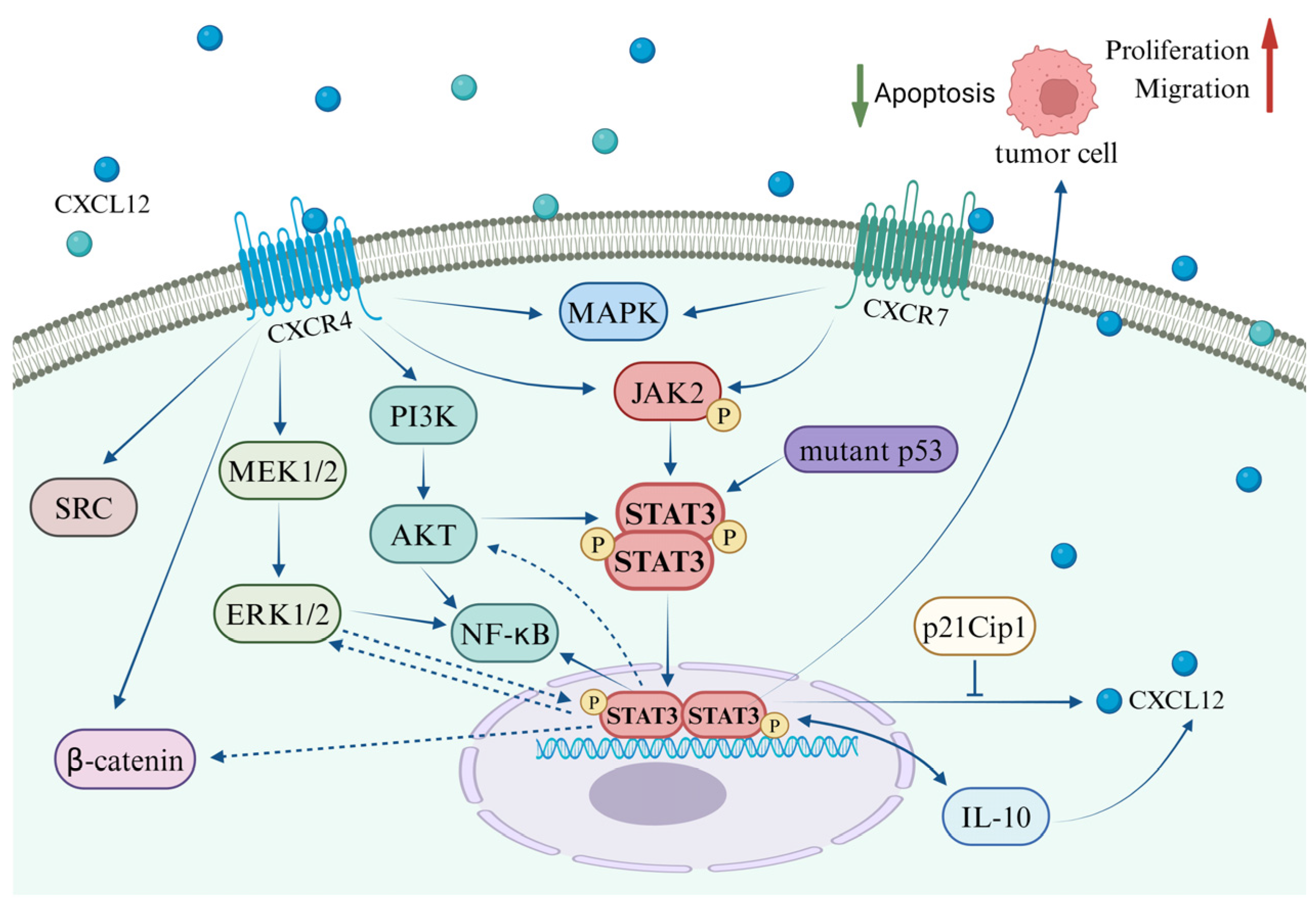
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).