
Dysfunction of Mitochondrial Dynamics Induces Endocytosis Defect and Cell Damage in Drosophila Nephrocytes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Drosophila Lines
2.2. Mitochondrial Membrane Potential Measurement in Drosophila Nephrocytes
2.3. Transmission Electron Microscopy (TEM) for Drosophila Nephrocytes
2.4. Measurement of Mitochondrial Dimensions
2.5. Assessment of Dextran and Albumin Uptake in Drosophila Nephrocytes
2.6. Fluorescent Immunochemistry
2.7. Reactive Oxygen Species Assay in Drosophila Nephrocytes
2.8. Fluorescence Confocal Microscopy of Drosophila Nephrocytes
2.9. Statistical Analysis of Drosophila Assays
3. Results
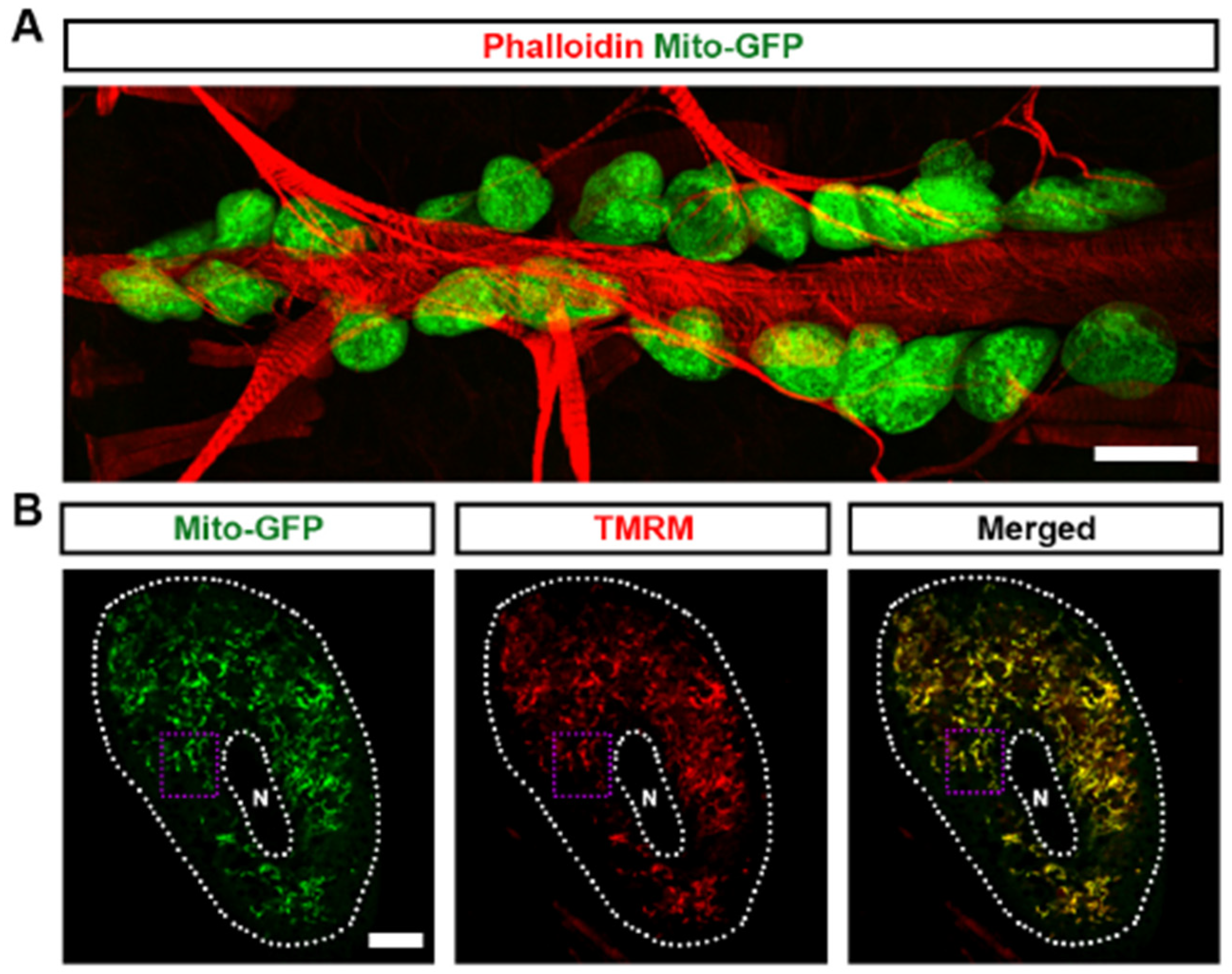
3.1. Mitochondria Highly Existed in Drosophila Nephrocytes
3.2. Pink1–Park-Mediated Defective Mitochondrial Dynamics Lead to Reduced Nephrocyte Function and a Disrupted Slit Diaphragm Filtration Structure
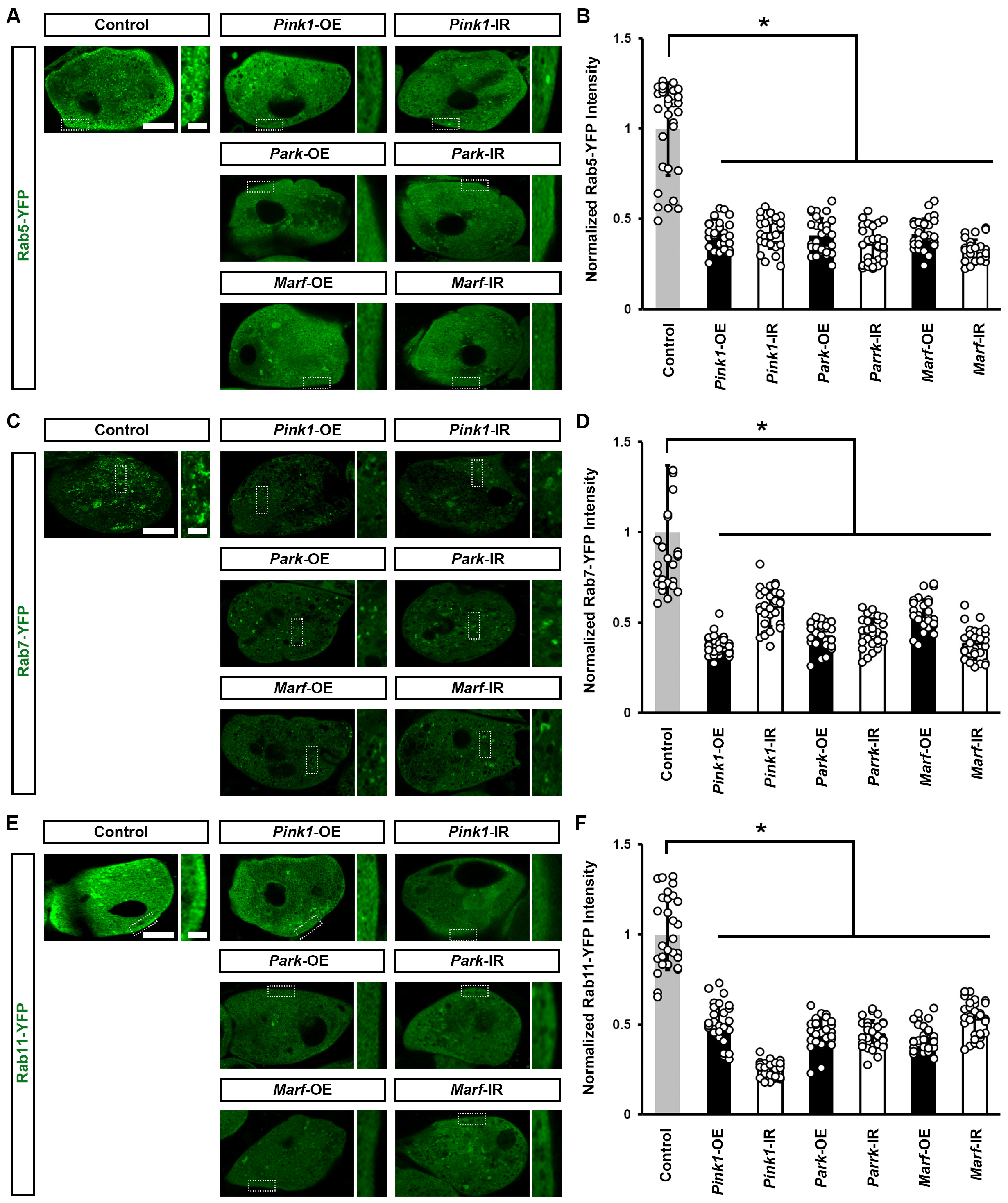
3.3. Pink1–Park-Mediated Defective Mitochondrial Dynamics Lead to Impaired Endocytic Membrane Trafficking
3.4. Pink1–Park-Mediated Defective Mitochondrial Dynamics Lead to Induced Cellular Protein Aggregation and Increased ROS
3.5. Defective Endocytosis Leads to Reduced Nephrocyte Function, Induced Cellular Protein Aggregation, and Increased ROS
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kühlbrandt, W. Structure and Function of Mitochondrial Membrane Protein Complexes. BMC Biol. 2015, 13, 89. [Google Scholar] [CrossRef] [PubMed]
- Al Ojaimi, M.; Salah, A.; El-Hattab, A.W. Mitochondrial Fission and Fusion: Molecular Mechanisms, Biological Functions, and Related Disorders. Membranes 2022, 12, 893. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Detmer, S.A.; Ewald, A.J.; Griffin, E.E.; Fraser, S.E.; Chan, D.C. Mitofusins Mfn1 and Mfn2 Coordinately Regulate Mitochondrial Fusion and Are Essential for Embryonic Development. J. Cell Biol. 2003, 160, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Dodson, M.W.; Huang, H.; Guo, M. The Parkinson’s Disease Genes Pink1 and Parkin Promote Mitochondrial Fission and/or Inhibit Fusion in Drosophila. Proc. Natl. Acad. Sci. USA 2008, 105, 14503–14508. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Sibilla, C.; Liu, R.; Yun, J.; Hay, B.A.; Blackstone, C.; Chan, D.C.; Harvey, R.J.; Guo, M. Clueless/CLUH Regulates Mitochondrial Fission by Promoting Recruitment of Drp1 to Mitochondria. Nat. Commun. 2022, 13, 1582. [Google Scholar] [CrossRef] [PubMed]
- Ge, P.; Dawson, V.L.; Dawson, T.M. PINK1 and Parkin Mitochondrial Quality Control: A Source of Regional Vulnerability in Parkinson’s Disease. Mol. Neurodegener. 2020, 15, 20. [Google Scholar] [CrossRef] [PubMed]
- McInnes, J. Insights on Altered Mitochondrial Function and Dynamics in the Pathogenesis of Neurodegeneration. Transl. Neurodegener. 2013, 2, 12. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Ying, J.; Wang, X.; Zhao, T.; Yoon, S.; Fang, Y.; Zheng, Q.; Liu, X.; Yu, W.; Hua, F. Mitochondrial Dynamics: A Key Role in Neurodegeneration and a Potential Target for Neurodegenerative Disease. Front. Neurosci. 2021, 15, 654785. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.; Mukherjee, A.; Nagotu, S. Mitochondrial Dynamics and Its Impact on Human Health and Diseases: Inside the DRP1 Blackbox. J. Mol. Med. 2022, 100, 1–21. [Google Scholar] [CrossRef]
- Ong, S.-B.; Hall, A.R.; Hausenloy, D.J. Mitochondrial Dynamics in Cardiovascular Health and Disease. Antioxid. Redox Signal. 2013, 19, 400–414. [Google Scholar] [CrossRef] [PubMed]
- Yapa, N.M.B.; Lisnyak, V.; Reljic, B.; Ryan, M.T. Mitochondrial Dynamics in Health and Disease. FEBS Lett. 2021, 595, 1184–1204. [Google Scholar] [CrossRef] [PubMed]
- Pacheu-Grau, D.; Rucktäschel, R.; Deckers, M. Mitochondrial Dysfunction and Its Role in Tissue-Specific Cellular Stress. Cell Stress 2018, 2, 184–199. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, N.; Nomura, M.; Jofuku, A.; Kato, H.; Suzuki, S.O.; Masuda, K.; Otera, H.; Nakanishi, Y.; Nonaka, I.; Goto, Y.-I.; et al. Mitochondrial Fission Factor Drp1 Is Essential for Embryonic Development and Synapse Formation in Mice. Nat. Cell Biol. 2009, 11, 958–966. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, Y.; Hoshijima, M.; Seo, K.; Bedja, D.; Sysa-Shah, P.; Andrabi, S.A.; Chen, W.; Höke, A.; Dawson, V.L.; Dawson, T.M.; et al. Parkin-Independent Mitophagy Requires Drp1 and Maintains the Integrity of Mammalian Heart and Brain. EMBO J. 2014, 33, 2798–2813. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, P.; Schnellmann, R.G. Mitochondrial Energetics in the Kidney. Nat. Rev. Nephrol. 2017, 13, 629–646. [Google Scholar] [CrossRef] [PubMed]
- Chaiyarit, S.; Thongboonkerd, V. Mitochondrial Dysfunction and Kidney Stone Disease. Front. Physiol. 2020, 11, 566506. [Google Scholar] [CrossRef] [PubMed]
- Gujarati, N.A.; Vasquez, J.M.; Bogenhagen, D.F.; Mallipattu, S.K. The Complicated Role of Mitochondria in the Podocyte. Am. J. Physiol. Renal Physiol. 2020, 319, F955–F965. [Google Scholar] [CrossRef] [PubMed]
- Reiser, J.; Altintas, M.M. Podocytes. F1000Research 2016, 5, 114. [Google Scholar] [CrossRef] [PubMed]
- Weavers, H.; Prieto-Sánchez, S.; Grawe, F.; Garcia-López, A.; Artero, R.; Wilsch-Bräuninger, M.; Ruiz-Gómez, M.; Skaer, H.; Denholm, B. The Insect Nephrocyte Is a Podocyte-like Cell with a Filtration Slit Diaphragm. Nature 2009, 457, 322–326. [Google Scholar] [CrossRef]
- Fu, Y.; Zhu, J.-Y.; Richman, A.; Zhao, Z.; Zhang, F.; Ray, P.E.; Han, Z. A Drosophila Model System to Assess the Function of Human Monogenic Podocyte Mutations That Cause Nephrotic Syndrome. Hum. Mol. Genet. 2017, 26, 768–780. [Google Scholar] [CrossRef] [PubMed]
- Simons, M.; Huber, T.B. Flying Podocytes. Kidney Int. 2009, 75, 455–457. [Google Scholar] [CrossRef] [PubMed]
- Na, J.; Cagan, R. The Drosophila Nephrocyte: Back on Stage. J. Am. Soc. Nephrol. 2013, 24, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-Y.; Fu, Y.; Richman, A.; Zhao, Z.; Ray, P.E.; Han, Z. A Personalized Model of COQ2 Nephropathy Rescued by the Wild-Type COQ2 Allele or Dietary Coenzyme Q10 Supplementation. J. Am. Soc. Nephrol. 2017, 28, 2607–2617. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-Y.; van de Leemput, J.; Han, Z. Promoting Mitochondrial Dynamics by Inhibiting the PINK1/PRKN Pathway to Relieve Diabetic Nephropathy. Dis. Model. Mech. 2024; ahead of print. [Google Scholar] [CrossRef]
- Kim, K.; Cha, S.J.; Choi, H.-J.; Kang, J.S.; Lee, E.Y. Dysfunction of Mitochondrial Dynamics in Drosophila Model of Diabetic Nephropathy. Life 2021, 11, 67. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wen, P.; van de Leemput, J.; Zhao, Z.; Han, Z. Slit Diaphragm Maintenance Requires Dynamic Clathrin-Mediated Endocytosis Facilitated by AP-2, Lap, Aux and Hsc70-4 in Nephrocytes. Cell Biosci. 2021, 11, 83. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Frederick, R.L.; Shaw, J.M. Moving Mitochondria: Establishing Distribution of an Essential Organelle. Traffic 2007, 8, 1668–1675. [Google Scholar] [CrossRef] [PubMed]
- Giacomello, M.; Pyakurel, A.; Glytsou, C.; Scorrano, L. The Cell Biology of Mitochondrial Membrane Dynamics. Nat. Rev. Mol. Cell Biol. 2020, 21, 204–224. [Google Scholar] [CrossRef] [PubMed]
- de Boer, L.; de Vries, M.C.; Smeitink, J.A.M.; Koopman, W.J.H. Disorders of Mitochondrial Homeostasis, Dynamics, Protein Import, and Quality Control. In Physician’s Guide to the Diagnosis, Treatment, and Follow-up of Inherited Metabolic Diseases; Springer International Publishing: Cham, Switzerland, 2022; pp. 889–913. ISBN 9783030677268. [Google Scholar]
- Zorova, L.D.; Popkov, V.A.; Plotnikov, E.Y.; Silachev, D.N.; Pevzner, I.B.; Jankauskas, S.S.; Babenko, V.A.; Zorov, S.D.; Balakireva, A.V.; Juhaszova, M.; et al. Mitochondrial Membrane Potential. Anal. Biochem. 2018, 552, 50–59. [Google Scholar] [CrossRef] [PubMed]
- van de Leemput, J.; Wen, P.; Han, Z. Using Drosophila Nephrocytes to Understand the Formation and Maintenance of the Podocyte Slit Diaphragm. Front. Cell Dev. Biol. 2022, 10, 837828. [Google Scholar] [CrossRef] [PubMed]
- Wen, P.; Zhang, F.; Fu, Y.; Zhu, J.-Y.; Han, Z. Exocyst Genes Are Essential for Recycling Membrane Proteins and Maintaining Slit Diaphragm in Drosophila Nephrocytes. J. Am. Soc. Nephrol. 2020, 31, 1024–1034. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhu, J.-Y.; Zhang, F.; Richman, A.; Zhao, Z.; Han, Z. Comprehensive Functional Analysis of Rab GTPases in Drosophila Nephrocytes. Cell Tissue Res. 2017, 368, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Sun, L.; Chen, X.; Zhang, D. Oxidative Stress, Mitochondrial Damage and Neurodegenerative Diseases. Neural Regen. Res. 2013, 8, 2003–2014. [Google Scholar] [PubMed]
- McCormick, R.; Pearson, T.; Vasilaki, A. Manipulation of Environmental Oxygen Modifies Reactive Oxygen and Nitrogen Species Generation during Myogenesis. Redox Biol. 2016, 8, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Birgisdottir, Å.B.; Johansen, T. Autophagy and Endocytosis—Interconnections and Interdependencies. J. Cell Sci. 2020, 133, jcs228114. [Google Scholar] [CrossRef] [PubMed]
- Tooze, S.A.; Abada, A.; Elazar, Z. Endocytosis and Autophagy: Exploitation or Cooperation? Cold Spring Harb. Perspect. Biol. 2014, 6, a018358. [Google Scholar] [CrossRef] [PubMed]
- Lévy, E.; El Banna, N.; Baïlle, D.; Heneman-Masurel, A.; Truchet, S.; Rezaei, H.; Huang, M.-E.; Béringue, V.; Martin, D.; Vernis, L. Causative Links between Protein Aggregation and Oxidative Stress: A Review. Int. J. Mol. Sci. 2019, 20, 3896. [Google Scholar] [CrossRef] [PubMed]
- Weng, M.; Xie, X.; Liu, C.; Lim, K.-L.; Zhang, C.-W.; Li, L. The Sources of Reactive Oxygen Species and Its Possible Role in the Pathogenesis of Parkinson’s Disease. Park. Dis. 2018, 2018, 9163040. [Google Scholar] [CrossRef] [PubMed]
- Passarella, S.; Schurr, A.; Portincasa, P. Mitochondrial Transport in Glycolysis and Gluconeogenesis: Achievements and Perspectives. Int. J. Mol. Sci. 2021, 22, 12620. [Google Scholar] [CrossRef] [PubMed]
- Haythorne, E.; Lloyd, M.; Walsby-Tickle, J.; Tarasov, A.I.; Sandbrink, J.; Portillo, I.; Exposito, R.T.; Sachse, G.; Cyranka, M.; Rohm, M.; et al. Altered Glycolysis Triggers Impaired Mitochondrial Metabolism and MTORC1 Activation in Diabetic β-Cells. Nat. Commun. 2022, 13, 6754. [Google Scholar] [CrossRef] [PubMed]
- Koehler, S.; Huber, T.B. Insights into Human Kidney Function from the Study of Drosophila. Pediatr. Nephrol. 2023, 38, 3875–3887. [Google Scholar] [CrossRef] [PubMed]
- Dow, J.A.T.; Simons, M.; Romero, M.F. Drosophila Melanogaster: A Simple Genetic Model of Kidney Structure, Function and Disease. Nat. Rev. Nephrol. 2022, 18, 417–434. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Chen, X. The Drosophila Nephrocyte Has a Glomerular Filtration System. Nat. Rev. Nephrol. 2014, 10, 491. [Google Scholar] [CrossRef] [PubMed]
- Rani, L.; Gautam, N.K. Drosophila Renal Organ as a Model for Identification of Targets and Screening of Potential Therapeutic Agents for Diabetic Nephropathy. Curr. Drug Targets 2018, 19, 1980–1990. [Google Scholar] [CrossRef] [PubMed]
- Abtahi, S.L.; Masoudi, R.; Haddadi, M. The Distinctive Role of Tau and Amyloid Beta in Mitochondrial Dysfunction through Alteration in Mfn2 and Drp1 MRNA Levels: A Comparative Study in Drosophila Melanogaster. Gene 2020, 754, 144854. [Google Scholar] [CrossRef] [PubMed]
- Gegg, M.E.; Mark Cooper, J.; Chau, K.-Y.; Rojo, M.; Schapira, A.H.V.; Taanman, J.-W. Mitofusin 1 and Mitofusin 2 Are Ubiquitinated in a PINK1/Parkin-Dependent Manner upon Induction of Mitophagy. Hum. Mol. Genet. 2013, 22, 1697. [Google Scholar] [CrossRef]
- Guo, M. Drosophila as a Model to Study Mitochondrial Dysfunction in Parkinson’s Disease. Cold Spring Harb. Perspect. Med. 2012, 2, a009944. [Google Scholar] [PubMed]
- Hermle, T.; Braun, D.A.; Helmstädter, M.; Huber, T.B.; Hildebrandt, F. Modeling Monogenic Human Nephrotic Syndrome in the Drosophila Garland Cell Nephrocyte. J. Am. Soc. Nephrol. 2017, 28, 1521–1533. [Google Scholar] [PubMed]
- Zhang, P.N.; Zhou, M.Q.; Guo, J.; Zheng, H.J.; Tang, J.; Zhang, C.; Liu, Y.N.; Liu, W.J.; Wang, Y.X. Mitochondrial Dysfunction and Diabetic Nephropathy: Nontraditional Therapeutic Opportunities. J. Diabetes Res. 2021, 2021, 1010268. [Google Scholar]
- Liu, S.; Yuan, Y.; Xue, Y.; Xing, C.; Zhang, B. Podocyte Injury in Diabetic Kidney Disease: A Focus on Mitochondrial Dysfunction. Front. Cell Dev. Biol. 2022, 10, 832887. [Google Scholar] [CrossRef] [PubMed]
- Na, J.; Sweetwyne, M.T.; Park, A.S.D.; Susztak, K.; Cagan, R.L. Diet-Induced Podocyte Dysfunction in Drosophila and Mammals. Cell Rep. 2015, 12, 636–647. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, J.-y.; Duan, J.; van de Leemput, J.; Han, Z. Dysfunction of Mitochondrial Dynamics Induces Endocytosis Defect and Cell Damage in Drosophila Nephrocytes. Cells 2024, 13, 1253. https://doi.org/10.3390/cells13151253
Zhu J-y, Duan J, van de Leemput J, Han Z. Dysfunction of Mitochondrial Dynamics Induces Endocytosis Defect and Cell Damage in Drosophila Nephrocytes. Cells. 2024; 13(15):1253. https://doi.org/10.3390/cells13151253
Chicago/Turabian StyleZhu, Jun-yi, Jianli Duan, Joyce van de Leemput, and Zhe Han. 2024. "Dysfunction of Mitochondrial Dynamics Induces Endocytosis Defect and Cell Damage in Drosophila Nephrocytes" Cells 13, no. 15: 1253. https://doi.org/10.3390/cells13151253
APA StyleZhu, J.-y., Duan, J., van de Leemput, J., & Han, Z. (2024). Dysfunction of Mitochondrial Dynamics Induces Endocytosis Defect and Cell Damage in Drosophila Nephrocytes. Cells, 13(15), 1253. https://doi.org/10.3390/cells13151253