
Imbalance of B-Cell Subpopulations in the Microenvironment of Sarcoidosis or Lung Cancer


 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Materials: Lymph Node Aspirates and Peripheral Blood
2.3. Methods
2.3.1. Hematological Parameters
2.3.2. Flow Cytometry Analysis
2.3.3. Flow Cytometry Analysis
3. Results
3.1. Leukocyte Subpopulations in Lymph Node Aspirates
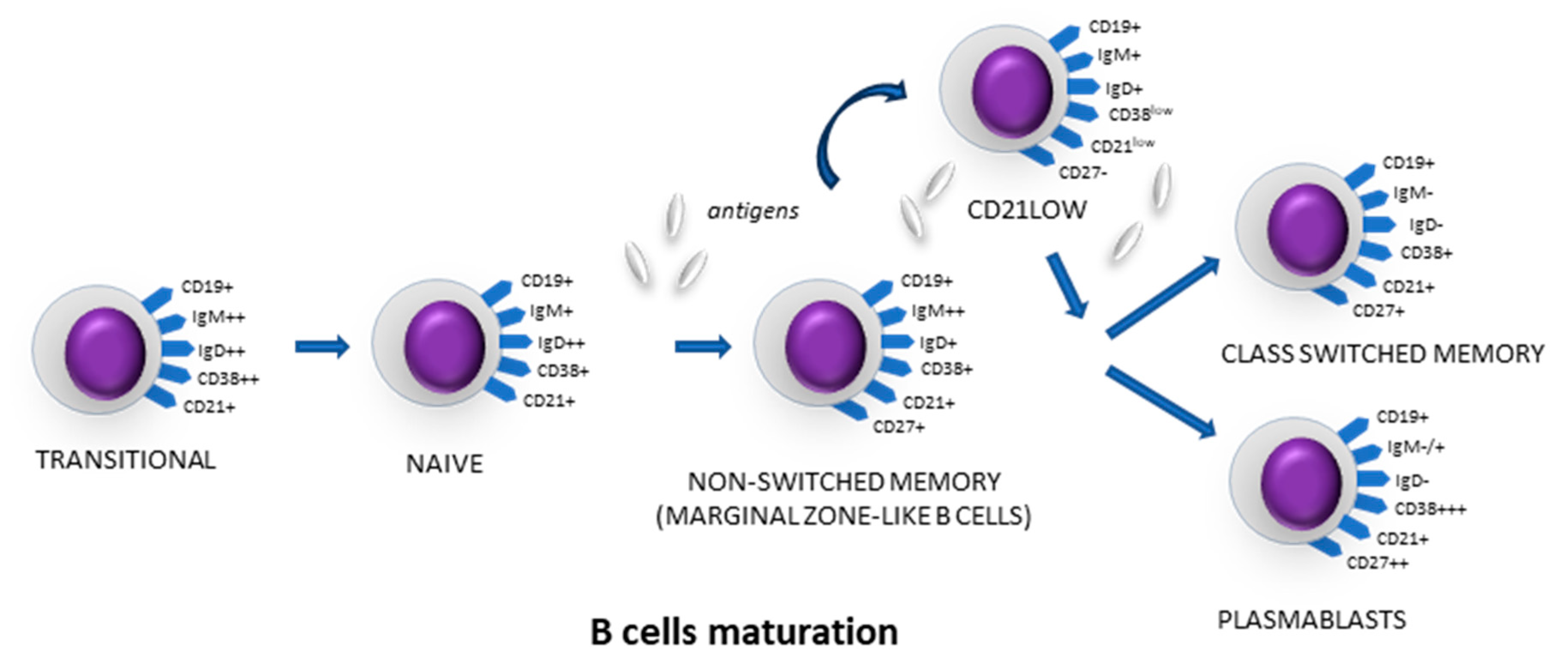
3.2. B-Cell Maturation Subsets in Lymph Node Aspirates
3.3. New Lymphocytes and Neutrophils Parameters in Lymph Node Aspirates
3.4. Leukocyte Subpopulations in Peripheral Blood
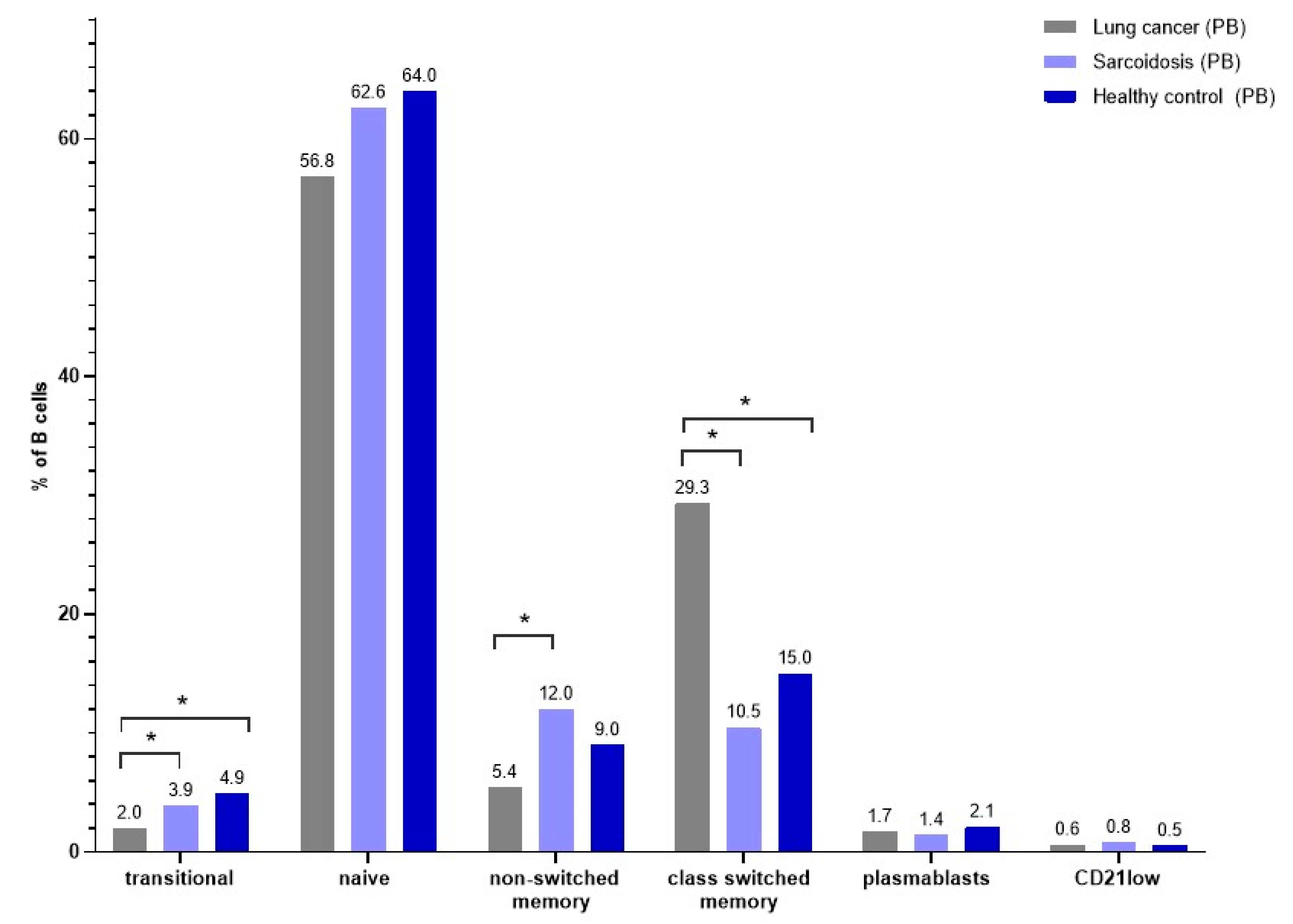
3.5. B-Cell Maturation Subsets in Peripheral Blood
3.6. New Lymphocyte Parameters in Peripheral Blood
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weeratunga, P.; Moller, D.R.; Ho, L.P. Immune mechanisms in fibrotic pulmonary sarcoidosis. Eur. Respir. Rev. 2022, 31, 220178. [Google Scholar] [CrossRef] [PubMed]
- Grunewald, J.; Grutters, J.C.; Arkema, E.V.; Saketkoo, L.A.; Moller, D.R.; Muller-Quernheim, J. Sarcoidosis. Nat. Rev. Dis. Primers 2019, 5, 45. [Google Scholar] [CrossRef] [PubMed]
- Grunewald, J.; Eklund, A. Role of CD4+ T cells in sarcoidosis. Proc. Am. Thorac. Soc. 2007, 4, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Broos, C.E.; van Nimwegen, M.; Hoogsteden, H.C.; Hendriks, R.W.; Kool, M.; van den Blink, B. Granuloma formation in pulmonary sarcoidosis. Front. Immunol. 2013, 4, 437. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, Y.; Han, J.; Yang, M.; Zhu, J.; Jin, T. Transitional B cells involved in autoimmunity and their impact on neuroimmunological diseases. J. Transl. Med. 2020, 18, 131. [Google Scholar] [CrossRef] [PubMed]
- Saussine, A.; Tazi, A.; Feuillet, S.; Rybojad, M.; Juillard, C.; Bergeron, A.; Dessirier, V.; Bouhidel, F.; Janin, A.; Bensussan, A.; et al. Active chronic sarcoidosis is characterized by increased transitional blood B cells, increased IL-10-producing regulatory B cells and high BAFF levels. PLoS ONE 2012, 7, e43588. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, A.; Maji, A.; Potdar, P.D.; Singh, N.; Parikh, P.; Bisht, B.; Mukherjee, A.; Paul, M.K. Lung cancer immunotherapy: Progress, pitfalls, and promises. Mol. Cancer 2023, 22, 40. [Google Scholar] [CrossRef]
- Sathish, G.; Monavarshini, L.K.; Sundaram, K.; Subramanian, S.; Kannayiram, G. Immunotherapy for lung cancer. Pathol. Res. Pract. 2024, 254, 155104. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef]
- Siegel, R.L.; Giaquinto, A.N.; Jemal, A. Cancer statistics, 2024. CA Cancer J. Clin. 2024, 74, 12–49. [Google Scholar] [CrossRef]
- Wang, S.S.; Liu, W.; Ly, D.; Xu, H.; Qu, L.; Zhang, L. Tumor-infiltrating B cells: Their role and application in anti-tumor immunity in lung cancer. Cell. Mol. Immunol. 2019, 16, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Leong, T.L.; Bryant, V.L. B cells in lung cancer-not just a bystander cell: A literature review. Transl. Lung. Cancer Res. 2021, 10, 2830–2841. [Google Scholar] [CrossRef] [PubMed]
- Yuseff, M.I.; Pierobon, P.; Reversat, A.; Lennon-Dumenil, A.M. How B cells capture, process and present antigens: A crucial role for cell polarity. Nat. Rev. Immunol. 2013, 13, 475–486. [Google Scholar] [CrossRef]
- LeBien, T.W.; Tedder, T.F. B lymphocytes: How they develop and function. Blood 2008, 112, 1570–1580. [Google Scholar] [CrossRef] [PubMed]
- Thorarinsdottir, K.; Camponeschi, A.; Gjertsson, I.; Martensson, I.L. CD21 -/low B cells: A Snapshot of a Unique B Cell Subset in Health and Disease. Scand. J. Immunol. 2015, 82, 254–261. [Google Scholar] [CrossRef]
- Gjertsson, I.; McGrath, S.; Grimstad, K.; Jonsson, C.A.; Camponeschi, A.; Thorarinsdottir, K.; Martensson, I.L. A close-up on the expanding landscape of CD21-/low B cells in humans. Clin. Exp. Immunol. 2022, 210, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Rutkowska, E.; Kwiecien, I.; Bednarek, J.; Sokolowski, R.; Raniszewska, A.; Jahnz-Rozyk, K.; Rzepecki, P. T Lymphocyte Maturation Profile in the EBUS-TBNA Lymph Node Depending on the DLCO Parameter in Patients with Pulmonary Sarcoidosis. Cells 2021, 10, 3404. [Google Scholar] [CrossRef]
- Raniszewska, A.; Vroman, H.; Dumoulin, D.; Cornelissen, R.; Aerts, J.; Domagala-Kulawik, J. PD-L1(+) lung cancer stem cells modify the metastatic lymph-node immunomicroenvironment in nsclc patients. Cancer Immunol. Immunother. 2021, 70, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Kwiecien, I.; Rutkowska, E.; Sokolowski, R.; Bednarek, J.; Raniszewska, A.; Jahnz-Rozyk, K.; Rzepecki, P.; Domagala-Kulawik, J. Effector Memory T Cells and CD45RO+ Regulatory T Cells in Metastatic vs. Non-Metastatic Lymph Nodes in Lung Cancer Patients. Front. Immunol. 2022, 13, 864497. [Google Scholar] [CrossRef]
- Vilmann, P.; Clementsen, P.F.; Colella, S.; Siemsen, M.; De Leyn, P.; Dumonceau, J.M.; Herth, F.J.; Larghi, A.; Vazquez-Sequeiros, E.; Hassan, C.; et al. Combined endobronchial and oesophageal endosonography for the diagnosis and staging of lung cancer. European Society of Gastrointestinal Endoscopy (ESGE) Guideline, in cooperation with the European Respiratory Society (ERS) and the European Society of Thoracic Surgeons (ESTS). Eur. Respir. J. 2015, 46, 40–60. [Google Scholar] [CrossRef]
- Wahidi, M.M.; Herth, F.; Yasufuku, K.; Shepherd, R.W.; Yarmus, L.; Chawla, M.; Lamb, C.; Casey, K.R.; Patel, S.; Silvestri, G.A.; et al. Technical Aspects of Endobronchial Ultrasound-Guided Transbronchial Needle Aspiration: CHEST Guideline and Expert Panel Report. Chest 2016, 149, 816–835. [Google Scholar] [CrossRef]
- Sysmex, E.G. Novel Haematological Parameters for Rapidly Monitoring the Immune System Response; Norderstedt, Germany. Sysmex Eur. GmbH 2017, 27, 1–5. [Google Scholar]
- Henriot, I.; Launay, E.; Boubaya, M.; Cremet, L.; Illiaquer, M.; Caillon, H.; Desjonqueres, A.; Gillet, B.; Bene, M.C.; Eveillard, M. New parameters on the hematology analyzer XN-10 (SysmexTM) allow to distinguish childhood bacterial and viral infections. Int. J. Lab. Hematol. 2017, 39, 14–20. [Google Scholar] [CrossRef]
- Poutakidou, D.; Ruth, I.; Gulbis, B. Differential diagnosis of lymphocytosis in routine laboratory practice: Contribution of lymphocyte parameters using the Sysmex-XN9000 haematology analyzer. Int. J. Lab. Hematol. 2023, 45, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Furundarena, J.R.; Uranga, A.; Sainz, M.R.; Gonzalez, C.; Uresandi, N.; Argoitia, N.; Araiz, M. Usefulness of the lymphocyte positional parameters in the Sysmex XN haematology analyser in lymphoproliferative disorders and mononucleosis syndrome. Int. J. Lab. Hematol. 2018, 40, 41–48. [Google Scholar] [CrossRef]
- Graham, B.L.; Brusasco, V.; Burgos, F.; Cooper, B.G.; Jensen, R.; Kendrick, A.; MacIntyre, N.R.; Thompson, B.R.; Wanger, J. 2017 ERS/ATS standards for single-breath carbon monoxide uptake in the lung. Eur. Respir. J. 2017, 49, 1600016. [Google Scholar] [CrossRef]
- Nicholson, A.G.; Tsao, M.S.; Beasley, M.B.; Borczuk, A.C.; Brambilla, E.; Cooper, W.A.; Dacic, S.; Jain, D.; Kerr, K.M.; Lantuejoul, S.; et al. The 2021 WHO Classification of Lung Tumors: Impact of Advances Since 2015. J. Thorac. Oncol. 2022, 17, 362–387. [Google Scholar] [CrossRef] [PubMed]
- Detterbeck, F.C.; Boffa, D.J.; Kim, A.W.; Tanoue, L.T. The Eighth Edition Lung Cancer Stage Classification. Chest 2017, 151, 193–203. [Google Scholar] [CrossRef]
- Li, H.Y.; Wang, Y.Y.; Liu, H.; Liu, H.X.; Jiang, L.Y.; Han, Y.C.; Zhou, W.Y.; Mao, T.; Fang, W.T. The ninth edition of TNM staging for lung cancer: Precise staging for precise diagnosis and treatment. Zhonghua. Wai. Ke. Za. Zhi. 2024, 62, 537–542. [Google Scholar] [CrossRef]
- van der Burg, M.; Kalina, T.; Perez-Andres, M.; Vlkova, M.; Lopez-Granados, E.; Blanco, E.; Bonroy, C.; Sousa, A.E.; Kienzler, A.K.; Wentink, M.; et al. The EuroFlow PID Orientation Tube for Flow Cytometric Diagnostic Screening of Primary Immunodeficiencies of the Lymphoid System. Front. Immunol. 2019, 10, 246. [Google Scholar] [CrossRef]
- Chen, C.; Luo, N.; Dai, F.; Zhou, W.; Wu, X.; Zhang, J. Advance in pathogenesis of sarcoidosis: Triggers and progression. Heliyon. 2024, 10, e27612. [Google Scholar] [CrossRef] [PubMed]
- Hinze, C.H.; Colbert, R.A. B-cell depletion in Wegener’s granulomatosis. Clin. Rev. Allergy. Immunol. 2008, 34, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Domagala-Kulawik, J.; Osinska, I.; Hoser, G. Mechanisms of immune response regulation in lung cancer. Transl. Lung. Cancer. Res. 2014, 3, 15–22. [Google Scholar] [PubMed]
- Kwiecien, I.; Rutkowska, E.; Raniszewska, A.; Rzepecki, P.; Domagala-Kulawik, J. Modulation of the immune response by heterogeneous monocytes and dendritic cells in lung cancer. World. J. Clin. Oncol. 2021, 12, 966–982. [Google Scholar] [CrossRef]
- Kwiecien, I.; Rutkowska, E.; Raniszewska, A.; Rzeszotarska, A.; Polubiec-Kownacka, M.; Domagala-Kulawik, J.; Korsak, J.; Rzepecki, P. Flow Cytometric Analysis of Macrophages and Cytokines Profile in the Bronchoalveolar Lavage Fluid in Patients with Lung Cancer. Cancers 2023, 15, 5175. [Google Scholar] [CrossRef] [PubMed]
- Kwiecien, I.; Rutkowska, E.; Raniszewska, A.; Sokolowski, R.; Bednarek, J.; Jahnz-Rozyk, K.; Rzepecki, P.; Domagala-Kulawik, J. Immunosuppressive properties of human PD-1 +, PDL-1 + and CD80 + dendritic cells from lymph nodes aspirates of lung cancer patients. Cancer Immunol. Immunother. 2022, 71, 2469–2483. [Google Scholar] [CrossRef] [PubMed]
- Kwiecien, I.; Rutkowska, E.; Polubiec-Kownacka, M.; Raniszewska, A.; Rzepecki, P.; Domagala-Kulawik, J. Identification of PD-1 ligands: PD-L1 and PD-L2 on macrophages in lung cancer milieu by flow cytometry. Transl. Lung Cancer Res. 2021, 10, 1679–1689. [Google Scholar] [CrossRef]
- Kwiecien, I.; Skirecki, T.; Polubiec-Kownacka, M.; Raniszewska, A.; Domagala-Kulawik, J. Immunophenotype of T Cells Expressing Programmed Death-1 and Cytotoxic T Cell Antigen-4 in Early Lung Cancer: Local vs. Systemic Immune Response. Cancers 2019, 11, 567. [Google Scholar] [CrossRef]
- Kwiecien, I.; Polubiec-Kownacka, M.; Dziedzic, D.; Wolosz, D.; Rzepecki, P.; Domagala-Kulawik, J. CD163 and CCR7 as markers for macrophage polarization in lung cancer microenvironment. Cent. Eur. J. Immunol. 2019, 44, 395–402. [Google Scholar] [CrossRef]
- Kwiecien, I.; Stelmaszczyk-Emmel, A.; Polubiec-Kownacka, M.; Dziedzic, D.; Domagala-Kulawik, J. Elevated regulatory T cells, surface and intracellular CTLA-4 expression and interleukin-17 in the lung cancer microenvironment in humans. Cancer Immunol. Immunother. 2017, 66, 161–170. [Google Scholar] [CrossRef]
- Osinska, I.; Stelmaszczyk-Emmel, A.; Polubiec-Kownacka, M.; Dziedzic, D.; Domagala-Kulawik, J. CD4+/CD25(high)/FoxP3+/CD127- regulatory T cells in bronchoalveolar lavage fluid of lung cancer patients. Hum. Immunol. 2016, 77, 912–915. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.J.; Richter, A.; Drayson, M.T.; Middleton, G.W. The role of B lymphocytes in the immuno-biology of non-small-cell lung cancer. Cancer. Immunol. Immunother. 2020, 69, 325–342. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.J.; Khan, N.; Richter, A.; Naidu, B.; Drayson, M.T.; Middleton, G.W. Deep immune B and plasma cell repertoire in non-small cell lung cancer. Front. Immunol. 2023, 14, 1198665. [Google Scholar] [CrossRef] [PubMed]
- Hao, D.; Han, G.; Sinjab, A.; Gomez-Bolanos, L.I.; Lazcano, R.; Serrano, A.; Hernandez, S.D.; Dai, E.; Cao, X.; Hu, J.; et al. The Single-Cell Immunogenomic Landscape of B and Plasma Cells in Early-Stage Lung Adenocarcinoma. Cancer Discov. 2022, 12, 2626–2645. [Google Scholar] [CrossRef] [PubMed]
- Asaoka, M.; Wu, R.; Ishikawa, T. Abstract 1408: Increased infiltrating class-switched memory B cell in breast cancer tumors were associated with favorable prognostic outcome. Cancer Res. 2023, 83, 1408. [Google Scholar] [CrossRef]
- Kamphuis, L.S.; van Zelm, M.C.; Lam, K.H.; Rimmelzwaan, G.F.; Baarsma, G.S.; Dik, W.A.; Thio, H.B.; van Daele, P.L.; van Velthoven, M.E.; Batstra, M.R.; et al. Perigranuloma localization and abnormal maturation of B cells: Emerging key players in sarcoidosis? Am. J. Respir. Crit. Care Med. 2013, 187, 406–416. [Google Scholar] [CrossRef]
- Kudryavtsev, I.; Serebriakova, M.; Starshinova, A.; Zinchenko, Y.; Basantsova, N.; Malkova, A.; Soprun, L.; Churilov, L.P.; Toubi, E.; Yablonskiy, P.; et al. Imbalance in B cell and T Follicular Helper Cell Subsets in Pulmonary Sarcoidosis. Sci. Rep. 2020, 10, 1059. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.S.; Barber, L.; Akula, S.M.; Sigounas, G.; Kataria, Y.P.; Arce, S. Disturbed homeostasis and multiple signaling defects in the peripheral blood B-cell compartment of patients with severe chronic sarcoidosis. Clin. Vacc. Immunol. 2011, 18, 1306–1316. [Google Scholar] [CrossRef] [PubMed]
- Wehr, C.; Kivioja, T.; Schmitt, C.; Ferry, B.; Witte, T.; Eren, E.; Vlkova, M.; Hernandez, M.; Detkova, D.; Bos, P.R.; et al. The EUROclass trial: Defining subgroups in common variable immunodeficiency. Blood 2008, 111, 77–85. [Google Scholar] [CrossRef]
- Wiesik-Szewczyk, E.; Rutkowska, E.; Kwiecien, I.; Korzeniowska, M.; Soldacki, D.; Jahnz-Rozyk, K. Patients with Common Variable Immunodeficiency Complicated by Autoimmune Phenomena Have Lymphopenia and Reduced Treg, Th17, and NK Cells. J. Clin. Med. 2021, 10, 3356. [Google Scholar] [CrossRef]
- Zhang, H.; Costabel, U.; Dai, H. The Role of Diverse Immune Cells in Sarcoidosis. Front. Immunol. 2021, 12, 788502. [Google Scholar] [CrossRef] [PubMed]
- Ueda-Hayakawa, I.; Tanimura, H.; Osawa, M.; Iwasaka, H.; Ohe, S.; Yamazaki, F.; Mizuno, K.; Okamoto, H. Elevated serum BAFF levels in patients with sarcoidosis: Association with disease activity. Rheumatology 2013, 52, 1658–1666. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Ding, Y.; Yi, X.; Zheng, J. CD19+ B cell subsets in the peripheral blood and skin lesions of psoriasis patients and their correlations with disease severity. Braz. J. Med. Biol. Res. 2016, 49, e5374. [Google Scholar] [CrossRef] [PubMed]
- Bosello, S.; Angelucci, C.; Lama, G.; Alivernini, S.; Proietti, G.; Tolusso, B.; Sica, G.; Gremese, E.; Ferraccioli, G. Characterization of inflammatory cell infiltrate of scleroderma skin: B cells and skin score progression. Arthr. Res. Ther. 2018, 20, 75. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.C.; Hosler, G.A.; Chong, B.F. Changes in T cell and B cell composition in discoid lupus erythematosus skin at different stages. J. Dermatol. Sci. 2017, 85, 247–249. [Google Scholar] [CrossRef] [PubMed]
- Parodis, I.; Long, X.; Karlsson, M.C.I.; Huang, X. B Cell Tolerance and Targeted Therapies in SLE. J. Clin. Med. 2023, 12, 6268. [Google Scholar] [CrossRef] [PubMed]
- Meffre, E.; O’Connor, K.C. Impaired B-cell tolerance checkpoints promote the development of autoimmune diseases and pathogenic autoantibodies. Immunol. Rev. 2019, 292, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.B.; Silverman, M.; Monroe, J.G. Transitional B cells: Step by step towards immune competence. Trends. Immunol. 2003, 24, 343–349. [Google Scholar] [CrossRef]
- Sims, G.P.; Ettinger, R.; Shirota, Y.; Yarboro, C.H.; Illei, G.G.; Lipsky, P.E. Identification and characterization of circulating human transitional B cells. Blood 2005, 105, 4390–4398. [Google Scholar] [CrossRef]
- Velounias, R.L.; Tull, T.J. Human B-cell subset identification and changes in inflammatory diseases. Clin. Exp. Immunol. 2022, 210, 201–216. [Google Scholar] [CrossRef]
- Tan, R.; Nie, M.; Long, W. The role of B cells in cancer development. Front. Oncol. 2022, 12, 958756. [Google Scholar] [CrossRef] [PubMed]
- Keller, B.; Strohmeier, V.; Harder, I.; Unger, S.; Payne, K.J.; Andrieux, G.; Boerries, M.; Felixberger, P.T.; Landry, J.J.M.; Nieters, A.; et al. The expansion of human T-bet(high)CD21(low) B cells is T cell dependent. Sci. Immunol. 2021, 6, eabh0891. [Google Scholar] [CrossRef] [PubMed]
- Sutton, H.J.; Aye, R.; Idris, A.H.; Vistein, R.; Nduati, E.; Kai, O.; Mwacharo, J.; Li, X.; Gao, X.; Andrews, T.D.; et al. Atypical B cells are part of an alternative lineage of B cells that participates in responses to vaccination and infection in humans. Cell. Rep. 2021, 34, 108684. [Google Scholar] [CrossRef] [PubMed]
- Reincke, M.E.; Payne, K.J.; Harder, I.; Strohmeier, V.; Voll, R.E.; Warnatz, K.; Keller, B. The Antigen Presenting Potential of CD21(low) B Cells. Front. Immunol. 2020, 11, 535784. [Google Scholar] [CrossRef]
- Thanou, A.; Jupe, E.; Purushothaman, M.; Niewold, T.B.; Munroe, M.E. Clinical disease activity and flare in SLE: Current concepts and novel biomarkers. J. Autoimmun. 2021, 119, 102615. [Google Scholar] [CrossRef] [PubMed]
- Freudenhammer, M.; Voll, R.E.; Binder, S.C.; Keller, B.; Warnatz, K. Naive- and Memory-like CD21(low) B Cell Subsets Share Core Phenotypic and Signaling Characteristics in Systemic Autoimmune Disorders. J. Immunol. 2020, 205, 2016–2025. [Google Scholar] [CrossRef]
- Centuori, S.M.; Gomes, C.J.; Kim, S.S.; Putnam, C.W.; Larsen, B.T.; Garland, L.L.; Mount, D.W.; Martinez, J.D. Double-negative (CD27(-)IgD(-)) B cells are expanded in NSCLC and inversely correlate with affinity-matured B cell populations. J. Transl. Med. 2018, 16, 30. [Google Scholar] [CrossRef]
- Belderbos, R.A.; Corneth, O.B.J.; Dumoulin, D.; Hendriks, R.W.; Aerts, J.; Willemsen, M. Atypical B cells (CD21-CD27-IgD-) correlate with lack of response to checkpoint inhibitor therapy in NSCLC. Eur. J. Cancer 2024, 196, 113428. [Google Scholar] [CrossRef]
- Klein, J.; Wood, J.; Jaycox, J.; Lu, P.; Dhodapkar, R.M.; Gehlhausen, J.R.; Tabachnikova, A.; Tabacof, L.; Malik, A.A.; Kamath, K.; et al. Distinguishing features of Long COVID identified through immune profiling. Nature 2023, 623, 139–148. [Google Scholar] [CrossRef]
- Holla, P.; Dizon, B.; Ambegaonkar, A.A.; Rogel, N.; Goldschmidt, E.; Boddapati, A.K.; Sohn, H.; Sturdevant, D.; Austin, J.W.; Kardava, L.; et al. Shared transcriptional profiles of atypical B cells suggest common drivers of expansion and function in malaria, HIV, and autoimmunity. Sci. Adv. 2021, 7, eabg8384. [Google Scholar] [CrossRef]
- Portugal, S.; Obeng-Adjei, N.; Moir, S.; Crompton, P.D.; Pierce, S.K. Atypical memory B cells in human chronic infectious diseases: An interim report. Cell. Immunol. 2017, 321, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Doi, H.; Tanoue, S.; Kaplan, D.E. Peripheral CD27-CD21- B-cells represent an exhausted lymphocyte population in hepatitis C cirrhosis. Clin. Immunol. 2014, 150, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Helmink, B.A.; Reddy, S.M.; Gao, J.; Zhang, S.; Basar, R.; Thakur, R.; Yizhak, K.; Sade-Feldman, M.; Blando, J.; Han, G.; et al. B cells and tertiary lymphoid structures promote immunotherapy response. Nature 2020, 577, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Petitprez, F.; de Reynies, A.; Keung, E.Z.; Chen, T.W.; Sun, C.M.; Calderaro, J.; Jeng, Y.M.; Hsiao, L.P.; Lacroix, L.; Bougouin, A.; et al. B cells are associated with survival and immunotherapy response in sarcoma. Nature 2020, 577, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Rutkowska, E.; Kwiecien, I.; Kulik, K.; Chelstowska, B.; Klos, K.; Rzepecki, P.; Chcialowski, A. Usefulness of the New Hematological Parameter: Reactive Lymphocytes RE-LYMP with Flow Cytometry Markers of Inflammation in COVID-19. Cells 2021, 10, 82. [Google Scholar] [CrossRef] [PubMed]
- Di Luise, D.; Giannotta, J.A.; Ammirabile, M.; De Zordi, V.; Torricelli, S.; Bottalico, S.; Chiaretto, M.L.; Fattizzo, B.; Migliorini, A.C.; Ceriotti, F. Cell Population Data NE-WX, NE-FSC, LY-Y of Sysmex XN-9000 can provide additional information to differentiate macrocytic anaemia from myelodysplastic syndrome: A preliminary study. Int. J. Lab. Hematol. 2022, 44, e40–e43. [Google Scholar] [CrossRef]
- Solta, A.; Ernhofer, B.; Boettiger, K.; Megyesfalvi, Z.; Heeke, S.; Hoda, M.A.; Lang, C.; Aigner, C.; Hirsch, F.R.; Schelch, K.; et al. Small cells—Big issues: Biological implications and preclinical advancements in small cell lung cancer. Mol. Cancer 2024, 23, 41. [Google Scholar] [CrossRef]
- Araghi, M.; Mannani, R.; Heidarnejad Maleki, A.; Hamidi, A.; Rostami, S.; Safa, S.H.; Faramarzi, F.; Khorasani, S.; Alimohammadi, M.; Tahmasebi, S.; et al. Recent advances in non-small cell lung cancer targeted therapy; an update review. Cancer Cell Int. 2023, 23, 162. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SA | LC | HC | |
|---|---|---|---|
| Number of patients | 34 | 33 | 20 |
| Sex F/M (n) | 9/25 | 19/14 | 19/1 |
| Age (mean ± SD years) | 42.5 ± 13.0 | 67.0 ± 8.5 | 51.5 ± 9.7 |
| LN 4/7/10/11/ * | 2/16/1/15/0 | 15/6/1/5/6 | n/a |
| Stage I/II/III/IV | 11/23/ n/a /n/a | 2/4/17/10 | n/a |
| SA patients characteristic | |||
| BMI (mean ± SD) | 29.6 ± 5.0 | ||
| DLCO (>80%/<80%) | 17/17 | ||
| DLCO > 80% (mean ± SD) | 94.2 ± 7.6 | ||
| DLCO < 80% (mean ± SD) | 70.6 ± 7.0 | ||
| FVC women [L] | 3.10 ± 0.42 | ||
| FVC men [L] | 4.22 ± 0.63 | ||
| FEV1 women [L] | 2.50 ± 0.31 | ||
| FEV1 men [L] | 3.10 ± 0.45 | ||
| FEV1/FVC | 82.70 ± 6.50 | ||
| Parameter | Parameter Description |
|---|---|
| AS-LYMP | quantifies the activated B lymphocytes that synthesize antibodies. |
| RE-LYMP | all activated lymphocytes with a higher level of fluorescence than resting lymphocytes |
| LY-X | laterally scattered light intensity; lymphocyte complexity |
| LY-Y | intensity of fluorescent light; reflects lymphocyte fluorescence |
| LY-Z | intensity of frontally scattered light; reflects lymphocyte size |
| LY-WX | laterally scattered light intensity; width of dispersion of lymphocyte complexity |
| LY-WY | intensity of fluorescent light; width of dispersion of lymphocyte fluorescence |
| Leukocytes Subpopulation: [% of All Cells] | Lung Cancer Median (Q1–Q3) | Sarcoidosis Median (Q1–Q3) | * p < 0.05 Mann–Whitney U Test) |
|---|---|---|---|
| Lymphocytes | 6.0 (1.5–8.4) | 88.6 (82.1–98.3) | * p < 0.0001 |
| Lymphocytes T | 3.4 (0.9–6.4) | 50.2 (39.4–63.5) | * p < 0.0001 |
| CD4 | 1.4 (0.5–4.0) | 37.0 (27.1–45.7) | * p < 0.0001 |
| CD8 | 1.4 (0.4–3.7) | 12.9 (8.8–16.2) | * p < 0.0001 |
| Ratio CD4/CD8 | 1.2 (1.0–1.8) | 3.0 (2.2–4.6) | * p < 0.0001 |
| Lymphocytes B | 0.6 (0.3–1.5) | 27.5 (17.6–43.3) | * p < 0.0001 |
| NK cells | 0.2 (0.0–0.8) | 1.4 (0.0–2.8) | * p = 0.0073 |
| Neutrophils | 2.9 (0.0–14.2) | 4.2 (0.0–13.4) | p = 0.7993 |
| Monocytes | 0.0 (0.0–0.0) | 2.8 (0.0–9.5) | * p < 0.0001 |
| Tumor cells | 88.5 (68.4–94.3) | - | - |
| [% of all lymphocytes] | |||
| Lymphocytes T | 75.6 (57.1–81.7) | 70.0 (54.5–76.0) | p = 0.1084 |
| CD4 | 37.9 (24.8–48.8) | 49.8 (39.1–54.5) | * p = 0.0097 |
| CD8 | 32.4 (24.7–42.1) | 16.1 (11.1–23.6) | * p = 0.0002 |
| Lymphocytes B | 16.4 (11.9–26.1) | 26.9 (21.1–45.5) | * p = 0.0039 |
| NK cells | 2.6 (0.0–12.0) | 1.7 (0.1–3.8) | p = 0.3848 |
| Lung Cancer Median (Q1–Q3) | Sarcoidosis Median (Q1–Q3) | * p < 0.05 Mann–Whitney U Test) | |
|---|---|---|---|
| B-cell subpopulation: [% of B lymphocytes] | |||
| Naïve | 31.8 (20.1–53.3) | 50.4 (42.5–59.7) | * p = 0.0033 |
| Non-switched memory | 4.7 (3.1–7.0) | 5.4 (2.5–14.8) | p = 0.3777 |
| Class-switched memory | 49.9 (26.3–62.7) | 26.0 (15.0–39.6) | * p = 0.0019 |
| Transitional | 2.4 (1.1–4.7) | 3.4 (1.2–8.7) | p = 0.1449 |
| Plasmablast | 2.8 (1.5–5.6) | 1.7 (0.9–5.2) | p = 0.3533 |
| CD21low cells | 0.5 (0.0–0.9) | 0.8 (0.2–1.6) | * p = 0.0377 |
| Hematological parameter: | |||
| LY-X [ch] | 85.6 (83.0–96.6) | 81.7 (79.9–82.8) | * p < 0.0001 |
| LY-Y [ch] | 48.7 (44.2–54.4) | 68.0 (65.5–73.1) | * p < 0.0001 |
| Lung Cancer (A) Median (Q1–Q3) | Sarcoidosis (B) Median (Q1–Q3) | Healthy Control (C) Median (Q1–Q3) | * p < 0.05 Group A-B-C ANOVA, Kruskal–Wallis | * p < 0.05 Group, in Groups Post-Hoc | ||
|---|---|---|---|---|---|---|
| WBC | [k/µL] | 8310 (6500–11870) | 5730 (4720–7460) | 5365 (4695–6720) | * p = 0.0006 | A.-C. p = 0.0041 A.-B. p = 0.0026 |
| Lymphocytes | [%] | 14.8 (11.6–22.4) | 25.5 (16.0–30.1) | 41.3 (36.9–45.5) | * p < 0.0001 | A.-C. p < 0.0001 A.-B. p = 0.0174 B.-C. p = 0.0001 |
| T Lymphocytes | [%] | 10.7 (6.7–17.1) | 17.0 (10.0–20.4) | 31.4 (26.4–35.4) | * p < 0.0001 | A.-C. p < 0.0001 B.-C. p < 0.0001 |
| CD4 | [%] | 4.9 (3.4–14.1) | 10.1 (6.0–12.6) | 19.7 (17.0–22.0) | * p < 0.0001 | A.-C. p = 0.0075 B.-C. p < 0.0001 |
| CD8 | [%] | 2.9 (1.4–3.8) | 6.0 (4.2–8.5) | 11.4 (8.3–14.7) | * p < 0.0001 | A.-C. p = 0.0014 B.-C. p = 0.0002 |
| Ratio CD4/CD8 | 2.4 (1.3–4.9) | 1.5 (0.9–2.3) | 1.7 (1.3–2.3) | p = 0.1944 | - | |
| B Lymphocytes | [%] | 2.2 (1.2–2.9) | 2.8 (1.3–3.7) | 3.9 (2.7–5.6) | * p = 0.0089 | A.-C. p = 0.0065 |
| NK cells | [%] | 1.7 (0.8–3.5) | 2.9 (1.4–3.7) | 5.7 (2.7–7.7) | * p = 0.0002 | A.-C. p = 0.0001 B.-C. p = 0.0134 |
| Neutrophils | [%] | 77.3 (69.2–80.5) | 69.0 (61.5–72.0) | 47.1 (42.4–52.3) | * p < 0.0001 | A.-C. p < 0.0001 A.-B. p = 0.0191 B.-C. p < 0.0001 |
| Monocytes | [%] | 6.8 (5.6–7.8) | 9.8 (6.5–11.8) | 7.9 (6.7–10.6) | * p = 0.0045 | A.-B. p = 0.0046 |
| Cells Subsets: [% of B Lymphocytes] | Lung Cancer (A) Median (Q1–Q3) | Sarcoidosis (B) Median (Q1–Q3) | Healthy Control (C) Median (Q1–Q3) | * p < 0.05 Group A-B-C ANOVA, Kruskal–Wallis | * p < 0.05 Group, in Groups Post-Hoc |
|---|---|---|---|---|---|
| B-cell subsets: [% of B lymphocytes] | |||||
| Naïve | 56.8 (45.9–68.7) | 62.6 (56.5–73.9) | 64.0 (57.9–75.5) | p = 0.1216 | - |
| Non-switched memory | 5.4 (4.3–7.8) | 12.0 (7.3–20.8) | 9.0 (6.6–11.7) | * p = 0.0002 | A.-B. p = 0.0001 |
| Class-switched memory | 29.3 (17.1–37.8) | 10.5 (6.6–15.2) | 15.0 (12.6–21.5) | * p < 0.0001 | A.-B. p < 0.0001 A.-C. p = 0.0103 |
| Transitional | 2.0 (0.9–2.9) | 3.9 (2.7–9.2) | 4.9 (2.8–6.0) | * p < 0.0001 | A.-B. p = 0.0003 A.-C. p = 0.0004 |
| Plasmablast | 1.7 (4.3–7.8) | 1.4 (0.8–4.3) | 2.1 (0.7–3.3) | p = 0.9488 | - |
| CD21low | 0.6 (0.3–1.4) | 0.8 (0.3–1.7) | 0.5 (0.3–0.7) | p = 0.4567 | - |
| Hematological parameters: | |||||
| RE-LYMP [×103/µL] | 0.1 (0.1–0.2) | 0.1 (0.0–0.1) | 0.1 (0.0–0.1) | * p = 0.0017 | A.-B. p = 0.0013 |
| RE-LYMP [% of all cells] | 1.9 (1.4–2.4) | 0.9 (0.6–1.1) | 1.0 (0.5–1.4) | * p < 0.0001 | A.-B. p = 0.0001 A.-C. p = 0.0003 |
| RE-LYMP [% of lymphocytes] | 5.9 (4.2–7.6) | 4.0 (2.5–5.7) | 4.4 (3.0–6.5) | * p = 0.0378 | A.-B. p = 0.0316 |
| AS-LYMP [×103/µL] | 0.0 (0.0–0.1) | 0.0 (0.0–0.0) | 0.0 (0.0–0.0) | * p = 0.0002 | A.-C. p = 0.0030 |
| AS-LYMP [% of all cells] | 0.1 (0.0–0.2) | 0.0 (0.0–0.0) | 0.0 (0.0–0.0) | * p = 0.0003 | A.-C. p = 0.0038 |
| AS-LYMP [% of lymphocytes] | 0.4 (0.0–0.9) | 0.0 (0.0–0.0) | 0.0 (0.0–0.0) | * p = 0.0002 | A.-C. p = 0.0032 |
| LY-X [ch] | 78.5 (77.8–79.6) | 78.4 (77.5–79.7) | 82.5 (82.1–83.5) | * p < 0.0001 | A.-C. p < 0.0001 B.-C. p < 0.0001 |
| LY-Y [ch] | 68.5 (66.0–72.0) | 67.2 (65.9–70.6) | 72.3 (70.4–73.6) | * p = 0.0028 | A.-C. p = 0.0283 B.-C. p = 0.0022 |
| LY-Z [ch] | 58.6( 57.6–59.9) | 58.4 (58.0–59.0) | 61.6 (61.0–62.1) | * p < 0.0001 | A.-C. p < 0.0001 B.-C. p < 0.0001 |
| LY-WX | 508.0 (491.0–540.0) | 492.0 (464.0–530.0) | 470.5 (452.0–500.5) | * p = 0.0032 | A.-C. p = 0.0021 |
| LY-WY | 850.0 (813.0–990.0) | 853.0 (804.0–928.0) | 873.0 (806.0–907.5) | p = 0.7664 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raniszewska, A.; Kwiecień, I.; Rutkowska, E.; Bednarek, J.; Sokołowski, R.; Miklusz, P.; Rzepecki, P.; Jahnz-Różyk, K. Imbalance of B-Cell Subpopulations in the Microenvironment of Sarcoidosis or Lung Cancer. Cells 2024, 13, 1274. https://doi.org/10.3390/cells13151274
Raniszewska A, Kwiecień I, Rutkowska E, Bednarek J, Sokołowski R, Miklusz P, Rzepecki P, Jahnz-Różyk K. Imbalance of B-Cell Subpopulations in the Microenvironment of Sarcoidosis or Lung Cancer. Cells. 2024; 13(15):1274. https://doi.org/10.3390/cells13151274
Chicago/Turabian StyleRaniszewska, Agata, Iwona Kwiecień, Elżbieta Rutkowska, Joanna Bednarek, Rafał Sokołowski, Piotr Miklusz, Piotr Rzepecki, and Karina Jahnz-Różyk. 2024. "Imbalance of B-Cell Subpopulations in the Microenvironment of Sarcoidosis or Lung Cancer" Cells 13, no. 15: 1274. https://doi.org/10.3390/cells13151274
APA StyleRaniszewska, A., Kwiecień, I., Rutkowska, E., Bednarek, J., Sokołowski, R., Miklusz, P., Rzepecki, P., & Jahnz-Różyk, K. (2024). Imbalance of B-Cell Subpopulations in the Microenvironment of Sarcoidosis or Lung Cancer. Cells, 13(15), 1274. https://doi.org/10.3390/cells13151274