
Fibroblast Activation Protein Is Expressed by Altered Osteoprogenitors and Associated to Disease Burden in Fibrous Dysplasia
 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
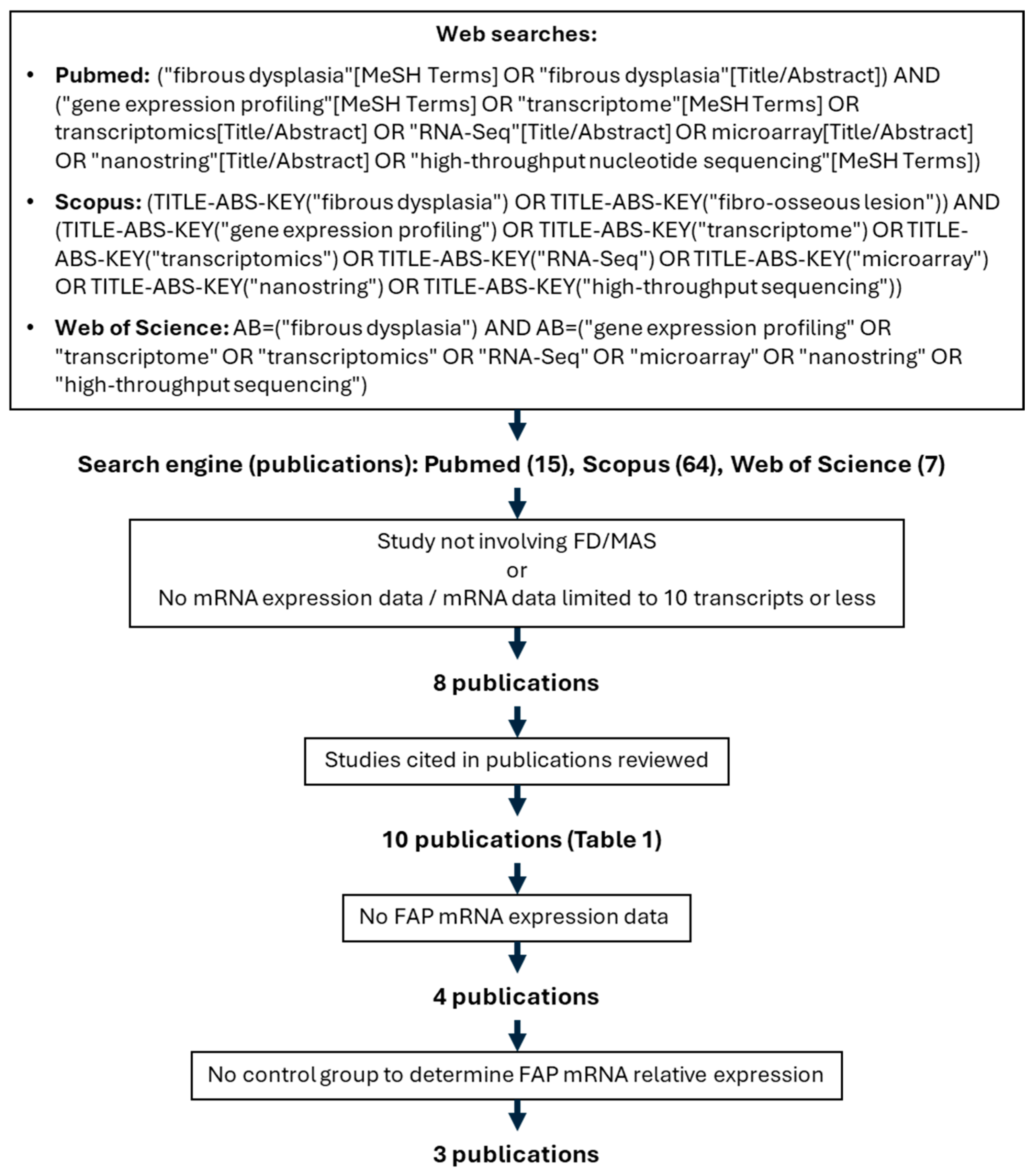
2.1. Literature Search
2.2. Human Specimens
2.3. FAPα Determination
2.4. FAP/Fap mRNA Expression Level Analyses
2.5. Statistical Analysis
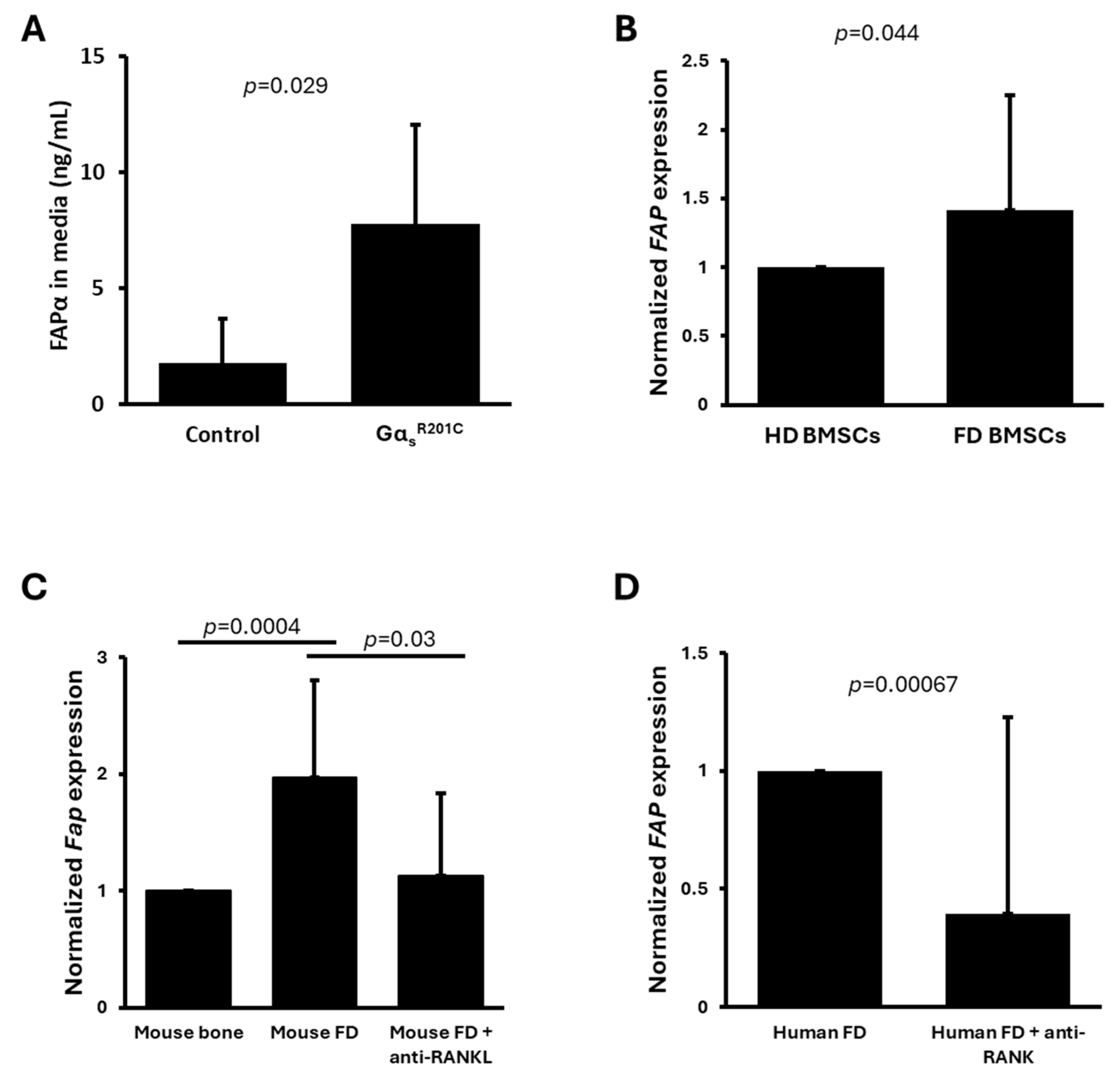
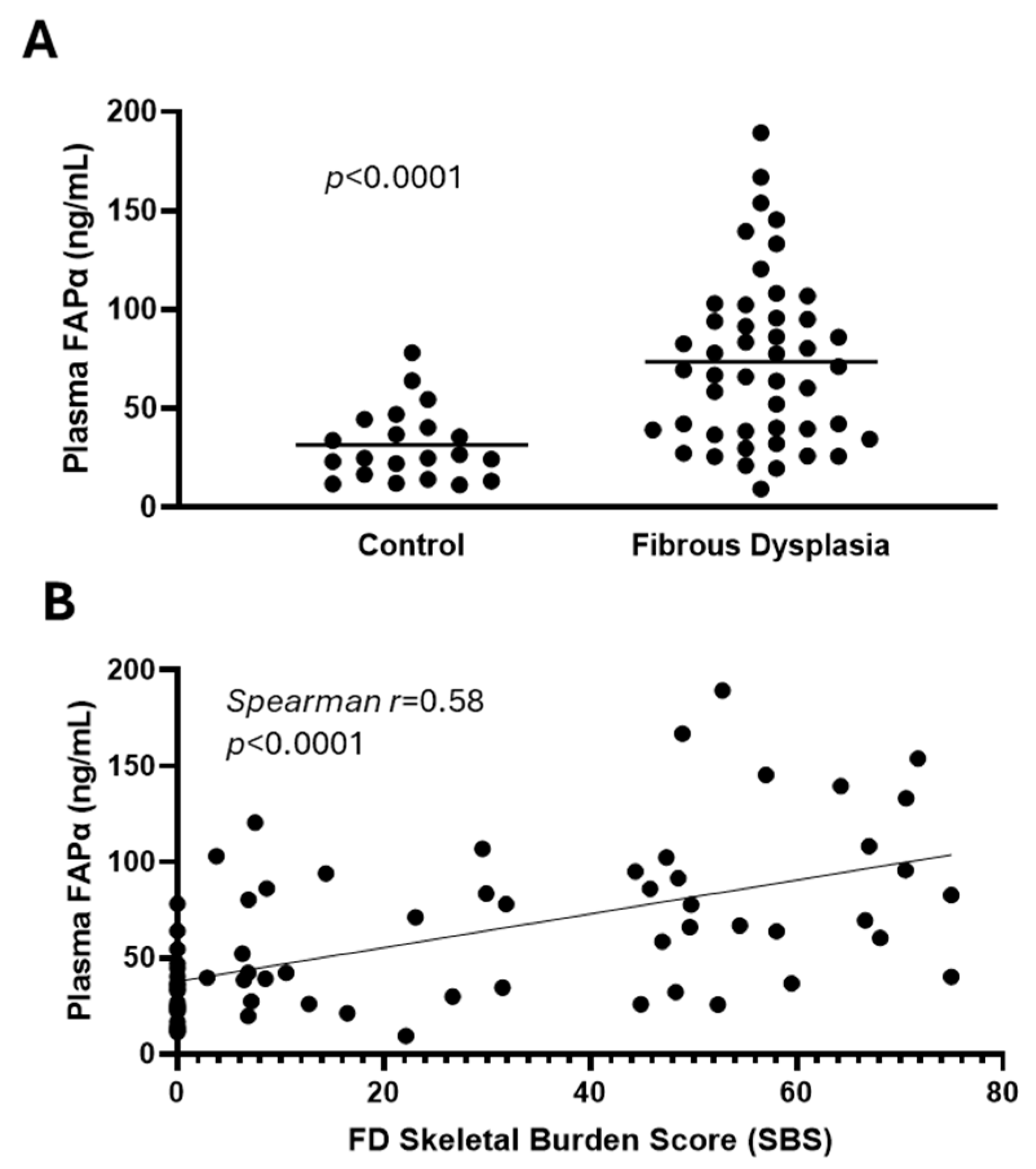
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Castro, L.F.; Whitlock, J.M.; Michel, Z.; Pan, K.; Taylor, J.; Szymczuk, V.; Boyce, B.; Martin, D.; Kram, V.; Galisteo, R.; et al. RANKL inhibition reduces lesional cellularity and Gαs variant expression and enables osteogenic maturation in fibrous dysplasia. Bone Res. 2024, 12, 10. [Google Scholar] [CrossRef]
- de Castro, L.F.; Michel, Z.; Pan, K.; Taylor, J.; Szymczuk, V.; Paravastu, S.; Saboury, B.; Papadakis, G.Z.; Li, X.; Milligan, K.; et al. Safety and Efficacy of Denosumab for Fibrous Dysplasia of Bone. N. Engl. J. Med. 2023, 388, 766–768. [Google Scholar] [CrossRef]
- Anastasilakis, A.D.; Makras, P.; Yavropoulou, M.P.; Tabacco, G.; Naciu, A.M.; Palermo, A. Denosumab Discontinuation and the Rebound Phenomenon: A Narrative Review. J. Clin. Med. 2021, 10, 152. [Google Scholar] [CrossRef]
- Bhattacharyya, N.; Hu, X.; Chen, C.Z.; Mathews Griner, L.A.; Zheng, W.; Inglese, J.; Austin, C.P.; Marugan, J.J.; Southall, N.; Neumann, S.; et al. A high throughput screening assay system for the identification of small molecule inhibitors of gsp. PLoS ONE 2014, 9, e90766. [Google Scholar] [CrossRef]
- Scanlan, M.J.; Raj, B.K.; Calvo, B.; Garin-Chesa, P.; Sanz-Moncasi, M.P.; Healey, J.H.; Old, L.J.; Rettig, W.J. Molecular cloning of fibroblast activation protein alpha, a member of the serine protease family selectively expressed in stromal fibroblasts of epithelial cancers. Proc. Natl. Acad. Sci. USA 1994, 91, 5657–5661. [Google Scholar] [CrossRef]
- Yazbeck, R.; Jaenisch, S.E.; Abbott, C.A. Potential disease biomarkers: Dipeptidyl peptidase 4 and fibroblast activation protein. Protoplasma 2018, 255, 375–386. [Google Scholar] [CrossRef]
- Hofheinz, R.D.; al-Batran, S.E.; Hartmann, F.; Hartung, G.; Jager, D.; Renner, C.; Tanswell, P.; Kunz, U.; Amelsberg, A.; Kuthan, H.; et al. Stromal antigen targeting by a humanised monoclonal antibody: An early phase II trial of sibrotuzumab in patients with metastatic colorectal cancer. Onkologie 2003, 26, 44–48. [Google Scholar] [CrossRef]
- Nemunaitis, J.; Vukelja, S.J.; Richards, D.; Cunningham, C.; Senzer, N.; Nugent, J.; Duncan, H.; Jones, B.; Haltom, E.; Uprichard, M.J. Phase I trial of PT-100 (PT-100), a cytokine-inducing small molecule, following chemotherapy for solid tumor malignancy. Cancer Investig. 2006, 24, 553–561. [Google Scholar] [CrossRef]
- Narra, K.; Mullins, S.R.; Lee, H.O.; Strzemkowski-Brun, B.; Magalong, K.; Christiansen, V.J.; McKee, P.A.; Egleston, B.; Cohen, S.J.; Weiner, L.M.; et al. Phase II trial of single agent Val-boroPro (Talabostat) inhibiting Fibroblast Activation Protein in patients with metastatic colorectal cancer. Cancer Biol. Ther. 2007, 6, 1691–1699. [Google Scholar] [CrossRef]
- Kratochwil, C.; Flechsig, P.; Lindner, T.; Abderrahim, L.; Altmann, A.; Mier, W.; Adeberg, S.; Rathke, H.; Rohrich, M.; Winter, H.; et al. (68)Ga-FAPI PET/CT: Tracer Uptake in 28 Different Kinds of Cancer. J. Nucl. Med. 2019, 60, 801–805. [Google Scholar] [CrossRef]
- Mu, X.; Mo, B.; Qin, J.; Li, Z.; Chong, W.; Zeng, Y.; Lu, L.; Zhang, L.; Fu, W. Comparative analysis of two timepoints on [(18)F]FAPI-42 PET/CT in various cancers. Eur. J. Hybrid Imaging 2023, 7, 27. [Google Scholar] [CrossRef]
- Luo, Y.; Zeng, Z.; Shan, T.; Xu, X.; Chen, J.; He, Y.; Zhang, T.; Huang, Z.; Chai, G.; Huang, Y.; et al. Fibroblast activation protein alpha activatable theranostic pro-photosensitizer for accurate tumor imaging and highly-specific photodynamic therapy. Theranostics 2022, 12, 3610–3627. [Google Scholar] [CrossRef]
- Brennen, W.N.; Rosen, D.M.; Chaux, A.; Netto, G.J.; Isaacs, J.T.; Denmeade, S.R. Pharmacokinetics and toxicology of a fibroblast activation protein (FAP)-activated prodrug in murine xenograft models of human cancer. Prostate 2014, 74, 1308–1319. [Google Scholar] [CrossRef]
- Chai, X.P.; Sun, G.L.; Fang, Y.F.; Hu, L.H.; Liu, X.; Zhang, X.W. Tumor-targeting efficacy of a BF211 prodrug through hydrolysis by fibroblast activation protein-alpha. Acta Pharmacol. Sin. 2018, 39, 415–424. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H.; Suen, C.S.; Lin, C.T.; Chien, C.H.; Lee, H.Y.; Chung, K.M.; Tsai, T.Y.; Jiaang, W.T.; Hwang, M.J.; Chen, X. Cleavage-site specificity of prolyl endopeptidase FAP investigated with a full-length protein substrate. J. Biochem. 2011, 149, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Michel, Z.; Raborn, L.N.; Spencer, T.; Pan, K.S.; Martin, D.; Roszko, K.L.; Wang, Y.; Robey, P.G.; Collins, M.T.; Boyce, A.M.; et al. Transcriptomic Signature and Pro-Osteoclastic Secreted Factors of Abnormal Bone-Marrow Stromal Cells in Fibrous Dysplasia. Cells 2024, 13, 774. [Google Scholar] [CrossRef]
- Kim, H.Y.; Charton, C.; Shim, J.H.; Lim, S.Y.; Kim, J.; Lee, S.; Ohn, J.H.; Kim, B.K.; Heo, C.Y. Patient-Derived Organoids Recapitulate Pathological Intrinsic and Phenotypic Features of Fibrous Dysplasia. Cells 2024, 13, 729. [Google Scholar] [CrossRef] [PubMed]
- Persichetti, A.; Milanetti, E.; Palmisano, B.; di Filippo, A.; Spica, E.; Donsante, S.; Coletta, I.; Venti, M.D.S.; Ippolito, E.; Corsi, A.; et al. Nanostring technology on Fibrous Dysplasia bone biopsies. A pilot study suggesting different histology-related molecular profiles. Bone Rep. 2022, 16, 101156. [Google Scholar] [CrossRef] [PubMed]
- Raimondo, D.; Remoli, C.; Astrologo, L.; Burla, R.; La Torre, M.; Verni, F.; Tagliafico, E.; Corsi, A.; Del Giudice, S.; Persichetti, A.; et al. Changes in gene expression in human skeletal stem cells transduced with constitutively active Gsalpha correlates with hallmark histopathological changes seen in fibrous dysplastic bone. PLoS ONE 2020, 15, e0227279. [Google Scholar] [CrossRef]
- Onodera, S.; Saito, A.; Hojo, H.; Nakamura, T.; Zujur, D.; Watanabe, K.; Morita, N.; Hasegawa, D.; Masaki, H.; Nakauchi, H.; et al. Hedgehog Activation Regulates Human Osteoblastogenesis. Stem Cell Rep. 2020, 15, 125–139. [Google Scholar] [CrossRef]
- Zhou, S.H.; Yang, W.J.; Liu, S.W.; Li, J.; Zhang, C.Y.; Zhu, Y.; Zhang, C.P. Gene expression profiling of craniofacial fibrous dysplasia reveals ADAMTS2 overexpression as a potential marker. Int. J. Clin. Exp. Pathol. 2014, 7, 8532–8541. [Google Scholar]
- Piersanti, S.; Remoli, C.; Saggio, I.; Funari, A.; Michienzi, S.; Sacchetti, B.; Robey, P.G.; Riminucci, M.; Bianco, P. Transfer, analysis, and reversion of the fibrous dysplasia cellular phenotype in human skeletal progenitors. J. Bone Miner. Res. 2010, 25, 1103–1116. [Google Scholar] [CrossRef] [PubMed]
- Kiss, J.; Balla, B.; Kosa, J.P.; Borsy, A.; Podani, J.; Takacs, I.; Lazary, A.; Nagy, Z.; Bacsi, K.; Kis, A.; et al. Gene expression patterns in the bone tissue of women with fibrous dysplasia. Am. J. Med. Genet. A 2010, 152A, 2211–2220. [Google Scholar] [CrossRef]
- Lee, C.H.; Espinosa, I.; Jensen, K.C.; Subramanian, S.; Zhu, S.X.; Varma, S.; Montgomery, K.D.; Nielsen, T.O.; van de Rijn, M.; West, R.B. Gene expression profiling identifies p63 as a diagnostic marker for giant cell tumor of the bone. Mod. Pathol. 2008, 21, 531–539. [Google Scholar] [CrossRef]
- Papadakis, G.Z.; Manikis, G.C.; Karantanas, A.H.; Florenzano, P.; Bagci, U.; Marias, K.; Collins, M.T.; Boyce, A.M. (18) F-NaF PET/CT IMAGING IN FIBROUS DYSPLASIA OF BONE. J. Bone Miner. Res. 2019, 34, 1619–1631. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Tian, Z.; Zhang, C.; He, Y. Monosotic fibrous dysplasia and solitary intramuscular myxoma of the head and neck: A unique presentation of Mazabraud’s syndrome and a literature review. Oncol. Lett. 2015, 10, 3087–3094. [Google Scholar] [CrossRef]
- Mu, X.; Li, Z.; Qin, J.; Wang, Z.; Fu, W. Comparison of 18 F-FAPI and 18 F-FDG PET/CT in a Patient with Fibrous Dysplasia. Clin. Nucl. Med. 2024, 49, e182–e183. [Google Scholar] [CrossRef] [PubMed]
- Poreba, M. Protease-activated prodrugs: Strategies, challenges, and future directions. FEBS J. 2020, 287, 1936–1969. [Google Scholar] [CrossRef]
- Lu, Y.C.; Chuang, C.H.; Chuang, K.H.; Chen, I.J.; Huang, B.C.; Lee, W.H.; Wang, H.E.; Li, J.J.; Cheng, Y.A.; Cheng, K.W.; et al. Specific activation of pro-Infliximab enhances selectivity and safety of rheumatoid arthritis therapy. PLoS Biol. 2019, 17, e3000286. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Study [Ref] | Sample Type | GEO Accession | mRNA Expression Technique | FAP Expression Measured? | FAP Upregulation |
|---|---|---|---|---|---|
| Kim, HY, 2024 [17] | Single cells from FD tissue | GSE263294 | scRNAseq | Yes (scRNAseq) | N/A, no control group |
| Kim, HY, 2024 [17] | FD BSMC vs. normal BMSC-derived organoids (culture) | Not submitted | qPCR | No | N/A |
| Michel Z, 2024 [16] | Human FD and HD BMSCs | GSE261360 | RNAseq | Yes | Yes, FD vs. HD BMSCs |
| Michel Z, 2024 [16] | Mouse BMSCs induced or not to express GαsR201C | GSE261360 | RNAseq | Yes | No |
| De Castro LF, 2023 [1] | Human FD tissue before and after denosumab treatment | GSE250357 | RNAseq | Yes | Yes, FD vs. denosumab-treated FD |
| De Castro LF, 2023 [1] | Mouse FD tissue with or without anti-RANKL treatment and same site control bone | GSE250357 | RNAseq | Yes | Yes, FD vs. anti-RANKL-treated FD and vs. control bone |
| Persichetti, 2021 [18] | FD biopsies | GSE176243 | Nanostring | No | N/A |
| Raimondo D, 2020 [19] | Human BMSC expressing GαsR201C by lentiviral transduction | GSE109818 | Microarray | Yes | No |
| Onodera S, 2020 [20] | GNAS p.R201H iPSC vs. WT iPSC | Not submitted | qPCR | No | N/A |
| Zhou S-H, 2014 [21] | Human FD tissue vs. normal bone | Not submitted | Microarray | No | N/A |
| Piersanti S, 2010 [22] | Human BMSC expressing GαsR201C by lentiviral transduction | Not submitted | qPCR | No | N/A |
| Kiss J, 2010 [23] | FD affected women’s tissue vs. control women’s bone | Not submitted | qPCR | No | N/A |
| Lee C-H, 2008 [24] | FD biopsies vs. GCT and ABC biopsies | Not submitted | Microarray | Unk * | Unk |
| Fibrous Dysplasia (FD) Patients | Healthy Donors (HDs) | |
|---|---|---|
| Subjects—n | 47 | 22 |
| Females—n (%) | 30 (63.8%) | 8 (36.4%) |
| Age—mean ± SD (range) | 33 ± 13 (18–71) | 42 ± 15 (18–70) |
| FD burden—mean ± SD (range) | 37 ± 23 (2.8–75) | 0 |
| Currently on bisphosphonates—n (%) | 10 (21%) | 0 |
| Any endocrinopathy—n (%) | 35 (74%) | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raborn, L.N.; Michel, Z.; Collins, M.T.; Boyce, A.M.; de Castro, L.F. Fibroblast Activation Protein Is Expressed by Altered Osteoprogenitors and Associated to Disease Burden in Fibrous Dysplasia. Cells 2024, 13, 1434. https://doi.org/10.3390/cells13171434
Raborn LN, Michel Z, Collins MT, Boyce AM, de Castro LF. Fibroblast Activation Protein Is Expressed by Altered Osteoprogenitors and Associated to Disease Burden in Fibrous Dysplasia. Cells. 2024; 13(17):1434. https://doi.org/10.3390/cells13171434
Chicago/Turabian StyleRaborn, Layne N., Zachary Michel, Michael T. Collins, Alison M. Boyce, and Luis F. de Castro. 2024. "Fibroblast Activation Protein Is Expressed by Altered Osteoprogenitors and Associated to Disease Burden in Fibrous Dysplasia" Cells 13, no. 17: 1434. https://doi.org/10.3390/cells13171434