
Enhanced Performance of the Optimized Dye CF583R in Direct Stochastic Optical Reconstruction Microscopy of Active Zones in Drosophila Melanogaster

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fly Husbandry
2.2. Dissection, Fixation and Staining
2.3. dSTORM (Direct Stochastic Optical Reconstruction Micrsocopy)
2.4. Data Analysis
2.5. Statistics
3. Results
3.1. Application of AF647, CF568 and CF583R Permits dSTORM Imaging of AZs
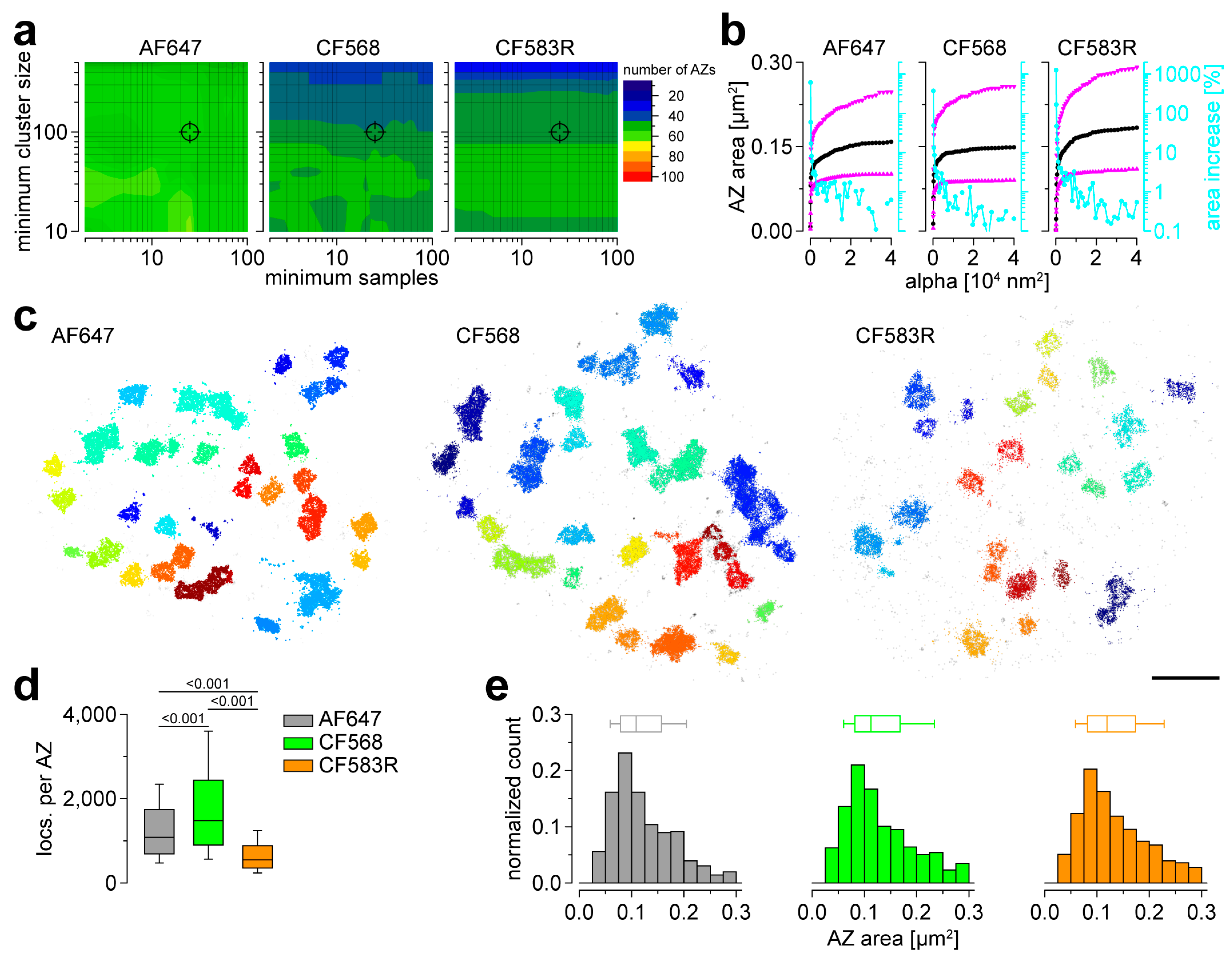
3.2. AF647, CF568 and CF583R Perform Equally in Basal AZ Morphometry
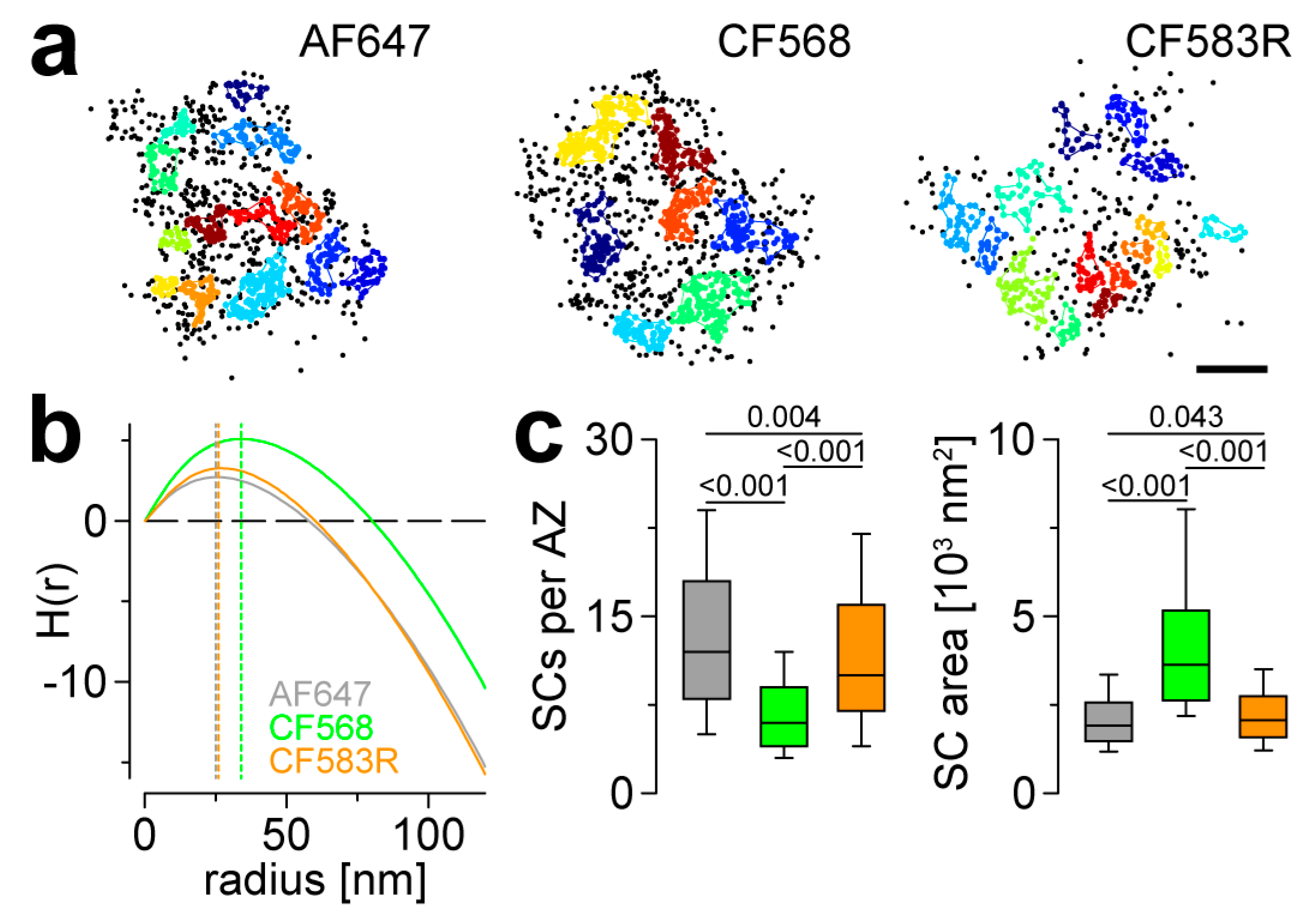
3.3. CF583R Outperforms CF568 and Matches AF647 in AZ Nanocluster Analyses
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Werner, C.; Sauer, M.; Geis, C. Super-resolving Microscopy in Neuroscience. Chem. Rev. 2021, 121, 11971–12015. [Google Scholar] [CrossRef]
- Lelek, M.; Gyparaki, M.T.; Beliu, G.; Schueder, F.; Griffié, J.; Manley, S.; Jungmann, R.; Sauer, M.; Lakadamyali, M.; Zimmer, C. Single-molecule localization microscopy. Nat. Rev. Methods Primers 2021, 1, 39. [Google Scholar] [CrossRef] [PubMed]
- Heilemann, M.; van de Linde, S.; Schüttpelz, M.; Kasper, R.; Seefeldt, B.; Mukherjee, A.; Tinnefeld, P.; Sauer, M. Subdiffraction-resolution fluorescence imaging with conventional fluorescent probes. Angew. Chem. Int. Ed. Engl. 2008, 47, 6172–6176. [Google Scholar] [CrossRef] [PubMed]
- van de Linde, S.; Löschberger, A.; Klein, T.; Heidbreder, M.; Wolter, S.; Heilemann, M.; Sauer, M. Direct stochastic optical reconstruction microscopy with standard fluorescent probes. Nat. Protoc. 2011, 6, 991–1009. [Google Scholar] [CrossRef]
- Dempsey, G.T.; Vaughan, J.C.; Chen, K.H.; Bates, M.; Zhuang, X. Evaluation of fluorophores for optimal performance in localization-based super-resolution imaging. Nat. Methods 2011, 8, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Xiong, M.; Susanto, J.; Li, X.; Leung, W.Y.; Xu, K. Transforming Rhodamine Dyes for (d)STORM Super-Resolution Microscopy via 1,3-Disubstituted Imidazolium Substitution. Angew. Chem. Int. Ed. Engl. 2022, 61, e202113612. [Google Scholar] [CrossRef]
- Ehmann, N.; van de Linde, S.; Alon, A.; Ljaschenko, D.; Keung, X.Z.; Holm, T.; Rings, A.; DiAntonio, A.; Hallermann, S.; Ashery, U.; et al. Quantitative super-resolution imaging of Bruchpilot distinguishes active zone states. Nat. Commun. 2014, 5, 4650. [Google Scholar] [CrossRef] [PubMed]
- Mrestani, A.; Pauli, M.; Kollmannsberger, P.; Repp, F.; Kittel, R.J.; Eilers, J.; Doose, S.; Sauer, M.; Sirén, A.L.; Heckmann, M.; et al. Active zone compaction correlates with presynaptic homeostatic potentiation. Cell Rep. 2021, 37, 109770. [Google Scholar] [CrossRef] [PubMed]
- Held, R.G.; Kaeser, P.S. ELKS active zone proteins as multitasking scaffolds for secretion. Open Biol. 2018, 8, 170258. [Google Scholar] [CrossRef] [PubMed]
- Kittel, R.J.; Wichmann, C.; Rasse, T.M.; Fouquet, W.; Schmidt, M.; Schmid, A.; Wagh, D.A.; Pawlu, C.; Kellner, R.R.; Willig, K.I.; et al. Bruchpilot promotes active zone assembly, Ca2+ channel clustering, and vesicle release. Science 2006, 312, 1051–1054. [Google Scholar] [CrossRef] [PubMed]
- Wagh, D.A.; Rasse, T.M.; Asan, E.; Hofbauer, A.; Schwenkert, I.; Dürrbeck, H.; Buchner, S.; Dabauvalle, M.C.; Schmidt, M.; Qin, G.; et al. Bruchpilot, a protein with homology to ELKS/CAST, is required for structural integrity and function of synaptic active zones in Drosophila. Neuron 2006, 49, 833–844. [Google Scholar] [CrossRef]
- Stewart, B.A.; Atwood, H.L.; Renger, J.J.; Wang, J.; Wu, C.F. Improved stability of Drosophila larval neuromuscular preparations in haemolymph-like physiological solutions. J. Comp. Physiol. A. 1994, 175, 179–191. [Google Scholar] [CrossRef]
- Tokunaga, M.; Imamoto, N.; Sakata-Sogawa, K. Highly inclined thin illumination enables clear single-molecule imaging in cells. Nat. Methods 2008, 5, 159–161. [Google Scholar] [CrossRef]
- Wolter, S.; Löschberger, A.; Holm, T.; Aufmkolk, S.; Dabauvalle, M.C.; van de Linde, S.; Sauer, M. rapidSTORM: Accurate, fast open-source software for localization microscopy. Nat. Methods 2012, 9, 1040–1041. [Google Scholar] [CrossRef] [PubMed]
- Endesfelder, U.; Malkusch, S.; Fricke, F.; Heilemann, M. A simple method to estimate the average localization precision of a single-molecule localization microscopy experiment. Histochem. Cell Biol. 2014, 141, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Malkusch, S.; Heilemann, M. Extracting quantitative information from single-molecule super-resolution imaging data with LAMA-LocAlization Microscopy Analyzer. Sci. Rep. 2016, 6, 34486. [Google Scholar] [CrossRef] [PubMed]
- Dannhäuser, S.; Mrestani, A.; Gundelach, F.; Pauli, M.; Komma, F.; Kollmannsberger, P.; Sauer, M.; Heckmann, M.; Paul, M.M. Endogenous tagging of Unc-13 reveals nanoscale reorganization at active zones during presynaptic homeostatic potentiation. Front. Cell. Neurosci. 2022, 16, 1074304. [Google Scholar] [CrossRef]
- Mrestani, A.; Dannhäuser, S.; Pauli, M.; Kollmannsberger, P.; Hübsch, M.; Morris, L.; Langenhan, T.; Heckmann, M.; Paul, M.M. Nanoscaled RIM clustering at presynaptic active zones revealed by endogenous tagging. Life Sci. Alliance 2023, 6, e202302021. [Google Scholar] [CrossRef] [PubMed]
- Beckers, C.J.; Mrestani, A.; Komma, F.; Dannhäuser, S. Versatile Endogenous Editing of GluRIIA in Drosophila melanogaster. Cells 2024, 13, 323. [Google Scholar] [CrossRef]
- Kluyver, T.; Ragan-Kelley, B.; Pérez, F.; Granger, B.; Bussonnier, M.; Frederic, J.; Kelley, K.; Hamrick, J.; Grout, J.; Corlay, S.; et al. Jupyter Notebooks—A publishing format for reproducible computational workflows. In Positioning and Power in Academic Publishing: Players, Agents and Agendas; Loizides, F., Schmidt, B., Eds.; IOS Press: Amsterdam, The Netherlands, 2016; pp. 87–90. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- McInnes, L.; Healy, J.; Astels, S. Hdbscan: Hierarchical Density Based Clustering. J. Open Source Softw. 2017, 2, 205. [Google Scholar] [CrossRef]
- Robitaille, T.P.; Tollerud, E.J.; Greenfield, P.; Droettboom, M.; Bray, E.; Aldcroft, T.; Davis, M.; Ginsburg, A.; Price-Whelan, A.M.; Kerzendorf, W.E.; et al. Astropy: A community Python package for astronomy. Astron. Astrophys. 2013, 558, A33. [Google Scholar] [CrossRef]
- Kiskowski, M.A.; Hancock, J.F.; Kenworthy, A.K. On the use of Ripley’s K-function and its derivatives to analyze domain size. Biophys. J. 2009, 97, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Glebov, O.O.; Jackson, R.E.; Winterflood, C.M.; Owen, D.M.; Barker, E.A.; Doherty, P.; Ewers, H.; Burrone, J. Nanoscale Structural Plasticity of the Active Zone Matrix Modulates Presynaptic Function. Cell Rep. 2017, 18, 2715–2728. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, H.; Ariyoshi, T.; Kimpara, N.; Sugao, K.; Taiko, I.; Takikawa, K.; Asanuma, D.; Namiki, S.; Hirose, K. Synaptic weight set by Munc13-1 supramolecular assemblies. Nat. Neurosci. 2018, 21, 41–49. [Google Scholar] [CrossRef]
- Tang, A.H.; Chen, H.; Li, T.P.; Metzbower, S.R.; MacGillavry, H.D.; Blanpied, T.A. A trans-synaptic nanocolumn aligns neurotransmitter release to receptors. Nature 2016, 536, 210–214. [Google Scholar] [CrossRef]
- Siddig, S.; Aufmkolk, S.; Doose, S.; Jobin, M.L.; Werner, C.; Sauer, M.; Calebiro, D. Super-resolution imaging reveals the nanoscale organization of metabotropic glutamate receptors at presynaptic active zones. Sci. Adv. 2020, 6, eaay7193. [Google Scholar] [CrossRef]
- Newman, Z.L.; Bakshinskaya, D.; Schultz, R.; Kenny, S.J.; Moon, S.; Aghi, K.; Stanley, C.; Marnani, N.; Li, R.; Bleier, J.; et al. Determinants of synapse diversity revealed by super-resolution quantal transmission and active zone imaging. Nat. Commun. 2022, 13, 229. [Google Scholar] [CrossRef]
- Ghelani, T.; Escher, M.; Thomas, U.; Esch, K.; Lützkendorf, J.; Depner, H.; Maglione, M.; Parutto, P.; Gratz, S.; Matkovic-Rachid, T.; et al. Interactive nanocluster compaction of the ELKS scaffold and Cacophony Ca2+ channels drives sustained active zone potentiation. Sci. Adv. 2023, 9, eade7804. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noß, M.; Ljaschenko, D.; Mrestani, A. Enhanced Performance of the Optimized Dye CF583R in Direct Stochastic Optical Reconstruction Microscopy of Active Zones in Drosophila Melanogaster. Cells 2024, 13, 1445. https://doi.org/10.3390/cells13171445
Noß M, Ljaschenko D, Mrestani A. Enhanced Performance of the Optimized Dye CF583R in Direct Stochastic Optical Reconstruction Microscopy of Active Zones in Drosophila Melanogaster. Cells. 2024; 13(17):1445. https://doi.org/10.3390/cells13171445
Chicago/Turabian StyleNoß, Marvin, Dmitrij Ljaschenko, and Achmed Mrestani. 2024. "Enhanced Performance of the Optimized Dye CF583R in Direct Stochastic Optical Reconstruction Microscopy of Active Zones in Drosophila Melanogaster" Cells 13, no. 17: 1445. https://doi.org/10.3390/cells13171445
APA StyleNoß, M., Ljaschenko, D., & Mrestani, A. (2024). Enhanced Performance of the Optimized Dye CF583R in Direct Stochastic Optical Reconstruction Microscopy of Active Zones in Drosophila Melanogaster. Cells, 13(17), 1445. https://doi.org/10.3390/cells13171445