
LIP1 Regulates the Plant Circadian Oscillator by Modulating the Function of the Clock Component GIGANTEA

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Analysis of Gene Expression
2.3. Yeast Two-Hybrid Screening and Testing Pairwise Protein Interactions
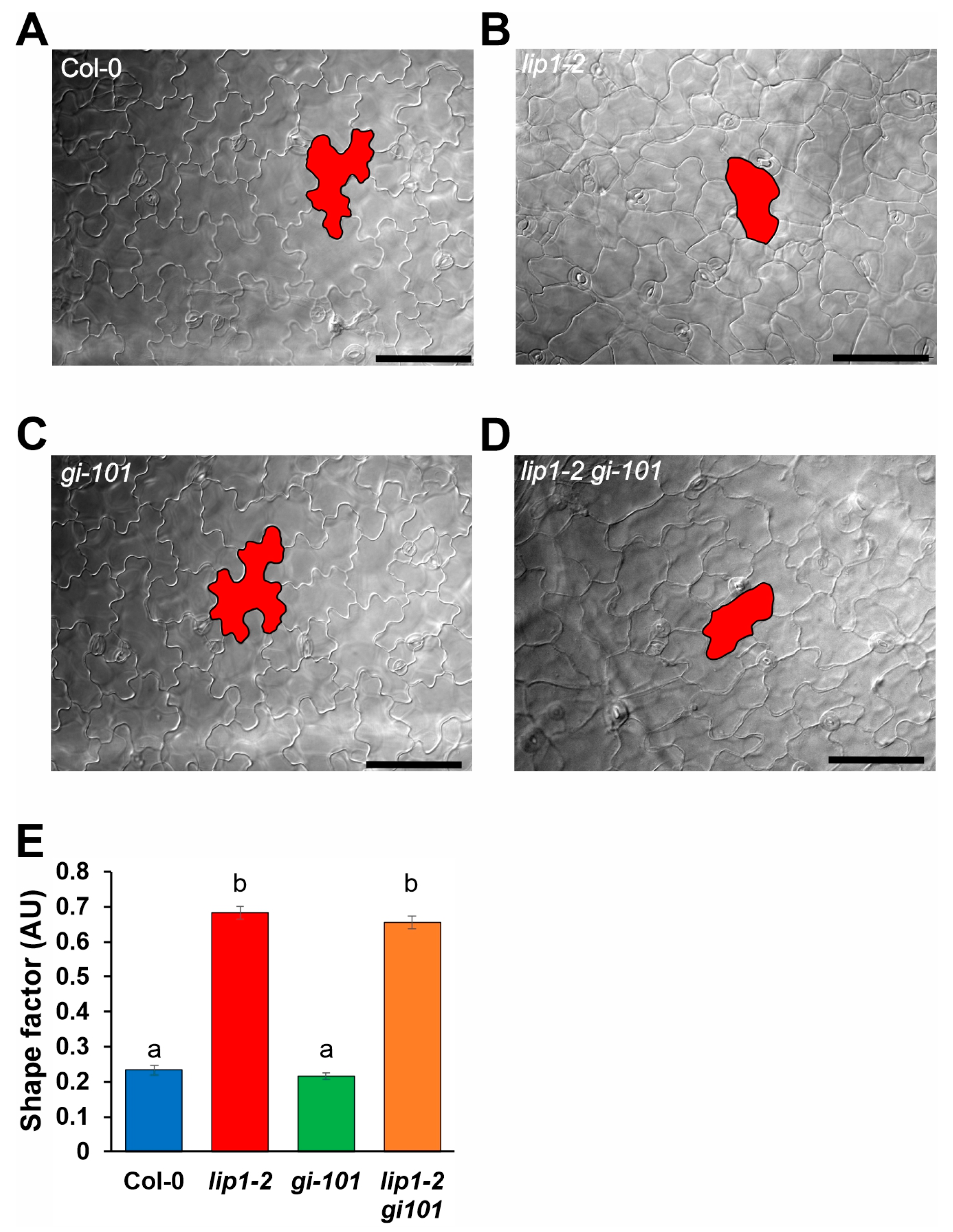
2.4. Analysis of Pavement Cell Morphology
2.5. Physiological Assays
2.6. Statistical Analysis
3. Results
3.1. Pattern and Level of Clock Gene Expression in the lip1-2 Mutant
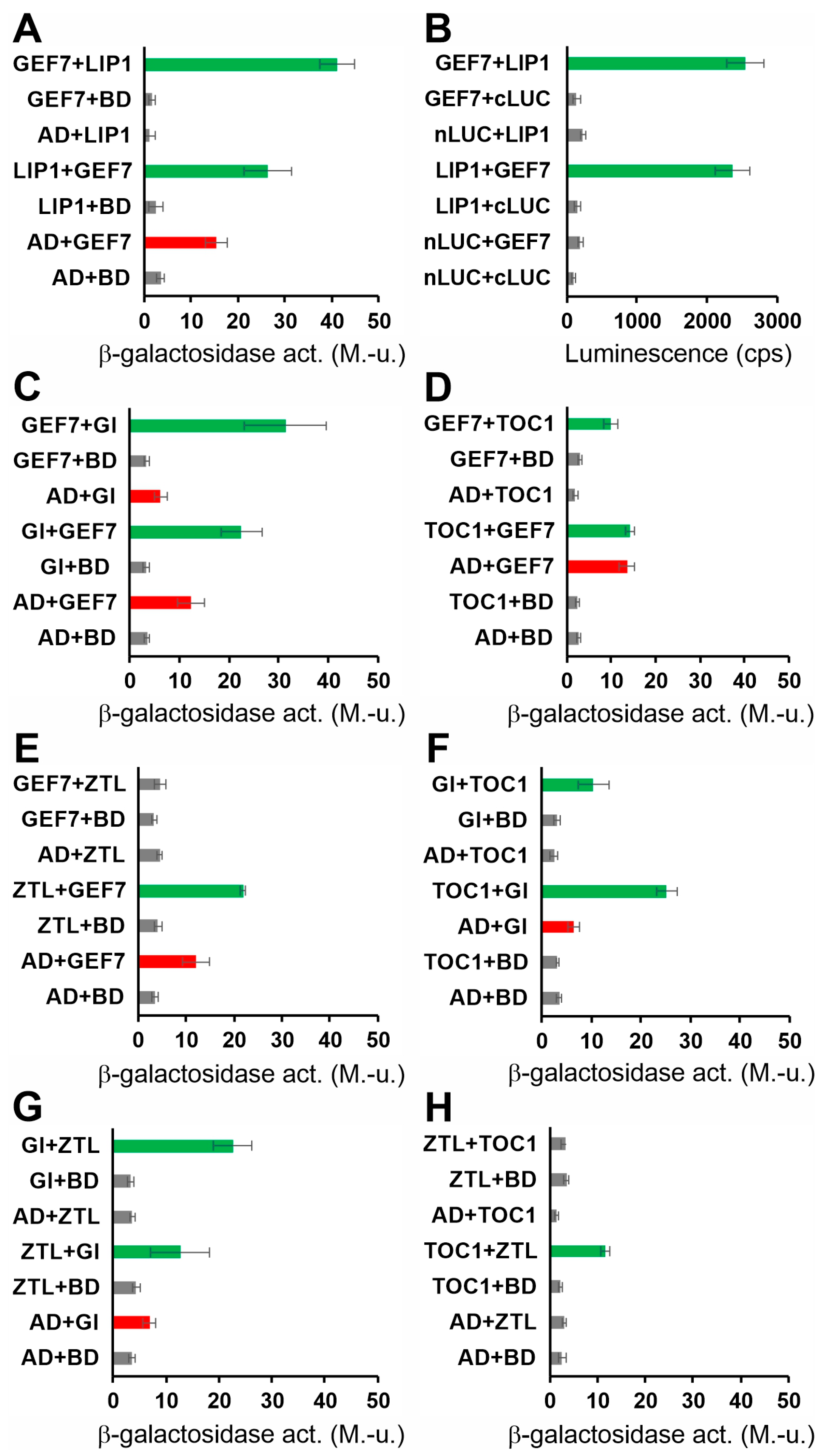
3.2. Identification of Proteins Interacting with LIP1
3.3. Genetic Analysis Identifies GI as the Clock Component Targeted by LIP1
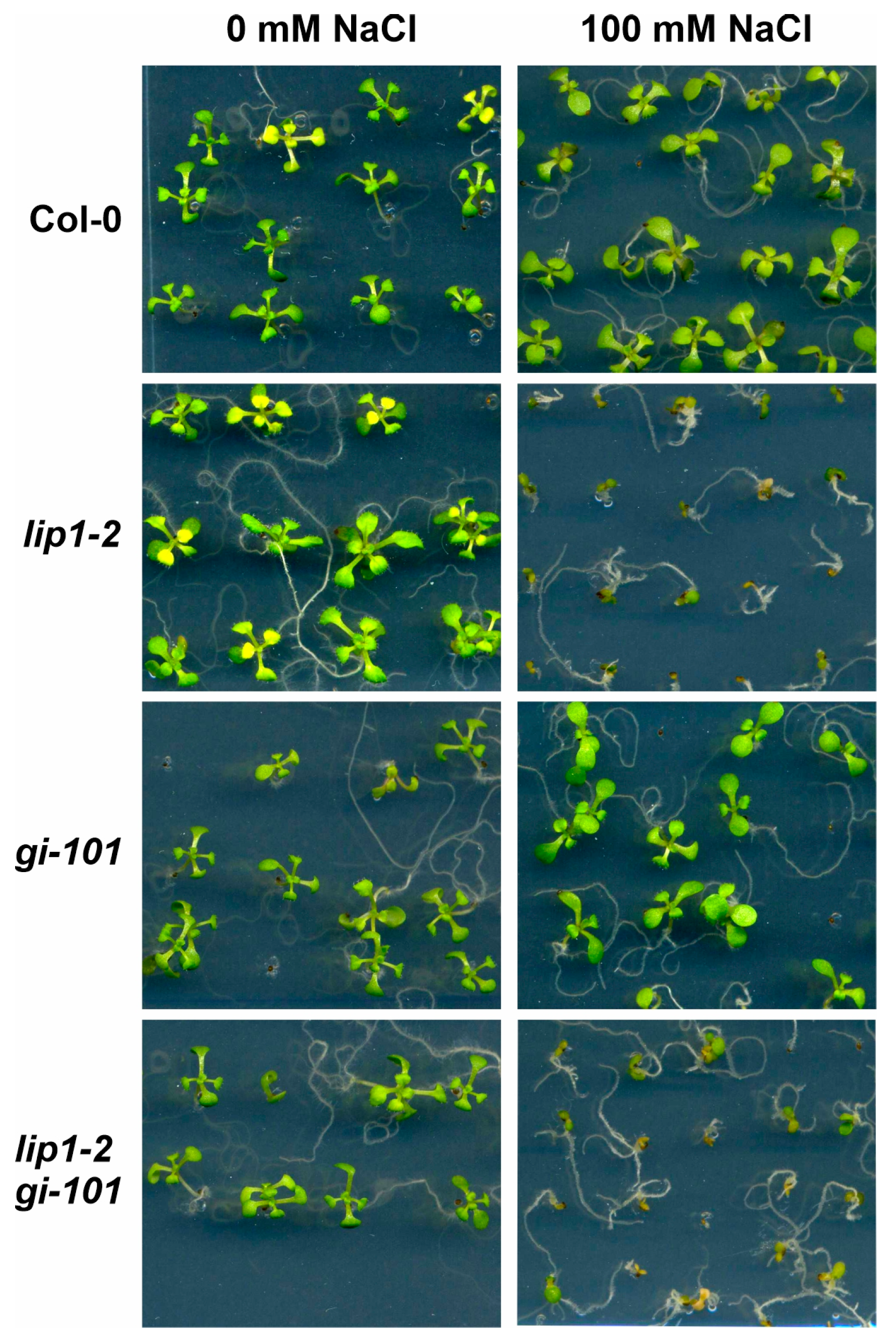
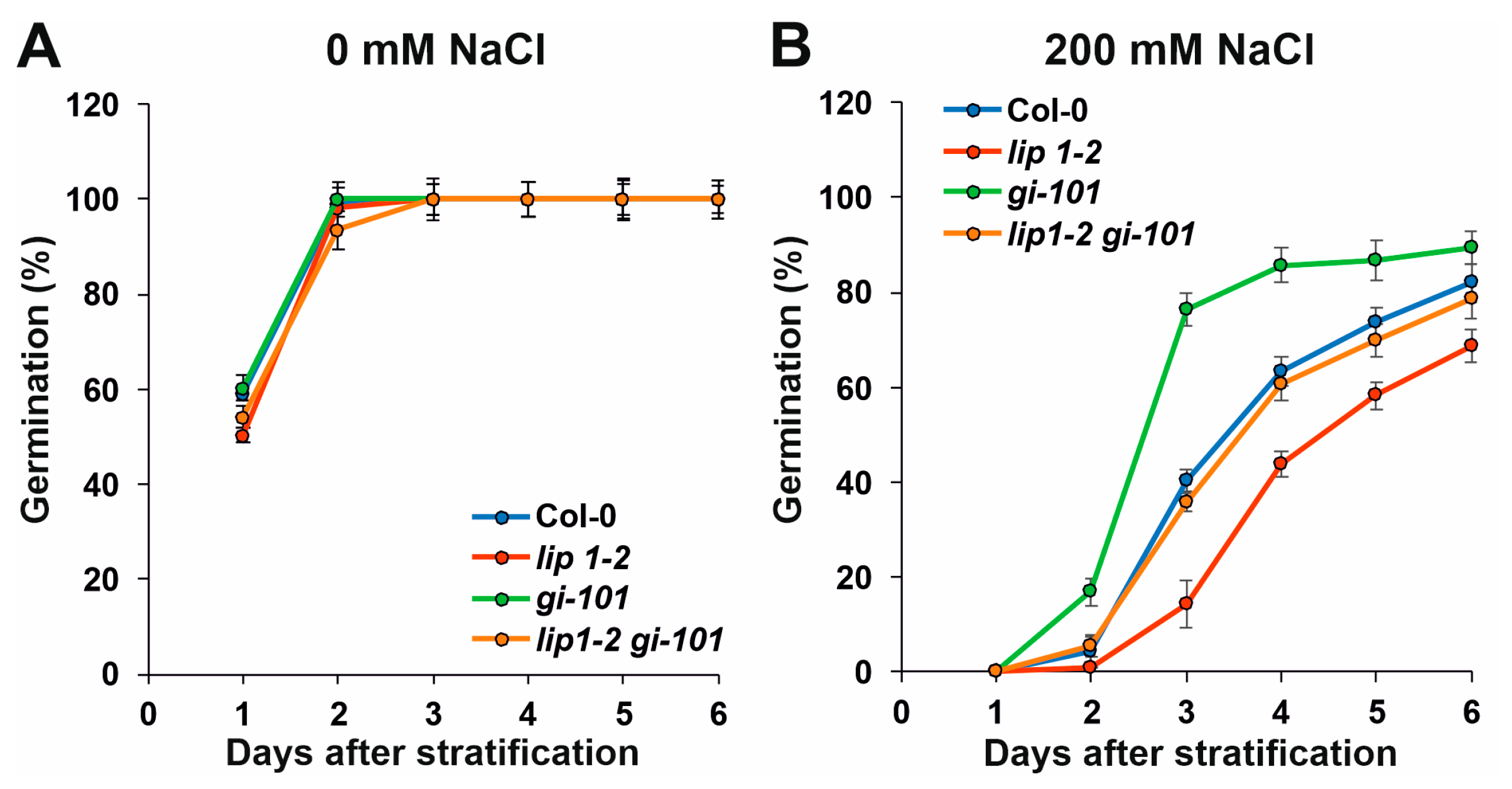
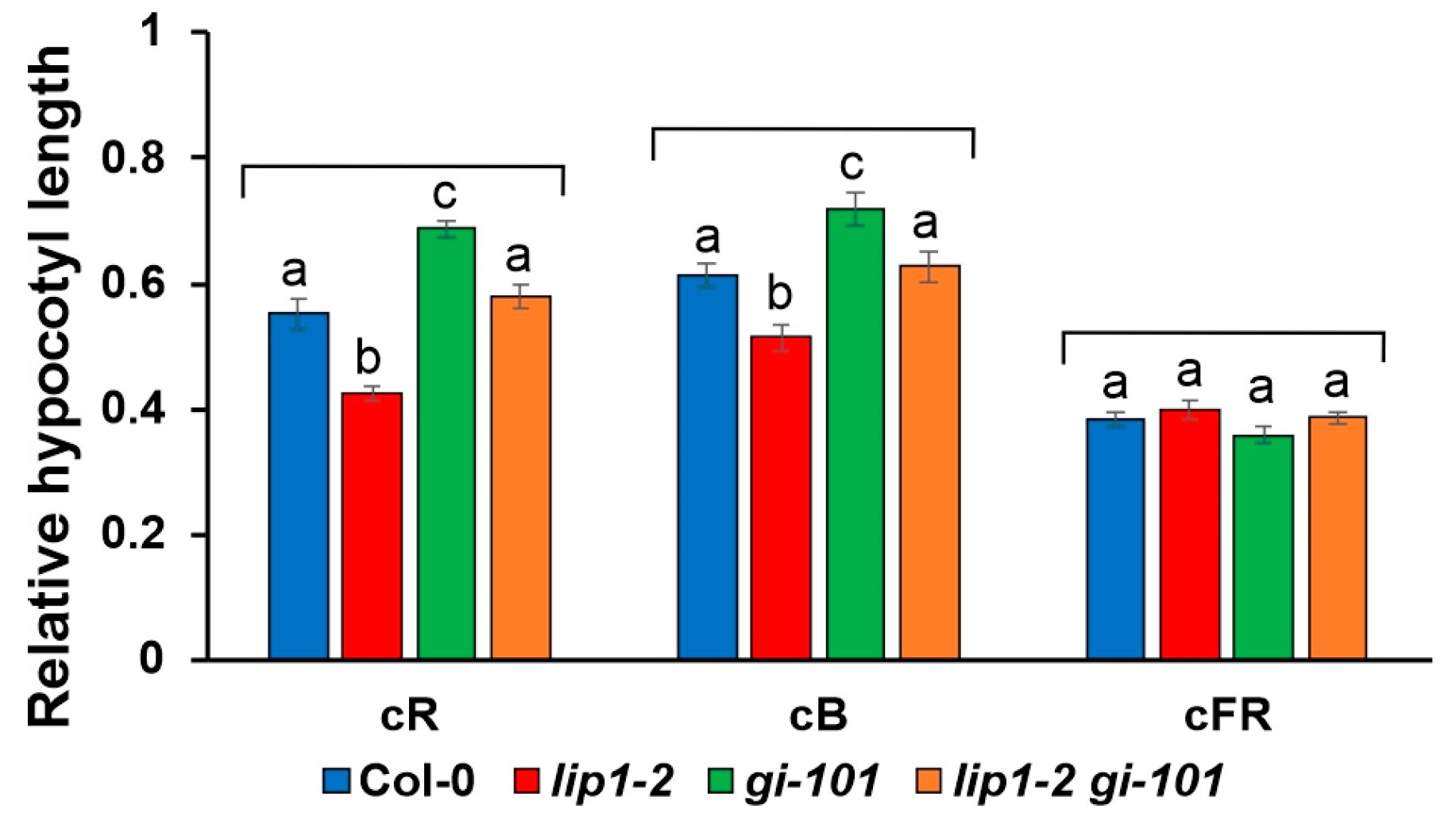
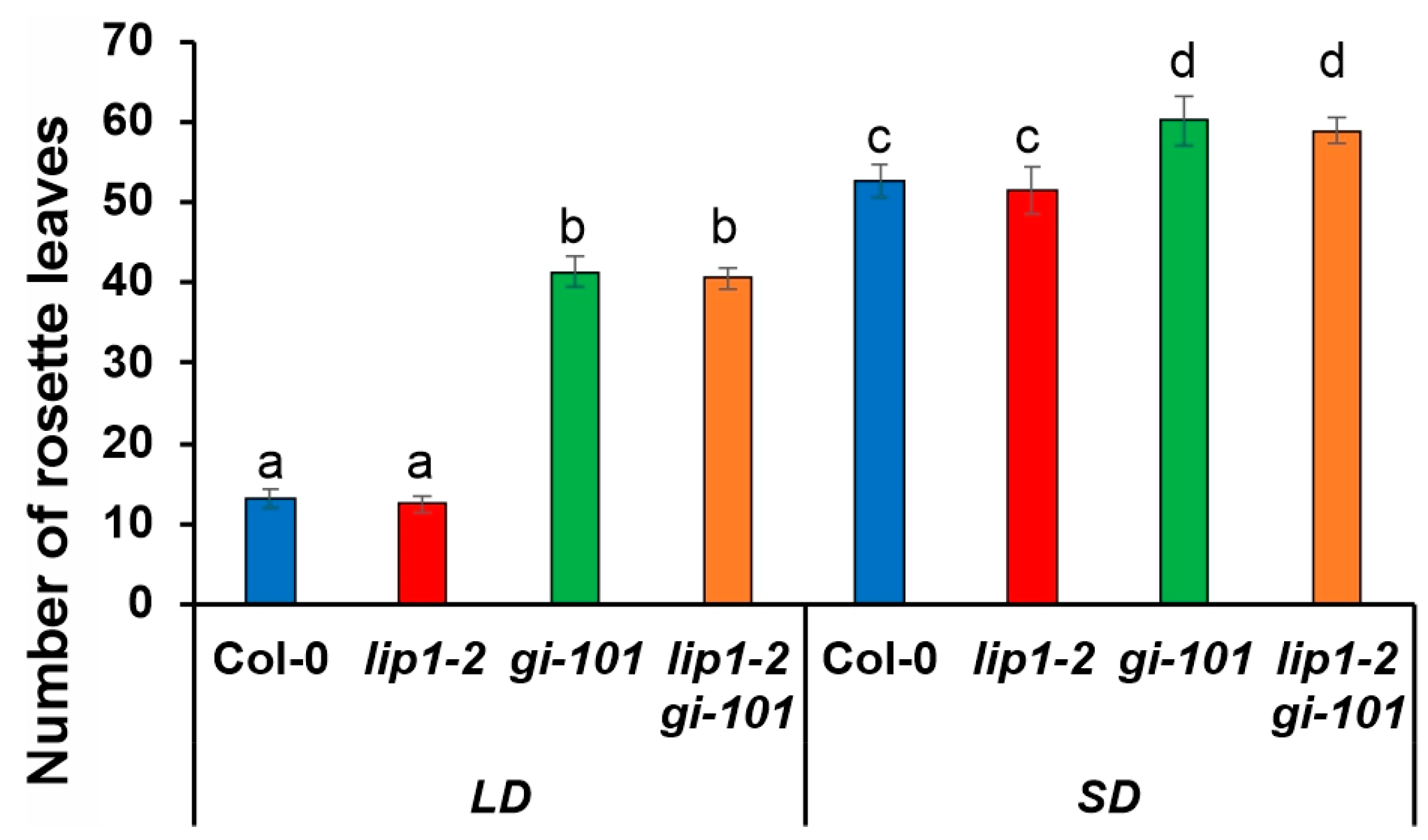
3.4. Additive Effects of LIP1 and GI on Salt Stress Responses and Photomorphogenesis
3.5. Regulation of Cell Morphology and Control of Flowering Time Are Not Shared Functions of LIP1 and GI
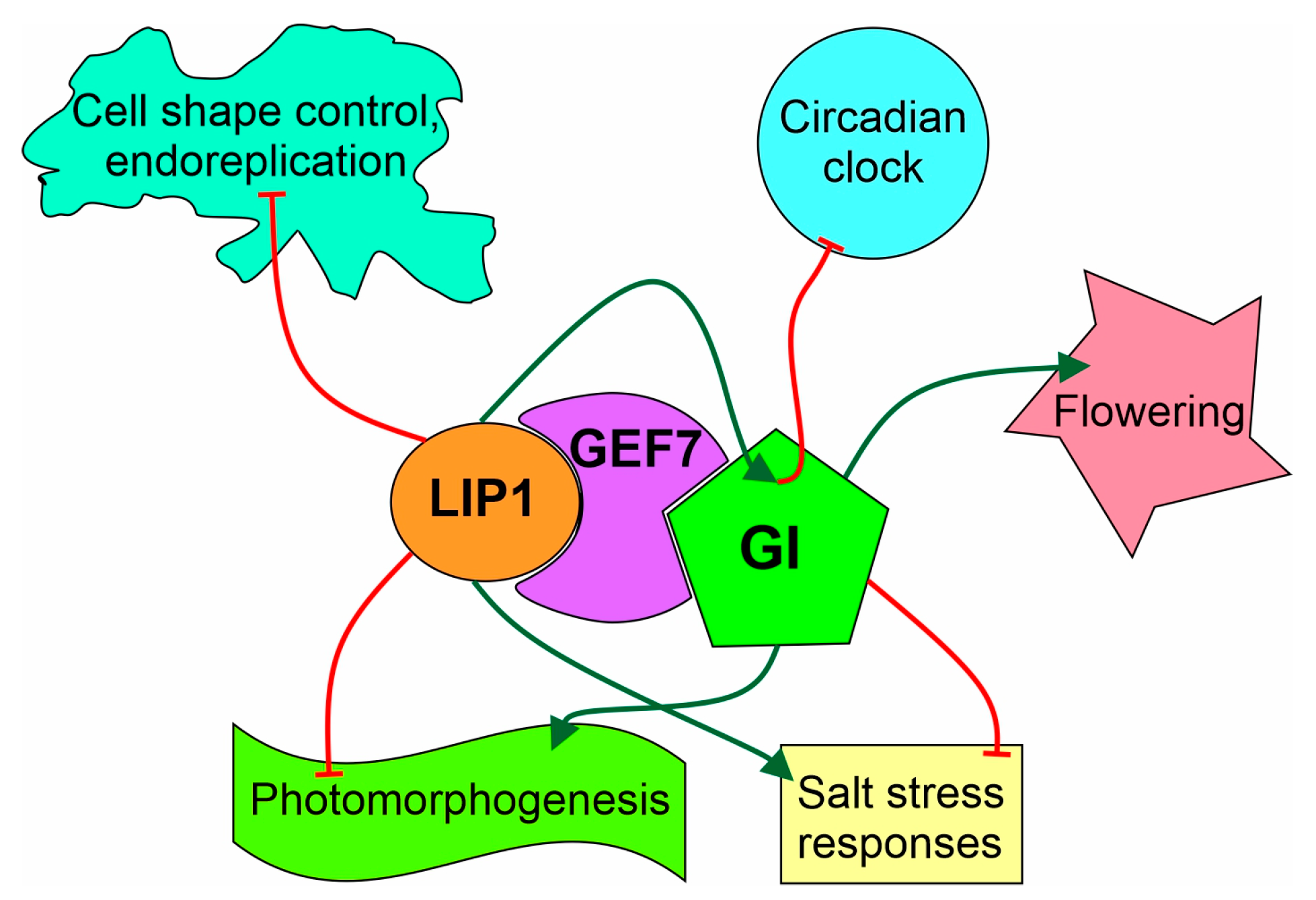
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harmer, S.L.; Panda, S.; Kay, S.A. Molecular bases of circadian rhythms. Annu. Rev. Cell Dev. Biol. 2001, 17, 215–253. [Google Scholar] [CrossRef]
- Patke, A.; Young, M.W.; Axelrod, S. Molecular mechanisms and physiological importance of circadian rhythms. Nat. Rev. Mol. Cell Biol. 2020, 21, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, S.E.; Rugnone, M.L.; Kay, S.A. Light Perception: A Matter of Time. Mol. Plant 2020, 13, 363–385. [Google Scholar] [CrossRef] [PubMed]
- Gliech, C.R.; Holland, A.J. Keeping track of time: The fundamentals of cellular clocks. J. Cell Biol. 2020, 219, e202005136. [Google Scholar] [CrossRef] [PubMed]
- Green, R.M.; Tobin, E.M. The role of CCA1 and LHY in the plant circadian clock. Dev. Cell 2002, 2, 516–518. [Google Scholar] [CrossRef] [PubMed]
- Strayer, C.; Oyama, T.; Schultz, T.F.; Raman, R.; Somers, D.E.; Mas, P.; Panda, S.; Kreps, J.A.; Kay, S.A. Cloning of the Arabidopsis clock gene TOC1, an autoregulatory response regulator homolog. Science 2000, 289, 768–771. [Google Scholar] [CrossRef]
- Huang, W.; Perez-Garcia, P.; Pokhilko, A.; Millar, A.J.; Antoshechkin, I.; Riechmann, J.L.; Mas, P. Mapping the core of the Arabidopsis circadian clock defines the network structure of the oscillator. Science 2012, 336, 75–79. [Google Scholar] [CrossRef]
- Hotta, C.T. The evolution and function of the PSEUDO RESPONSE REGULATOR gene family in the plant circadian clock. Genet. Mol. Biol. 2022, 45, e20220137. [Google Scholar] [CrossRef]
- Nakamichi, N.; Kiba, T.; Henriques, R.; Mizuno, T.; Chua, N.H.; Sakakibara, H. PSEUDO-RESPONSE REGULATORS 9, 7, and 5 are transcriptional repressors in the Arabidopsis circadian clock. Plant Cell 2010, 22, 594–605. [Google Scholar] [CrossRef]
- Huang, H.; Nusinow, D.A. Into the Evening: Complex Interactions in the Arabidopsis Circadian Clock. Trends Genet. 2016, 32, 674–686. [Google Scholar] [CrossRef]
- Wu, J.F.; Tsai, H.L.; Joanito, I.; Wu, Y.C.; Chang, C.W.; Li, Y.H.; Wang, Y.; Hong, J.C.; Chu, J.W.; Hsu, C.P.; et al. LWD-TCP complex activates the morning gene CCA1 in Arabidopsis. Nat. Commun. 2016, 7, 13181. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Gil, S.; Grasser, K.D.; Mas, P. Targeted Recruitment of the Basal Transcriptional Machinery by LNK Clock Components Controls the Circadian Rhythms of Nascent RNAs in Arabidopsis. Plant Cell 2018, 30, 907–924. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Li, P.; Li, M.; Li, W.; Yao, T.; Wu, J.W.; Gu, W.; Cohen, R.E.; Shi, Y. Crystal structure of a UBP-family deubiquitinating enzyme in isolation and in complex with ubiquitin aldehyde. Cell 2002, 111, 1041–1054. [Google Scholar] [CrossRef] [PubMed]
- Kiba, T.; Henriques, R.; Sakakibara, H.; Chua, N.H. Targeted degradation of PSEUDO-RESPONSE REGULATOR5 by an SCFZTL complex regulates clock function and photomorphogenesis in Arabidopsis thaliana. Plant Cell 2007, 19, 2516–2530. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.Y.; Fujiwara, S.; Suh, S.S.; Kim, J.; Kim, Y.; Han, L.; David, K.; Putterill, J.; Nam, H.G.; Somers, D.E. ZEITLUPE is a circadian photoreceptor stabilized by GIGANTEA in blue light. Nature 2007, 449, 356–360. [Google Scholar] [CrossRef]
- Lee, C.M.; Li, M.W.; Feke, A.; Liu, W.; Saffer, A.M.; Gendron, J.M. GIGANTEA recruits the UBP12 and UBP13 deubiquitylases to regulate accumulation of the ZTL photoreceptor complex. Nat. Commun. 2019, 10, 3750. [Google Scholar] [CrossRef]
- Mishra, P.; Panigrahi, K.C. GIGANTEA—An emerging story. Front. Plant Sci. 2015, 6, 8. [Google Scholar] [CrossRef]
- Liu, L.; Xie, Y.; Yahaya, B.S.; Wu, F. GIGANTEA Unveiled: Exploring Its Diverse Roles and Mechanisms. Genes 2024, 15, 94. [Google Scholar] [CrossRef]
- Krahmer, J.; Goralogia, G.S.; Kubota, A.; Zardilis, A.; Johnson, R.S.; Song, Y.H.; MacCoss, M.J.; Le Bihan, T.; Halliday, K.J.; Imaizumi, T.; et al. Time-resolved interaction proteomics of the GIGANTEA protein under diurnal cycles in Arabidopsis. FEBS Lett. 2019, 593, 319–338. [Google Scholar] [CrossRef]
- Sawa, M.; Nusinow, D.A.; Kay, S.A.; Imaizumi, T. FKF1 and GIGANTEA complex formation is required for day-length measurement in Arabidopsis. Science 2007, 318, 261–265. [Google Scholar] [CrossRef]
- Song, Y.H.; Smith, R.W.; To, B.J.; Millar, A.J.; Imaizumi, T. FKF1 conveys timing information for CONSTANS stabilization in photoperiodic flowering. Science 2012, 336, 1045–1049. [Google Scholar] [CrossRef] [PubMed]
- Gould, P.D.; Locke, J.C.; Larue, C.; Southern, M.M.; Davis, S.J.; Hanano, S.; Moyle, R.; Milich, R.; Putterill, J.; Millar, A.J.; et al. The molecular basis of temperature compensation in the Arabidopsis circadian clock. Plant Cell 2006, 18, 1177–1187. [Google Scholar] [CrossRef]
- Kevei, E.; Gyula, P.; Feher, B.; Toth, R.; Viczian, A.; Kircher, S.; Rea, D.; Dorjgotov, D.; Schafer, E.; Millar, A.J.; et al. Arabidopsis thaliana circadian clock is regulated by the small GTPase LIP1. Curr. Biol. 2007, 17, 1456–1464. [Google Scholar] [CrossRef] [PubMed]
- Terecskei, K.; Toth, R.; Gyula, P.; Kevei, E.; Bindics, J.; Coupland, G.; Nagy, F.; Kozma-Bognar, L. The circadian clock-associated small GTPase LIGHT INSENSITIVE PERIOD1 suppresses light-controlled endoreplication and affects tolerance to salt stress in Arabidopsis. Plant Physiol. 2013, 161, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Hazen, S.P.; Borevitz, J.O.; Harmon, F.G.; Pruneda-Paz, J.L.; Schultz, T.F.; Yanovsky, M.J.; Liljegren, S.J.; Ecker, J.R.; Kay, S.A. Rapid array mapping of circadian clock and developmental mutations in Arabidopsis. Plant Physiol. 2005, 138, 990–997. [Google Scholar] [CrossRef]
- Kevei, E.; Gyula, P.; Hall, A.; Kozma-Bognar, L.; Kim, W.Y.; Eriksson, M.E.; Toth, R.; Hanano, S.; Feher, B.; Southern, M.M.; et al. Forward genetic analysis of the circadian clock separates the multiple functions of ZEITLUPE. Plant Physiol. 2006, 140, 933–945. [Google Scholar] [CrossRef]
- Jarillo, J.A.; Capel, J.; Tang, R.H.; Yang, H.Q.; Alonso, J.M.; Ecker, J.R.; Cashmore, A.R. An Arabidopsis circadian clock component interacts with both CRY1 and phyB. Nature 2001, 410, 487–490. [Google Scholar] [CrossRef]
- Green, R.M.; Tobin, E.M. Loss of the circadian clock-associated protein 1 in Arabidopsis results in altered clock-regulated gene expression. Proc. Natl. Acad. Sci. USA 1999, 96, 4176–4179. [Google Scholar] [CrossRef]
- Palagyi, A.; Terecskei, K.; Adam, E.; Kevei, E.; Kircher, S.; Merai, Z.; Schafer, E.; Nagy, F.; Kozma-Bognar, L. Functional analysis of amino-terminal domains of the photoreceptor phytochrome B. Plant Physiol. 2010, 153, 1834–1845. [Google Scholar] [CrossRef]
- Hall, A.; Brown, P. Monitoring circadian rhythms in Arabidopsis thaliana using luciferase reporter genes. Methods Mol. Biol. 2007, 362, 143–152. [Google Scholar] [CrossRef]
- Yasuhara, M.; Mitsui, S.; Hirano, H.; Takanabe, R.; Tokioka, Y.; Ihara, N.; Komatsu, A.; Seki, M.; Shinozaki, K.; Kiyosue, T. Identification of ASK and clock-associated proteins as molecular partners of LKP2 (LOV kelch protein 2) in Arabidopsis. J. Exp. Bot. 2004, 55, 2015–2027. [Google Scholar] [CrossRef]
- Berken, A.; Thomas, C.; Wittinghofer, A. A new family of RhoGEFs activates the Rop molecular switch in plants. Nature 2005, 436, 1176–1180. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.H.; Kim, T.L.; Kwon, Y.K.; Cho, M.H.; Yoo, J.Y.; Jeon, J.S.; Hahn, T.R.; Bhoo, S. Characterization of Arabidopsis RopGEF family genes in response to abiotic stresses. Plant Biotechnol. Rep. 2009, 3, 183–190. [Google Scholar] [CrossRef]
- Mas, P.; Kim, W.Y.; Somers, D.E.; Kay, S.A. Targeted degradation of TOC1 by ZTL modulates circadian function in Arabidopsis thaliana. Nature 2003, 426, 567–570. [Google Scholar] [CrossRef]
- Ji, H.; Pardo, J.M.; Batelli, G.; Van Oosten, M.J.; Bressan, R.A.; Li, X. The Salt Overly Sensitive (SOS) pathway: Established and emerging roles. Mol. Plant 2013, 6, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.Y.; Ali, Z.; Park, H.J.; Park, S.J.; Cha, J.Y.; Perez-Hormaeche, J.; Quintero, F.J.; Shin, G.; Kim, M.R.; Qiang, Z.; et al. Release of SOS2 kinase from sequestration with GIGANTEA determines salt tolerance in Arabidopsis. Nat. Commun. 2013, 4, 1352. [Google Scholar] [CrossRef] [PubMed]
- Sakuraba, Y.; Bulbul, S.; Piao, W.; Choi, G.; Paek, N.C. Arabidopsis EARLY FLOWERING3 increases salt tolerance by suppressing salt stress response pathways. Plant J. 2017, 92, 1106–1120. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.G.; Khakurel, D.; Hwang, J.W.; Nguyen, C.C.; Nam, B.; Shin, G.I.; Jeong, S.Y.; Ahn, G.; Cha, J.Y.; Lee, S.H.; et al. The E3 ubiquitin ligase COP1 regulates salt tolerance via GIGANTEA degradation in roots. Plant Cell Environ. 2024, 47, 3241–3252. [Google Scholar] [CrossRef]
- Martin-Tryon, E.L.; Kreps, J.A.; Harmer, S.L. GIGANTEA acts in blue light signaling and has biochemically separable roles in circadian clock and flowering time regulation. Plant Physiol. 2007, 143, 473–486. [Google Scholar] [CrossRef]
- Huq, E.; Tepperman, J.M.; Quail, P.H. GIGANTEA is a nuclear protein involved in phytochrome signaling in Arabidopsis. Proc. Natl. Acad. Sci. USA 2000, 97, 9789–9794. [Google Scholar] [CrossRef]
- Sawa, M.; Kay, S.A. GIGANTEA directly activates Flowering Locus T in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2011, 108, 11698–11703. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Liu, H.; Kong, J.; Yang, Y.; Zhang, N.; Li, R.; Yue, J.; Huang, J.; Li, C.; Cheung, A.Y.; et al. RopGEF7 regulates PLETHORA-dependent maintenance of the root stem cell niche in Arabidopsis. Plant Cell 2011, 23, 2880–2894. [Google Scholar] [CrossRef]
- Liu, T.; Liu, Q.; Yu, Z.; Wang, C.; Mai, H.; Liu, G.; Li, R.; Pang, G.; Chen, D.; Liu, H.; et al. eIF4E1 Regulates Arabidopsis Embryo Development and Root Growth by Interacting With RopGEF7. Front. Plant Sci. 2022, 13, 938476. [Google Scholar] [CrossRef]
- Mizoguchi, T.; Wright, L.; Fujiwara, S.; Cremer, F.; Lee, K.; Onouchi, H.; Mouradov, A.; Fowler, S.; Kamada, H.; Putterill, J.; et al. Distinct roles of GIGANTEA in promoting flowering and regulating circadian rhythms in Arabidopsis. Plant Cell 2005, 17, 2255–2270. [Google Scholar] [CrossRef] [PubMed]
- Gunl, M.; Liew, E.F.; David, K.; Putterill, J. Analysis of a post-translational steroid induction system for GIGANTEA in Arabidopsis. BMC Plant Biol. 2009, 9, 141. [Google Scholar] [CrossRef] [PubMed]
- Nohales, M.A.; Liu, W.; Duffy, T.; Nozue, K.; Sawa, M.; Pruneda-Paz, J.L.; Maloof, J.N.; Jacobsen, S.E.; Kay, S.A. Multi-level Modulation of Light Signaling by GIGANTEA Regulates Both the Output and Pace of the Circadian Clock. Dev. Cell 2019, 49, 840–851 e848. [Google Scholar] [CrossRef]
- Cha, J.Y.; Kim, J.; Kim, T.S.; Zeng, Q.; Wang, L.; Lee, S.Y.; Kim, W.Y.; Somers, D.E. GIGANTEA is a co-chaperone which facilitates maturation of ZEITLUPE in the Arabidopsis circadian clock. Nat. Commun. 2017, 8, 3. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Genotype | Period (h) ± SE | p-Values |
|---|---|---|---|
| CCR2:LUC | Col | 27.71 ± 0.25 | <0.001 |
| lip1-2 | 24.41 ± 0.29 | 0.009 | |
| gi-101 | 25.91 ± 0.23 | 0.35 | |
| lip1-2 gi-101 | 25.72 ± 0.51 | - | |
| CAB2:LUC | Col | 27.36 ± 0.52 | <0.001 |
| lip1-2 | 25.02 ± 0.27 | 0.016 | |
| toc1-4 | 25.88 ± 0.38 | 0.006 | |
| lip1-2 toc1-4 | 24.26 ± 0.41 | - | |
| CCR2:LUC | Col | 27.81 ± 0.33 | 0.002 |
| lip1-2 | 24.16 ± 0.71 | <0.001 | |
| ztl-3 | 31.12 ± 0.44 | 0.021 | |
| lip1-2 ztl-3 | 29.48 ± 0.63 | - | |
| CCR2:LUC | Col | 27.43 ± 0.38 | <0.001 |
| lip1-2 | 24.38 ± 0.54 | 0.027 | |
| cca1-1 | 25.79 ± 0.65 | 0.007 | |
| lip1-2 cca1-1 | 23.51 ± 0.18 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hajdu, A.; Nyári, D.; Terecskei, K.; Gyula, P.; Ádám, É.; Dobos, O.; Mérai, Z.; Kozma-Bognár, L. LIP1 Regulates the Plant Circadian Oscillator by Modulating the Function of the Clock Component GIGANTEA. Cells 2024, 13, 1503. https://doi.org/10.3390/cells13171503
Hajdu A, Nyári D, Terecskei K, Gyula P, Ádám É, Dobos O, Mérai Z, Kozma-Bognár L. LIP1 Regulates the Plant Circadian Oscillator by Modulating the Function of the Clock Component GIGANTEA. Cells. 2024; 13(17):1503. https://doi.org/10.3390/cells13171503
Chicago/Turabian StyleHajdu, Anita, Dóra Nyári, Kata Terecskei, Péter Gyula, Éva Ádám, Orsolya Dobos, Zsuzsanna Mérai, and László Kozma-Bognár. 2024. "LIP1 Regulates the Plant Circadian Oscillator by Modulating the Function of the Clock Component GIGANTEA" Cells 13, no. 17: 1503. https://doi.org/10.3390/cells13171503