
Human Endometrial Pericytes: A Comprehensive Overview of Their Physiological Functions and Implications in Uterine Disorders

 , ,
, , {kind=link}
Abstract
:1. Introduction
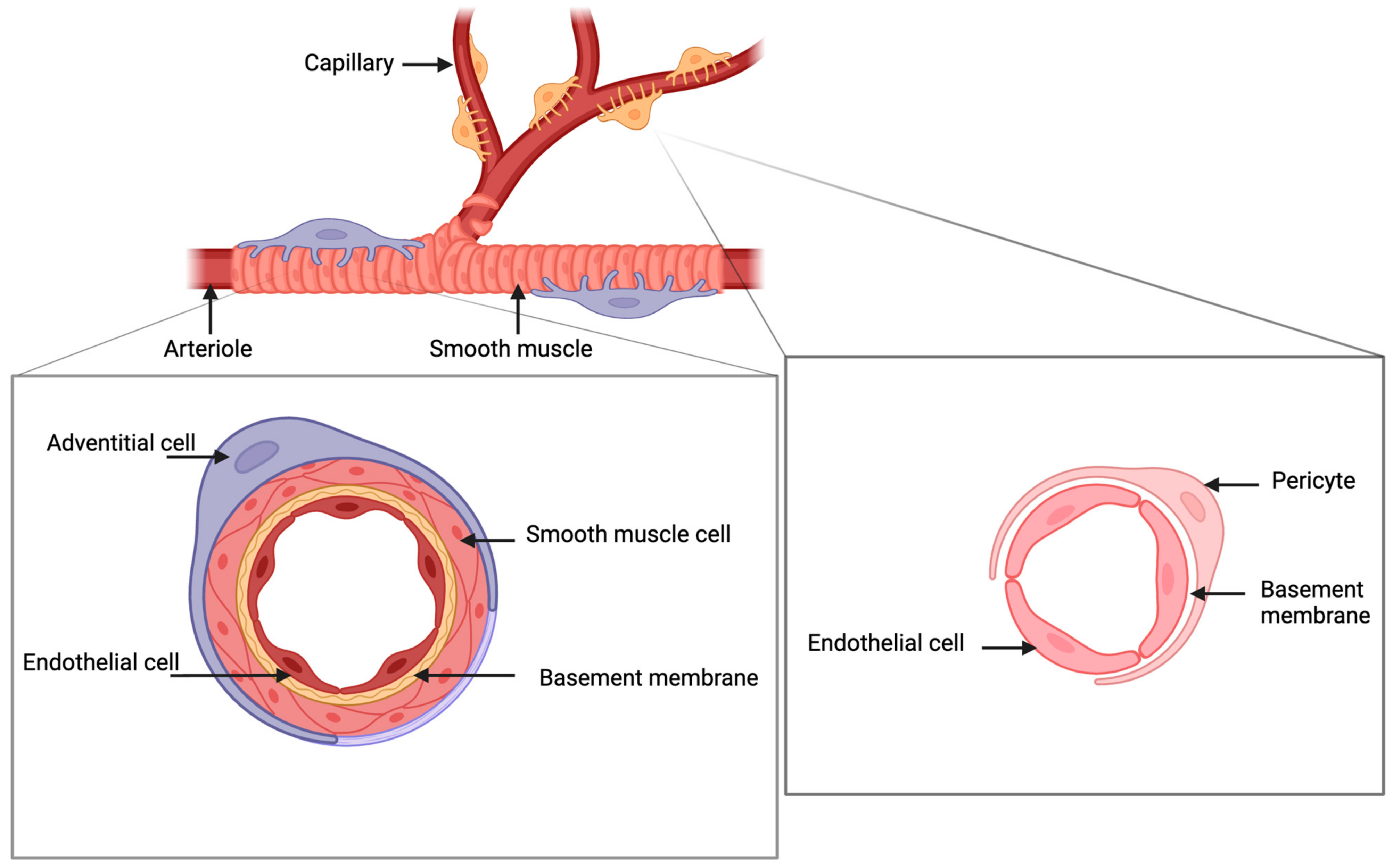
2. Characterization of Pericytes in the Endometrium
2.1. Origin
2.2. Molecular Markers
2.3. Plasticity
3. The Roles of Pericytes in the Endometrium
3.1. Vascularization
3.2. Immunomodulation
3.3. Endometrial Repair and Regeneration
3.4. Endometrial Decidulization
4. Pericytes in Endometrial Pathology
4.1. Intrauterine Adhesions and a Thin Endometrium
4.2. Adenomyosis and Endometriosis
4.3. Abnormal Uterine Bleeding and Related Conditions
5. Future Perspectives
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zimmermann, K.W. Der Feinere Bau der Blutcapillaren. Z. Anat. Entwicklungsgesch. 1923, 68, 29–109. [Google Scholar] [CrossRef]
- Krueger, M.; Bechmann, I. CNS Pericytes: Concepts, Misconceptions, and a Way Out. Glia 2010, 58, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hirschi, K.K.; D’Amore, P.A. Pericytes in the Microvasculature. Cardiovasc. Res. 1996, 32, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Armulik, A.; Genové, G.; Betsholtz, C. Pericytes: Developmental, Physiological, and Pathological Perspectives, Problems, and Promises. Dev. Cell 2011, 21, 193–215. [Google Scholar] [CrossRef]
- Queckbörner, S.; Davies, L.C.; von Grothusen, C.; Santamaria, X.; Simón, C.; Gemzell-Danielsson, K. Cellular Therapies for the Endometrium: An Update. Acta Obstet. Gynecol. Scand. 2019, 98, 672–677. [Google Scholar] [CrossRef]
- Gargett, C.E.; Schwab, K.E.; Deane, J.A. Endometrial Stem/Progenitor Cells: The First 10 Years. Hum. Reprod. Update 2016, 22, 137–163. [Google Scholar] [CrossRef]
- Cousins, F.L.; Filby, C.E.; Gargett, C.E. Endometrial Stem/Progenitor Cells–Their Role in Endometrial Repair and Regeneration. Front. Reprod. Health 2021, 3, 811537. [Google Scholar] [CrossRef]
- Nayak, N.R.; Brenner, R.M. Vascular Proliferation and Vascular Endothelial Growth Factor Expression in the Rhesus Macaque Endometrium. J. Clin. Endocrinol. Metab. 2002, 87, 1845–1855. [Google Scholar] [CrossRef]
- Evans, J.; Salamonsen, L.A. Inflammation, Leukocytes and Menstruation. Rev. Endocr. Metab. Disord. 2012, 13, 277–288. [Google Scholar] [CrossRef]
- Attwell, D.; Mishra, A.; Hall, C.N.; O’Farrell, F.M.; Dalkara, T. What Is a Pericyte? J. Cereb. Blood Flow Metab. 2016, 36, 451–455. [Google Scholar] [CrossRef]
- Smith, S.W.; Chand, S.; Savage, C.O.S. Biology of the Renal Pericyte. Nephrol. Dial. Transplant. 2012, 27, 2149–2155. [Google Scholar] [CrossRef] [PubMed]
- Ferland-McCollough, D.; Slater, S.; Richard, J.; Reni, C.; Mangialardi, G. Pericytes, an Overlooked Player in Vascular Pathobiology. Pharmacol. Ther. 2017, 171, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ding, L. Endometrial Perivascular Progenitor Cells and Uterus Regeneration. J. Pers. Med. 2021, 11, 477. [Google Scholar] [CrossRef]
- Crisan, M.; Corselli, M.; Chen, W.C.W.; Péault, B. Perivascular Cells for Regenerative Medicine. J. Cell. Mol. Med. 2012, 16, 2851–2860. [Google Scholar] [CrossRef]
- Saunders, W.B.; Bohnsack, B.L.; Faske, J.B.; Anthis, N.J.; Bayless, K.J.; Hirschi, K.K.; Davis, G.E. Coregulation of Vascular Tube Stabilization by Endothelial Cell TIMP-2 and Pericyte TIMP-3. J. Cell Biol. 2006, 175, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Fernández, R.; de la Mata, C.; Prados, A.; Perea, A.; Ruiz-Magaña, M.J.; Llorca, T.; Fernández-Rubio, P.; Blanco, O.; Abadía-Molina, A.C.; Olivares, E.G. Human Predecidual Stromal Cells Have Distinctive Characteristics of Pericytes: Cell Contractility, Chemotactic Activity, and Expression of Pericyte Markers and Angiogenic Factors. Placenta 2018, 61, 39–47. [Google Scholar] [CrossRef]
- Palomba, S.; Vitagliano, A.; Marci, R.; Caserta, D. Endometrial Scratching for Improving Endometrial Receptivity: A Critical Review of Old and New Clinical Evidence. Reprod. Sci. 2023, 30, 1701–1711. [Google Scholar] [CrossRef]
- Fan, Y.; Lee, R.W.K.; Ng, X.W.; Gargett, C.E.; Chan, J.K.Y. Subtle Changes in Perivascular Endometrial Mesenchymal Stem Cells after Local Endometrial Injury in Recurrent Implantation Failure. Sci. Rep. 2023, 13, 225. [Google Scholar] [CrossRef]
- Gharanei, S.; Fishwick, K.; Peter Durairaj, R.; Jin, T.; Siamantouras, E.; Liu, K.K.; Straube, A.; Lucas, E.S.; Weston, C.J.; Rantakari, P.; et al. Vascular Adhesion Protein-1 Determines the Cellular Properties of Endometrial Pericytes. Front. Cell Dev. Biol. 2021, 8, 621016. [Google Scholar] [CrossRef]
- Holm, A.; Heumann, T.; Augustin, H.G. Microvascular Mural Cell Organotypic Heterogeneity and Functional Plasticity. Trends Cell Biol. 2018, 28, 302–316. [Google Scholar] [CrossRef]
- Van Splunder, H.; Villacampa, P.; Martínez-Romero, A.; Graupera, M. Pericytes in the Disease Spotlight. Trends Cell Biol. 2024, 34, 58–71. [Google Scholar] [CrossRef]
- Africa, S.; Development, S.A.; Acyl, F.; Free, T.; Area, T.; Summit, T.; Committee, T.S.; Tfta, T.; Community, E.A.; African, S.; et al. Single-Cell RNA Sequencing Redefines the Mesenchymal Cell Landscape of Mouse Endometrium. FASEB J. 2020, 3, e21285. [Google Scholar] [CrossRef]
- Hennes, D.M.Z.B.; Rosamilia, A.; Werkmeister, J.A.; Gargett, C.E.; Mukherjee, S. Endometrial SUSD2+ Mesenchymal Stem/Stromal Cells in Tissue Engineering: Advances in Novel Cellular Constructs for Pelvic Organ Prolapse. J. Pers. Med. 2021, 11, 840. [Google Scholar] [CrossRef] [PubMed]
- Queckbörner, S.; von Grothusen, C.; Boggavarapu, N.R.; Francis, R.M.; Davies, L.C.; Gemzell-Danielsson, K. Stromal Heterogeneity in the Human Proliferative Endometrium-A Single-Cell RNA Sequencing Study. J. Pers. Med. 2021, 11, 448. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zeng, F.; Han, X.; Weng, J.; Gao, Y. Lineage Tracing: Technology Tool for Exploring the Development, Regeneration, and Disease of the Digestive System. Stem Cell Res. Ther. 2020, 11, 438. [Google Scholar] [CrossRef]
- Zipori, D. The Stem State: Plasticity Is Essential, Whereas Self-Renewal and Hierarchy Are Optional. Stem Cells 2005, 23, 719–726. [Google Scholar] [CrossRef]
- Zhu, X.; Yu, F.; Yan, G.; Hu, Y.; Sun, H.; Ding, L. Human Endometrial Perivascular Stem Cells Exhibit a Limited Potential to Regenerate Endometrium after Xenotransplantation. Human. Reprod. 2020, 36, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Gargett, C.E.; Rogers, P.A. Human Endometrial Angiogenesis. Reproduction 2001, 121, 181–186. [Google Scholar] [CrossRef]
- Bergers, G.; Song, S. The Role of Pericytes in Blood-Vessel Formation and Maintenance. Neuro Oncol. 2005, 7, 452–464. [Google Scholar] [CrossRef]
- Hickey, M.; Fraser, I.S.; Rogers, P.; Salamonsen, L. The Structure of Endometrial Microvessels. Hum. Reprod. 2000, 15, 57–66. [Google Scholar] [CrossRef]
- Huang, H. Pericyte-Endothelial Interactions in the Retinal Microvasculature. Int. J. Mol. Sci. 2020, 21, 7413. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Gao, J.; Ding, P.; Gao, Y. The Role of Endothelial Cell–Pericyte Interactions in Vascularization and Diseases. J. Adv. Res. 2024. epub ahead of print. [Google Scholar] [CrossRef]
- Eilken, H.M.; Diéguez-Hurtado, R.; Schmidt, I.; Nakayama, M.; Jeong, H.W.; Arf, H.; Adams, S.; Ferrara, N.; Adams, R.H. Pericytes Regulate VEGF-Induced Endothelial Sprouting through VEGFR1. Nat. Commun. 2017, 8, 1574. [Google Scholar] [CrossRef]
- Meyer, N.; Zenclussen, A.C. Immune Cells in the Uterine Remodeling: Are They the Target of Endocrine Disrupting Chemicals? Front. Immunol. 2020, 11, 246. [Google Scholar] [CrossRef] [PubMed]
- Berbic, M.; Fraser, I.S. Immunology of Normal and Abnormal Menstruation. Womens Health 2013, 9, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Maybin, J.A.; Critchley, H.O.D. Menstrual Physiology: Implications for Endometrial Pathology and Beyond. Hum. Reprod. Update 2015, 21, 748–761. [Google Scholar] [CrossRef] [PubMed]
- Gaceb, A.; Barbariga, M.; Özen, I.; Paul, G. The Pericyte Secretome: Potential Impact on Regeneration. Biochimie 2018, 155, 16–25. [Google Scholar] [CrossRef]
- Rustenhoven, J.; Jansson, D.; Smyth, L.C.; Dragunow, M. Brain Pericytes As Mediators of Neuroinflammation. Trends Pharmacol. Sci. 2017, 38, 291–304. [Google Scholar] [CrossRef]
- Thomas, H.M.; Cowin, A.J.; Mills, S.J. The Importance of Pericytes in Healing: Wounds and Other Pathologies. Int. J. Mol. Sci. 2017, 18, 1129. [Google Scholar] [CrossRef]
- Beckman, J.D.; Grazul-Bilska, A.T.; Johnson, M.L.; Reynolds, L.P.; Redmer, D.A. Isolation and Characterization of Ovine Luteal Pericytes and Effects of Nitric Oxide on Pericyte Expression of Angiogenic Factors. Endocrine 2006, 29, 467–476. [Google Scholar] [CrossRef]
- Gaceb, A.; Paul, G. Pericyte Secretome. Adv. Exp. Med. Biol. 2018, 1109, 139–163. [Google Scholar] [CrossRef] [PubMed]
- Ishitsuka, K.; Ago, T.; Arimura, K.; Nakamura, K.; Tokami, H.; Makihara, N.; Kuroda, J.; Kamouchi, M.; Kitazono, T. Neurotrophin Production in Brain Pericytes during Hypoxia: A Role of Pericytes for Neuroprotection. Microvasc. Res. 2012, 83, 352–359. [Google Scholar] [CrossRef]
- Sui, C.; Liao, Z.; Bai, J.; Hu, D.; Yue, J.; Yang, S. Current Knowledge on the Role of Extracellular Vesicles in Endometrial Receptivity. Eur. J. Med. Res. 2023, 28, 471. [Google Scholar] [CrossRef]
- Beal, J.R.; Ma, Q.; Bagchi, I.C.; Bagchi, M.K. Role of Endometrial Extracellular Vesicles in Mediating Cell-to-Cell Communication in the Uterus: A Review. Cells 2023, 12, 2584. [Google Scholar] [CrossRef]
- Gargett, C.E.; Masuda, H. Adult Stem Cells in the Endometrium. Mol. Hum. Reprod. 2010, 16, 818–834. [Google Scholar] [CrossRef]
- Crisan, M.; Corselli, M.; Chen, C.W.; Péault, B. Multilineage Stem Cells in the Adult: A Perivascular Legacy? Organogenesis 2011, 7, 101–104. [Google Scholar] [CrossRef]
- Spitzer, T.L.B.; Rojas, A.; Zelenko, Z.; Aghajanova, L.; Erikson, D.W.; Meyer, F.B.; Tamaresis, J.S.; Hamilton, A.E.; Irwin, J.C.; Giudice, L.C. Perivascular Human Endometrial Mesenchymal Stem Cells Express Pathways Relevant to Self-Renewal, Lineage Specification, and Functional Phenotype. Biol. Reprod. 2012, 86, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Barragan, F.; Irwin, J.C.; Balayan, S.; Erikson, D.W.; Chen, J.C.; Houshdaran, S.; Piltonen, T.T.; Spitzer, T.L.B.; George, A.; Rabban, J.T.; et al. Human Endometrial Fibroblasts Derived from Mesenchymal Progenitors Inherit Progesterone Resistance and Acquire an Inflammatory Phenotype in the Endometrial Niche in Endometriosis. Biol. Reprod. 2016, 94, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Goddard, L.M.; Murphy, T.J.; Org, T.; Enciso, J.M.; Hashimoto-Partyka, M.K.; Warren, C.M.; Domigan, C.K.; McDonald, A.I.; He, H.; Sanchez, L.A.; et al. Progesterone Receptor in the Vascular Endothelium Triggers Physiological Uterine Permeability Preimplantation. Cell 2014, 156, 549–562. [Google Scholar] [CrossRef]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L.; et al. A Perivascular Origin for Mesenchymal Stem Cells in Multiple Human Organs. Cell Stem Cell 2008, 3, 301–313. [Google Scholar] [CrossRef]
- Kirkwood, P.M.; Gibson, D.A.; Shaw, I.; Dobie, R.; Kelepouri, O.; Henderson, N.C.; Saunders, P.T.K. Single-Cell RNA Sequencing and Lineage Tracing Confirm Mesenchyme to Epithelial Transformation (MET) Contributes to Repair of the Endometrium at Menstruation. eLife 2022, 11, e77663. [Google Scholar] [CrossRef] [PubMed]
- Bouacida, A.; Rosset, P.; Trichet, V.; Guilloton, F.; Espagnolle, N.; Cordonier, T.; Heymann, D.; Layrolle, P.; Sensébé, L.; Deschaseaux, F. Pericyte-Like Progenitors Show High Immaturity and Engraftment Potential as Compared with Mesenchymal Stem Cells. PLoS ONE 2012, 7, e48648. [Google Scholar] [CrossRef]
- Gellersen, B.; Brosens, J.J. Cyclic Decidualization of the Human Endometrium in Reproductive Health and Failure. Endocr. Rev. 2014, 35, 851–905. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Nagasaka, T.; Nomura, M.; Tsukahara, S.I.; Kotani, Y.; Toda, S.; Murata, Y.; Itakura, A.; Mizutani, S. Premenstrual Disappearance of Aminopeptidase A in Endometrial Stromal Cells around Endometrial Spiral Arteries/Arterioles during the Decidual Change. J. Clin. Endocrinol. Metab. 2002, 87, 2303–2309. [Google Scholar] [CrossRef]
- Ahmed, A.; Li, X.F.; Shams, M.; Gregory, J.; Rollason, T.; Barnes, N.M.; Newton, J.R. Localization of the Angiotensin II and Its Receptor Subtype Expression in Human Endometrium and Identification of a Novel High-Affinity Angiotensin II Binding Site. J. Clin. Investig. 1995, 96, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Kitaya, K.; Yasuda, J.; Yagi, I.; Tada, Y.; Fushiki, S.; Honjo, H. IL-15 Expression at Human Endometrium and Decidua. Biol. Reprod. 2000, 63, 683–687. [Google Scholar] [CrossRef]
- Murakami, K.; Lee, Y.H.; Lucas, E.S.; Chan, Y.-W.W.; Durairaj, R.P.; Takeda, S.; Moore, J.D.; Tan, B.K.; Quenby, S.; Chan, J.K.Y.Y.; et al. Decidualization Induces a Secretome Switch in Perivascular Niche Cells of the Human Endometrium. Endocrinology 2014, 155, 4542–4553. [Google Scholar] [CrossRef]
- Harden, S.L.; Zhou, J.; Gharanei, S.; Diniz-da-Costa, M.; Lucas, E.S.; Cui, L.; Murakami, K.; Fang, J.; Chen, Q.; Brosens, J.J.; et al. Exometabolomic Analysis of Decidualizing Human Endometrial Stromal and Perivascular Cells. Front. Cell Dev. Biol. 2021, 9, 626619. [Google Scholar] [CrossRef]
- Gorsek Sparovec, T.; Markert, U.R.; Reif, P.; Schoell, W.; Moser, G.; Feichtinger, J.; Mihalic, Z.N.; Kargl, J.; Gargett, C.E.; Gold, D. The Fate of Human SUSD2+ Endometrial Mesenchymal Stem Cells during Decidualization. Stem Cell Res. 2022, 60, 102671. [Google Scholar] [CrossRef]
- Yu, D.; Wong, Y.M.; Cheong, Y.; Xia, E.; Li, T.C. Asherman Syndrome-One Century Later. Fertil. Steril. 2008, 89, 759–779. [Google Scholar] [CrossRef]
- Haines, M.; Taylor, C.W.; Fox, H.; Wells, M. Haines & Taylor Obstetrical and Gynaecological Pathology; Churchill Livingstone: Edinburgh, Scotland, 2003. [Google Scholar]
- Gargett, C.E.; Healy, D.L. Generating Receptive Endometrium in Asherman’s Syndrome. J. Hum. Reprod. Sci. 2011, 4, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Henderson, N.C.; Rieder, F.; Wynn, T.A. Fibrosis: From Mechanisms to Medicines. Nature 2020, 587, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Kramann, R.; Schneider, R.K.; Dirocco, D.P.; Machado, F.; Fleig, S.; Bondzie, P.A.; Henderson, J.M.; Ebert, B.L.; Humphreys, B.D. Perivascular Gli1+ Progenitors Are Key Contributors to Injury-Induced Organ Fibrosis. Cell Stem Cell 2015, 16, 51–66. [Google Scholar] [CrossRef]
- Lin, S.L.; Kisseleva, T.; Brenner, D.A.; Duffield, J.S. Pericytes and Perivascular Fibroblasts Are the Primary Source of Collagen-Producing Cells in Obstructive Fibrosis of the Kidney. Am. J. Pathol. 2008, 173, 1617–1627. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.T.; Chang, F.C.; Wu, C.F.; Chou, Y.H.; Hsu, H.L.; Chiang, W.C.; Shen, J.; Chen, Y.M.; Wu, K.D.; Tsai, T.J.; et al. Platelet-Derived Growth Factor Receptor Signaling Activates Pericyte-Myofibroblast Transition in Obstructive and Post-Ischemic Kidney Fibrosis. Kidney Int. 2011, 80, 1170–1181. [Google Scholar] [CrossRef]
- García-Solares, J.; Donnez, J.; Donnez, O.; Dolmans, M.M. Pathogenesis of Uterine Adenomyosis: Invagination or Metaplasia? Fertil. Steril. 2018, 109, 371–379. [Google Scholar] [CrossRef]
- Ahn, S.H.; Singh, V.; Tayade, C. Biomarkers in Endometriosis: Challenges and Opportunities. Fertil. Steril. 2017, 107, 523–532. [Google Scholar] [CrossRef]
- Li, Q.; Shi, J.; Yi, D.; Li, X.; Gu, Z.; Yan, H.; Leng, J. The Pathogenesis of Endometriosis and Adenomyosis: Insights from Single-Cell RNA Sequencing. Biol. Reprod. 2024, 110, 854–865. [Google Scholar] [CrossRef]
- Meenakshi, M.; Mccluggage, W.G. Vascular Involvement in Adenomyosis: Report of a Large Series of a Common Phenomenon with Observations on the Pathogenesis of Adenomyosis. Int. J. Gynecol. Pathol. 2010, 29, 117–121. [Google Scholar] [CrossRef]
- Sieiński, W. Tumor-like Intravascular Proliferations of the Stroma in Adenomyosis. Patol. Pol. 1993, 44, 1–4. [Google Scholar]
- Harmsen, M.J.; Arduç, A.; Bleeker, M.C.G.; Juffermans, L.J.M.; Griffioen, A.W.; Jordanova, E.S.; Huirne, J.A.F. Increased Angiogenesis and Lymphangiogenesis in Adenomyosis Visualized by Multiplex Immunohistochemistry. Int. J. Mol. Sci. 2022, 23, 8434. [Google Scholar] [CrossRef] [PubMed]
- Cheong, Y.; Cameron, I.T.; Critchley, H.O.D. Abnormal Uterine Bleeding. Br. Med. Bull. 2017, 123, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Andersson, E.; Zetterberg, E.; Vedin, I.; Hultenby, K.; Palmblad, J.; Mints, M. Low Pericyte Coverage of Endometrial Microvessels in Heavy Menstrual Bleeding Correlates with the Microvessel Expression of VEGF-A. Int. J. Mol. Med. 2015, 35, 433–438. [Google Scholar] [CrossRef]
- Starostanko, A.; Ayers, J.; Keeton, K.; Green, P. Endometrial Calcifications: A Novel Sonographic Marker of Endometrial Injury [29N]. Obstet. Gynecol. 2018, 131, 159S–160S. [Google Scholar] [CrossRef]
- Piltonen, T.T.; Chen, J.; Erikson, D.W.; Spitzer, T.L.B.; Barragan, F.; Rabban, J.T.; Huddleston, H.; Irwin, J.C.; Giudice, L.C. Mesenchymal Stem/Progenitors and Other Endometrial Cell Types from Women with Polycystic Ovary Syndrome (PCOS) Display Inflammatory and Oncogenic Potential. J. Clin. Endocrinol. Metab. 2013, 98, 3765–3775. [Google Scholar] [CrossRef] [PubMed]
- Palomba, S.; Piltonen, T.T.; Giudice, L.C. Endometrial Function in Women with Polycystic Ovary Syndrome: A Comprehensive Review. Hum. Reprod. Update 2021, 27, 584–618. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Frisendahl, C.; Piltonen, T.T.; Arffman, R.K.; Lalitkumar, P.G.; Gemzell-Danielsson, K. Human Endometrial Pericytes: A Comprehensive Overview of Their Physiological Functions and Implications in Uterine Disorders. Cells 2024, 13, 1510. https://doi.org/10.3390/cells13171510
Tang Y, Frisendahl C, Piltonen TT, Arffman RK, Lalitkumar PG, Gemzell-Danielsson K. Human Endometrial Pericytes: A Comprehensive Overview of Their Physiological Functions and Implications in Uterine Disorders. Cells. 2024; 13(17):1510. https://doi.org/10.3390/cells13171510
Chicago/Turabian StyleTang, Yiqun, Caroline Frisendahl, Terhi T. Piltonen, Riikka K. Arffman, Parameswaran Grace Lalitkumar, and Kristina Gemzell-Danielsson. 2024. "Human Endometrial Pericytes: A Comprehensive Overview of Their Physiological Functions and Implications in Uterine Disorders" Cells 13, no. 17: 1510. https://doi.org/10.3390/cells13171510