
Cytochalasins as Modulators of Stem Cell Differentiation

 , , , ,
, , , ,  and
and 
Abstract
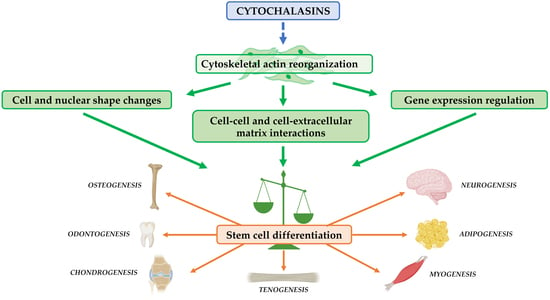
1. Introduction
2. Methods
Search Strategy and Study Selection
3. Cytochalasins
4. Human Mesenchymal Stem Cells
5. Cytochalasins and Osteogenesis
5.1. Effects of Cytochalasins on Cytoskeletal Organization and Cell Morphology during Osteogenesis
5.2. Effects of Cytochalasins on Cell–Extra-Cellular Matrix Interactions during Osteogenesis
5.3. Effects of Cytochalasins on the Cytoplasmic/Nuclear Actin Ratio in the Regulation of Osteogenesis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference Number | Title | Species and Cell Type | Molecular Treatment | Results * |
|---|---|---|---|---|
| [43] | Cytoskeletal Organization of Human Mesenchymal Stem Cells (MSC) Changes During Their Osteogenic Differentiation | hBM-MSCs | CD (0.1 μg/mL and 0.25 μg/mL) | ↓ |
| [44] | Inhibition of actin polymerization decreases osteogeneic differentiation of mesenchymal stem cells through p38 MAPK pathway | hBM-MSCs | CD (100–1000 ng/mL) | ↓ |
| [45] | Modulating the Actin Cytoskeleton Affects Mechanically Induced Signal Transduction and Differentiation in Mesenchymal Stem Cells | hBM-MSCs | CD (0.5 μM) | ↓ |
| [46] | TGFβ1-Induced Differentiation of Human Bone Marrow-Derived MSCs Is Mediated by Changes to the Actin Cytoskeleton | hBM-MSCs | CD (not specified) | ↓ |
| [47] | Regulation of the integrin αVβ3-actin filaments axis in early osteogenic differentiation of human mesenchymal stem cells under cyclic tensile stress | hBM-MSCs hADSCs | CD (0.2 μg/mL) | ↓ |
| [48] | Spatial organization and crosstalk of vimentin and actin stress fibers regulate the osteogenic differentiation of human adipose-derived stem cells | hASCs | CD (0.1 μg/mL) | ↓ |
| [49] | Cytochalasin B Influences Cytoskeletal Organization and Osteogenic Potential of Human Wharton’s Jelly Mesenchymal Stem Cells | hWJ-MSCs | CB (0.01–5 μM) | ↑ |
| [50] | Effects of aspect ratios of stem cells on lineage commitments with and without induction media | rBM-MSCs | CD (0.25 mg/mL) | ↓ |
| [51] | Spreading Shape and Area Regulate the Osteogenesis of Mesenchymal Stem Cells | mBM-MSCs | CD (1 μg/mL) | ↓ |
| [52] | Connective Tissue Growth Factor in Regulation of RhoA Mediated Cytoskeletal Tension Associated Osteogenesis of Mouse Adipose-Derived Stromal Cells | mASCs | CD (0.5 μg/mL) | (related to seeding density) |
| [53] | Role of Microtubules in Osteogenic Differentiation of Mesenchymal Stem Cells on 3D Nanofibrous Scaffolds | hBM-MSCs | CD (1 μM) | ↓ |
| [54] | The Role and Activation Mechanism of TAZ in Hierarchical Microgroove/Nanopore Topography-Mediated Regulation of Stem Cell Differentiation | hPDLSCs | CD (0.2 μM) | ↓ |
| [55] | Correlation between ECM guidance and actin polymerization on osteogenic differentiation of human adipose-derived stem cells | hASCs | CD (2 μM) | ↑ |
| [56] | Relationship of matrix stiffness and cell morphology in regulation of osteogenesis and adipogenesis of BMSCs | rBM-MSCs | CD (1 μM) | ↓ |
| [57] | Intranuclear Actin Regulates Osteogenesis | hBM-MSCs mBM-MSCs | CD (0.1 μg/mL) | ↑ |
| [58] | LRP6/filamentous-actin signaling facilitates osteogenic commitment in mechanically induced periodontal ligament stem cells | hPDLSCs | CD (0.2 μg/mL) | ↓ |
| [59] | Intranuclear Actin Structure Modulates Mesenchymal Stem Cell Differentiation | mBM-MSCs | CD (0.1 mg/mL) | ↑ |
| [60] | Osteogenic Stimulation of Human Adipose-Derived Mesenchymal Stem Cells Using a Fungal Metabolite That Suppresses the Polycomb Group Protein EZH2 | hASCs | CD (0.1 μg/mL) | ↑ |
| [61] | Validation of Osteogenic Properties of Cytochalasin D by High-Resolution RNA-Sequencing in Mesenchymal Stem Cells Derived from Bone Marrow and Adipose Tissues | hASCs hBM-MSCs mBM-MSCs | CD (0.1 μg/mL) | ↑ |
| [62] #1 | Mesenchymal stem cell and chondrocyte fates in a multishear microdevice are regulated by Yes-associated protein | MSCs | CD (not specified) | ↓ |
| [63] #2 | Topographic cues of a novel bilayered scaffold modulate dental pulp stem cells differentiation by regulating YAP signalling through cytoskeleton adjustments | hDPSCs | CD (1 μg/mL) | ↓ |

6. Cytochalasins and Adipogenesis
7. Cytochalasins and Chondrogenesis
8. Cytochalasins and Other Mesodermal Differentiation Commitments
8.1. Odontogenesis
8.2. Myogenesis
8.3. Tenogenesis
9. Cytochalasins and Neurogenesis
10. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mason, C.; Dunnill, P. A brief definition of regenerative medicine. Regen. Med. 2008, 3, 1–5. [Google Scholar] [CrossRef]
- Mahla, R.S. Stem Cells Applications in Regenerative Medicine and Disease Therapeutics. Int. J. Cell Biol. 2016, 2016, 6940283. [Google Scholar] [CrossRef] [PubMed]
- Leferink, A.M.; Chng, Y.C.; van Blitterswijk, C.A.; Moroni, L. Distribution and Viability of Fetal and Adult Human Bone Marrow Stromal Cells in a Biaxial Rotating Vessel Bioreactor after Seeding on Polymeric 3D Additive Manufactured Scaffolds. Front. Bioeng. Biotechnol. 2015, 3, 169. [Google Scholar] [CrossRef] [PubMed]
- Madrigal, M.; Rao, K.S.; Riordan, N.H. A review of therapeutic effects of mesenchymal stem cell secretions and induction of secretory modification by different culture methods. J. Transl. Med. 2014, 12, 260. [Google Scholar] [CrossRef] [PubMed]
- Castro-Manrreza, M.E.; Montesinos, J.J. Immunoregulation by mesenchymal stem cells: Biological aspects and clinical applications. J. Immunol. Res. 2015, 2015, 394917. [Google Scholar] [CrossRef] [PubMed]
- Turinetto, V.; Vitale, E.; Giachino, C. Senescence in Human Mesenchymal Stem Cells: Functional Changes and Implications in Stem Cell-Based Therapy. Int. J. Mol. Sci. 2016, 17, 1164. [Google Scholar] [CrossRef] [PubMed]
- Aldridge, D.C.; Armstrong, J.J.; Speake, R.N.; Turner, W.B. The cytochalasins, a new class of biologically active mould metabolites. Chem. Commun. 1967, 1, 26–27. [Google Scholar] [CrossRef]
- Brown, S.S.; Spudich, J.A. Mechanism of action of cytochalasin: Evidence that it binds to actin filament ends. J. Cell Biol. 1981, 88, 487–491. [Google Scholar] [CrossRef]
- Wodnicka, M.; Pierzchalska, M.; Bereiter-Hahn, J.; Kajstura, J. Comparative study on effects of cytochalasins B and D on F-actin content in different cell lines and different culture conditions. Folia Histochem. Cytobiol. 1992, 30, 107–111. [Google Scholar]
- Martino, F.; Perestrelo, A.R.; Vinarsky, V.; Pagliari, S.; Forte, G. Cellular Mechanotransduction: From Tension to Function. Front. Physiol. 2018, 9, 824. [Google Scholar] [CrossRef]
- Saidova, A.A.; Vorobjev, I.A. Lineage Commitment, Signaling Pathways, and the Cytoskeleton Systems in Mesenchymal Stem Cells. Tissue Eng. Part B Rev. 2020, 26, 13–25. [Google Scholar] [CrossRef]
- Mathieu, P.S.; Loboa, E.G. Cytoskeletal and focal adhesion influences on mesenchymal stem cell shape, mechanical properties, and differentiation down osteogenic, adipogenic, and chondrogenic pathways. Tissue Eng. Part B Rev. 2012, 18, 436–444. [Google Scholar] [CrossRef]
- Trendowski, M. Using cytochalasins to improve current chemotherapeutic approaches. Anticancer Agents Med. Chem. 2015, 15, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Rothweiler, W.; Tamm, C. Isolation and structure of Phomin. Experientia 1966, 22, 750–752. [Google Scholar] [CrossRef]
- Carter, S.B. Effects of Cytochalasins on Mammalian Cells. Nature 1967, 213, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Spudich, J.; Lin, S. Cytochalasin B, Its Interaction with actin and actomyosin from muscle (cell movement-microfilaments-rabbit striated muscle). Proc. Natl. Acad. Sci. USA 1972, 69, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Yahara, I.; Harada, F.; Sekita, S.; Yoshihira, K.; Natori, S. Correlation between effects of 24 different cytochalasins on cellular structures and cellular events and those on actin in vitro. J. Cell Biol. 1982, 92, 69–78. [Google Scholar] [CrossRef] [PubMed]
- MacLean-Fletcher, S.; Pollard, T.D. Mechanism of action of cytochalasin B on actin. Cell 1980, 20, 329–341. [Google Scholar] [CrossRef]
- Flanagan, M.D.; Lin, S. Cytochalasins block actin filament elongation by binding to high affinity sites associated with F-actin. J. Biol. Chem. 1980, 255, 835–838. [Google Scholar] [CrossRef]
- Ebstensen, R.D.; Plagemann, P.G. Cytochalasin B: Inhibition of glucose and glucosamine transport. Proc. Natl. Acad. Sci. USA 1972, 69, 1430–1434. [Google Scholar] [CrossRef]
- Miller, N.E.; Yin, J.A. Effects of cytochalasin B on low-density lipoproteins metabolism by cultured human fibroblasts. Biochim. Biophys. Acta 1978, 530, 145–150. [Google Scholar] [CrossRef]
- Simpson, P.B.; Armstrong, R.C. Intracellular signals and cytoskeletal elements involved in oligodendrocyte progenitor migration. Glia 1999, 26, 22–35. [Google Scholar] [CrossRef]
- Aldridge, D.C.; Turner, W.B. Structures of cytochalasins C and D. J. Chem. Soc. C. 1969, 923–928. [Google Scholar] [CrossRef]
- Brown, S.S.; Spudich, J.A. Cytochalasin inhibits the rate of elongation of actin filament fragments. J. Cell Biol. 1979, 83, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S.L.; Korn, E.D. Stimulation of actin ATPase activity by cytochalasins provides evidence for a new species of monomeric actin. J. Biol. Chem. 1981, 256, 8663–8670. [Google Scholar] [CrossRef] [PubMed]
- Goddette, D.W.; Frieden, C. Actin polymerization: The mechanism of action of cytochalasin D. J. Biol. Chem. 1986, 261, 15974–15980. [Google Scholar] [CrossRef]
- Bianco, P.; Robey, P.G.; Simmons, P.J. Mesenchymal stem cells: Revisiting history, concepts, and assays. Cell Stem Cell 2008, 2, 313–319. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Discher, D.E.; Peault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The international society for cellular therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Ciuffreda, M.C.; Malpasso, G.; Musaro, P.; Turco, V.; Gnecchi, M. Protocols for in vitro differentiation of human mesenchymal stem cells into Osteogenic, Chondrogenic and Adipogenic Lineages. Methods Mol. Biol. 2016, 1416, 149–158. [Google Scholar] [CrossRef]
- Heidari, B.; Shirazi, A.; Akhondi, M.M.; Hassanpour, H.; Behzadi, B.; Naderi, M.M.; Sarvari, A.; Borjian, S. Comparison of proliferative and multilineage differentiation potential of sheep mesenchymal stem cells derived from bone marrow, liver, and adipose tissue. Avicenna J. Med. Biotechnol. 2013, 5, 104–117. [Google Scholar]
- Ledesma-Martinez, E.; Mendoza-Nunez, V.M.; Santiago-Osorio, E. Mesenchymal stem cells derived from dental pulp: A review. Stem Cells Int. 2016, 2016, 4709572. [Google Scholar] [CrossRef]
- Chen, Y.T.; Wei, J.D.; Wang, J.P.; Lee, H.H.; Chiang, E.R.; Lai, H.C.; Chen, L.L.; Lee, Y.T.; Tsai, C.C.; Liu, C.L.; et al. Isolation of mesenchymal stem cells from human ligamentum flavum implicating etiology of ligamentum flavum hypertrophy. Spine 2011, 36, E1193–E1200. [Google Scholar] [CrossRef]
- Romanov, Y.A.; Svintsitskaya, V.A.; Smirnov, V.N. Searching for alternative sources of postnatal human mesenchymal stem cells: Candidate MSC-like cells from umbilical cord. Stem Cells 2003, 21, 105–110. [Google Scholar] [CrossRef]
- Papait, A.; Vertua, E.; Magatti, M.; Ceccariglia, S.; De Munari, S.; Silini, A.R.; Sheleg, M.; Ofir, R.; Parolini, O. Mesenchymal stromal cells from fetal and maternal placenta possess key similarities and differences: Potential implications for their applications in regenerative medicine. Cells 2020, 9, 127. [Google Scholar] [CrossRef] [PubMed]
- Hass, R.; Kasper, C.; Bohm, S.; Jacobs, R. Different populations and sources of human mesenchymal stem cells (MSC): A comparison of adult and neonatal tissue-derived MSC. Cell Commun. Signal. 2011, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Kern, S.; Eichler, H.; Stoeve, J.; Kluter, H.; Bieback, K. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells 2006, 24, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Aponte, P.M.; Caicedo, A. Stemness in Cancer: Stem Cells, Cancer Stem Cells, and Their Microenvironment. Stem Cells Int. 2017, 2017, 5619472. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Greenberger, J.S.; Epperly, M.W.; Goff, J.P.; Adler, C.; Leboff, M.S.; Glowacki, J. Age-related intrinsic changes in human bone-marrow-derived mesenchymal stem cells and their differentiation to osteoblasts. Aging Cell 2008, 7, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Praveen Kumar, L.; Kandoi, S.; Misra, R.; Vijayalakshmi, S.; Rajagopal, K.; Verma, R.S. The mesenchymal stem cell secretome: A new paradigm towards cell-free therapeutic mode in regenerative medicine. Cytokine Growth Factor Rev. 2019, 46, 1–9. [Google Scholar] [CrossRef]
- Dimarino, A.M.; Caplan, A.I.; Bonfield, T.L. Mesenchymal stem cells in tissue repair. Front. Immunol. 2013, 4, 201. [Google Scholar] [CrossRef]
- Zhang, B.; Yin, Y.; Lai, R.C.; Tan, S.S.; Choo, A.B.; Lim, S.K. Mesenchymal stem cells secrete immunologically active exosomes. Stem Cells Dev. 2014, 23, 1233–1244. [Google Scholar] [CrossRef]
- Rodríguez, J.P.; González, M.; Ríos, S.; Cambiazo, V. Cytoskeletal Organization of Human Mesenchymal Stem Cells (MSC) Changes During Their Osteogenic Differentiation. J. Cell. Biochem. 2004, 93, 721–731. [Google Scholar] [CrossRef]
- Sonowal, H.; Kumar, A.; Bhattacharyya, J.; Gogoi, P.K.; Jaganathan, B.G. Inhibition of actin polymerization decreases osteogeneic differentiation of mesenchymal stem cells through p38 MAPK pathway. J. Biomed. Sci. 2013, 20, 71. [Google Scholar] [CrossRef]
- Müller, P.; Langenbach, A.; Kaminski, A.; Rychly, J. Modulating the actin cytoskeleton affects mechanically induced signal transduction and differentiation in mesenchymal stem cells. PLoS ONE 2013, 8, e71283. [Google Scholar] [CrossRef]
- Elsafadi, M.; Manikandan, M.; Almalki, S.; Mobarak, M.; Atteya, M.; Iqbal, Z.; Hashmi, J.A.; Shaheen, S.; Alajez, N.; Alfayez, M.; et al. TGFβ1-Induced Differentiation of Human Bone Marrow-Derived MSCs Is Mediated by Changes to the Actin Cytoskeleton. Stem Cells Int. 2018, 2018, 6913594. [Google Scholar] [CrossRef]
- Peng, Y.; Qu, R.; Yang, Y.; Fan, T.; Sun, B.; Khan, A.U.; Wu, S.; Liu, W.; Zhu, J.; Chen, J.; et al. Regulation of the integrin αVβ3- actin filaments axis in early osteogenic differentiation of human mesenchymal stem cells under cyclic tensile stress. Cell Commun. Signal. 2023, 21, 308. [Google Scholar] [CrossRef]
- Fan, T.; Qu, R.; Jiang, X.; Yang, Y.; Sun, B.; Huang, X.; Zhou, Z.; Quyang, J.; Zhing, S.; Dai, J. Spatial organization and crosstalk of vimentin and actin stress fibers regulate the osteogenic differentiation of human adipose-derived stem cells. FASEB J. 2021, 35, 21175. [Google Scholar] [CrossRef] [PubMed]
- Pampanella, L.; Abruzzo, P.M.; Tassinari, R.; Alessandrini, A.; Petrocelli, G.; Ragazzini, G.; Cavallini, C.; Pizzuti, V.; Collura, N.; Canaider, S.; et al. Cytochalasin B Influences Cytoskeletal Organization and Osteogenic Potential of Human Wharton’s Jelly Mesenchymal Stem Cells. Pharmaceuticals 2023, 16, 289. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Peng, R.; Ding, J. Effects of aspect ratios of stem cells on lineage commitments with and without induction media. Biomaterials 2013, 34, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Sun, Q.; Wang, S.; Huo, B. Spreading Shape and Area Regulate the Osteogenesis of Mesenchymal Stem Cells. Tissue Eng. Regen. Med. 2019, 16, 573–583. [Google Scholar] [CrossRef]
- Xu, Y.; Wagner, D.R.; Bekerman, E.; Chiou, M.; James, A.W.; Carter, D.; Longaker, M.T. Connective tissue growth factor in regulation of RhoA mediated cytoskeletal tension associated osteogenesis of mouse adipose-derived stromal cells. PLoS ONE 2010, 5, e11279. [Google Scholar] [CrossRef] [PubMed]
- Meka, S.R.K.; Chacko, L.A.; Ravi, A.; Chatterjee, K.; Ananthanarayanan, V. Role of Microtubules in Osteogenic Differentiation of Mesenchymal Stem Cells on 3D Nanofibrous Scaffolds. ACS Biomater. Sci. Eng. 2017, 3, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Gao, Q.; Zheng, H.; Tian, Y.; Zheng, G.; Yao, X.; Zhang, J.; Wu, X.; Sui, L. The Role and Activation Mechanism of TAZ in Hierarchical Microgroove/Nanopore Topography-Mediated Regulation of Stem Cell Differentiation. Int. J. Neuromed. 2021, 16, 1021–1036. [Google Scholar] [CrossRef] [PubMed]
- Keller, V.; Deiwick, A.; Pflaum, M.; Schlie-Wolter, S. Correlation between ECM guidance and actin polymerization on osteogenic differentiation of human adipose-derived stem cells. Exp. Cell Res. 2016, 347, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Qiao, Y.; Quan, S.; Yang, C.; Li, J. Relationship of matrix stiffness and cell morphology in regulation of osteogenesis and adipogenesis of BMSCs. Mol. Biol. Rep. 2022, 9, 2677–2685. [Google Scholar] [CrossRef] [PubMed]
- Sen, B.; Xie, Z.; Uzer, G.; Thompson, W.R.; Styner, M.; Wu, X.; Rubin, J. Intranuclear Actin Regulates Osteogenesis. Stem Cells 2015, 33, 3065–3076. [Google Scholar] [CrossRef]
- Wang, J.; Yang, H.; Ma, X.; Liu, J.; Li, L.; Chen, L.; Wei, F. LRP6/filamentous-actin signaling facilitates osteogenic commitment in mechanically induced periodontal ligament stem cells. Cell. Mol. Biol. Lett. 2023, 28, 7. [Google Scholar] [CrossRef]
- Sen, B.; Uzer, G.; Samsonraj, R.M.; Xie, Z.; McGrath, C.; Styner, M.; Dudakovic, A.; van Wijnen, A.J.; Rubin, J. Intranuclear Actin Structure Modulates Mesenchymal Stem Cell Differentiation. Stem Cells 2017, 35, 1624–1635. [Google Scholar] [CrossRef]
- Samsonraj, R.M.; Dudakovic, A.; Manzar, B.; Sen, B.; Dietz, A.B.; Cool, S.M.; Rubin, J.; van Wijnen, A.J. Osteogenic Stimulation of Human Adipose-Derived Mesenchymal Stem Cells Using a Fungal Metabolite That Suppresses the Polycomb Group Protein EZH2. Stem Cells Trans. Med. 2018, 7, 197–209. [Google Scholar] [CrossRef]
- Samsonraj, R.M.; Paradise, C.R.; Dudakovic, A.; Sen, B.; Nair, A.A.; Dietz, A.B.; Deyle, D.R.; Cool, S.M.; Rubin, J.; van Wijnen, A.J. Validation of Osteogenic Properties of Cytochalasin D by High-Resolution RNA-Sequencing in Mesenchymal Stem Cells Derived from Bone Marrow and Adipose Tissues. Stem Cells Dev. 2018, 27, 1136–1145. [Google Scholar] [CrossRef]
- Zhong, W.; Tian, K.; Zheng, X.; Li, L.; Zhang, W.; Wang, S.; Qin, J. Mesenchymal stem cell and chondrocyte fates in a multishear microdevice are regulated by Yes-associated protein. Stem Cells Dev. 2013, 22, 2083–2093. [Google Scholar] [CrossRef]
- Du, Y.; Montoya, C.; Orrego, S.; Wei, X.; Ling, J.; Lelkes, P.I.; Yang, M. Topographic cues of a novel bilayered scaffold modulate dental pulp stem cells differentiation by regulating YAP signalling through cytoskeleton adjustments. Cell Prolif. 2019, 52, e12676. [Google Scholar] [CrossRef]
- Chen, L.; Hu, H.; Qiu, W.; Shi, K.; Kassem, M. Actin depolymerization enhances adipogenic differentiation in human stromal stem cells. Stem Cell Res. 2018, 29, 76–83. [Google Scholar] [CrossRef]
- Young, D.A.; Choi, Y.S.; Engler, A.J.; Christman, K.L. Stimulation of adipogenesis of adult adipose-derived stem cells using substrates that mimic the stiffness of adipose tissue. Biomaterials 2013, 34, 8581–8588. [Google Scholar] [CrossRef]
- Xu, B.; Ju, Y.; Song, G. Role of p38, ERK1/2, focal adhesion kinase, RhoA/ROCK and cytoskeleton in the adipogenesis of human mesenchymal stem cells. J. Biosci. Bioeng. 2014, 117, 624–631. [Google Scholar] [CrossRef]
- Feng, T.; Szabo, E.; Dziak, E.; Opas, M. Cytoskeletal disassembly and cell rounding promotes adipogenesis from ES cells. Stem Cell Rev. Rep. 2010, 6, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Bianconi, E.; Tassinari, R.; Alessandrini, A.; Ragazzini, G.; Cavallini, C.; Abruzzo, P.M.; Petrocelli, G.; Pampanella, L.; Casadei, R.; Maioli, M.; et al. Cytochalasin B Modulates Nanomechanical Patterning and Fate in Human Adipose-Derived Stem Cells. Cells 2022, 11, 1629. [Google Scholar] [CrossRef] [PubMed]
- Schiller, Z.A.; Schiele, N.R.; Sims, J.K.; Lee, K.; Kuo, C.K. Adipogenesis of adipose-derived stem cells may be regulated via the cytoskeleton at physiological oxygen levels in vitro. Stem Cell Res. Ther. 2013, 4, 79. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Wu, S.; Chu, L.T.; Kwong, H.K.; Hartanto, H.; Huang, Y.; Lam, M.L.; Lam, R.H.W.; Chen, T.H. Early Committed Clockwise Cell Chirality Upregulates Adipogenic Differentiation of Mesenchymal Stem Cells. Adv. Biosyst. 2020, 4, e2000161. [Google Scholar] [CrossRef] [PubMed]
- Connelly, J.T.; García, A.J.; Levenston, M.E. Interactions between integrin ligand density and cytoskeletal integrity regulate BMSC chondrogenesis. J. Cell. Physiol. 2008, 217, 145–154. [Google Scholar] [CrossRef]
- Zhang, Z.; Messana, J.; Hwang, N.S.; Elisseeff, J.H. Reorganization of actin filaments enhances chondrogenic differentiation of cells derived from murine embryonic stem cells. Biochem. Biophys. Res. Commun. 2006, 348, 421–427. [Google Scholar] [CrossRef]
- Hung, S.C.; Kuo, P.Y.; Chang, C.F.; Chen, T.H.; Ho, L.L. Alpha-smooth muscle actin expression and structure integrity in chondrogenesis of human mesenchymal stem cells. Cell Tissue Res. 2006, 324, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Raghothaman, D.; Leong, M.F.; Lim, T.C.; Wan, A.C.; Ser, Z.; Lee, E.H.; Yang, Z. Cell type dependent morphological adaptation in polyelectrolyte hydrogels governs chondrogenic fate. Biomed. Mater. 2016, 11, 025013. [Google Scholar] [CrossRef] [PubMed]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef]
- Jones, T.D.; Naimipour, H.; Sun, S.; Cho, M.; Alapati, S.B. Mechanical changes in human dental pulp stem cells during early odontogenic differentiation. J. Endod. 2015, 41, 50–55. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zheng, L.; Zhang, L.; Chen, L.; Jiang, J.; Zhou, X.; Wang, M.; Fan, Y. Static magnetic field regulates proliferation, migration, differentiation, and YAP/TAZ activation of human dental pulp stem cells. J. Tissue Eng. Regen. Med. 2018, 12, 2029–2040. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.; Antoku, S.; Östlund, C.; Worman, H.J.; Gundersen, G.G. Linker of nucleoskeleton and cytoskeleton (LINC) complex-mediated actin-dependent nuclear positioning orients centrosomes in migrating myoblasts. Nucleus 2015, 6, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Balogh, A.; Mège, R.M.; Sobel, A. Growth and cell density-dependent expression of stathmin in C2 myoblasts in culture. Exp. Cell Res. 1996, 224, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Formigli, L.; Meacci, E.; Sassoli, C.; Squecco, R.; Nosi, D.; Chellini, F.; Naro, F.; Francini, F.; Zecchi-Orlandini, S. Cytoskeleton/stretch-activated ion channel interaction regulates myogenic differentiation of skeletal myoblasts. J. Cell. Physiol. 2007, 211, 296–306. [Google Scholar] [CrossRef]
- Kneppers, A.; Verdijk, L.; de Theije, C.; Corten, M.; Gielen, E.; van Loon, L.; Schols, A.; Langen, R. A novel in vitro model for the assessment of postnatal myonuclear accretion. Skelet. Muscle 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Charó, N.L.; Galigniana, N.M.; Piwien-Pilipuk, G. Heterochromatin protein (HP)1γ is not only in the nucleus but also in the cytoplasm interacting with actin in both cell compartments. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 432–443. [Google Scholar] [CrossRef]
- Hoffmann, A.; Pelled, G.; Turgeman, G.; Eberle, P.; Zilberman, Y.; Shinar, H.; Keinan-Adamsky, K.; Winkel, A.; Shahab, S.; Navon, G.; et al. Neotendon formation induced by manipulation of the Smad8 signalling pathway in mesenchymal stem cells. J. Clin. Investig. 2006, 116, 940–952. [Google Scholar] [CrossRef]
- Xu, B.; Song, G.; Ju, Y.; Li, X.; Song, Y.; Watanabe, S. RhoA/ROCK, cytoskeletal dynamics, and focal adhesion kinase are required for mechanical stretch-induced tenogenic differentiation of human mesenchymal stem cells. J. Cell. Physiol. 2012, 227, 2722–2729. [Google Scholar] [CrossRef] [PubMed]
- Laplante, I.; Béliveau, R.; Paquin, J. RhoA/ROCK and Cdc42 regulate cell-cell contact and N-cadherin protein level during neurodetermination of P19 embryonal stem cells. J. Neurobiol. 2004, 60, 289–307. [Google Scholar] [CrossRef] [PubMed]
- Czeisler, C.; Short, A.; Nelson, T.; Gygli, P.; Ortiz, C.; Catacutan, F.P.; Stocker, B.; Cronin, J.; Lannutti, J.; Winter, J.; et al. Surface topography during neural stem cell differentiation regulates cell migration and cell morphology. J. Comp. Neurol. 2016, 524, 3485–3502. [Google Scholar] [CrossRef]
- Kronenberg, G.; Gertz, K.; Baldinger, T.; Kirste, I.; Eckart, S.; Yildirim, F.; Ji, S.; Heuser, I.; Schröck, H.; Hörtnagl, H.; et al. Impact of actin filament stabilization on adult hippocampal and olfactory bulb neurogenesis. J. Neurosci. 2010, 30, 3419–3431. [Google Scholar] [CrossRef]
- Ohara, Y.; Koganezawa, N.; Yamazaki, H.; Roppongi, R.T.; Sato, K.; Sekino, Y.; Shirao, T. Early-stage development of human induced pluripotent stem cell-derived neurons. J. Neurosci. Res. 2015, 93, 1804–1813. [Google Scholar] [CrossRef]
- Sawangmake, C.; Pavasant, P.; Chansiripornchai, P.; Osathanon, T. High glucose condition suppresses neurosphere formation by human periodontal ligament-derived mesenchymal stem cells. J. Cell. Biochem. 2014, 115, 928–939. [Google Scholar] [CrossRef]
- Su, X.; Zhou, H.; Bao, G.; Wang, J.; Liu, L.; Zheng, Q.; Guo, M.; Zhang, J. Nanomorphological and mechanical reconstruction of mesenchymal stem cells during early apoptosis detected by atomic force microscopy. Biol. Open 2020, 9, bio048108. [Google Scholar] [CrossRef]


| Reference Number | Title | Species and Cell Type | Molecular Treatment | Results * |
|---|---|---|---|---|
| [64] | Actin depolymerization enhances adipogenic differentiation in human stromal stem cells | hBM-MSCs | CD (1–20 μM) | ↑ |
| [56] | Relationship of matrix stiffness and cell morphology in regulation of osteogenesis and adipogenesis of BMSCs | rBM-MSCs | CD (1 μM) | ↑ |
| [65] | Stimulation of adipogenesis of adult adipose-derived stem cells using substrates that mimic the stiffness of adipose tissue | hASCs | CD (0.25 μg/mL) | ↑ |
| [54] | The Role and Activation Mechanism of TAZ in Hierarchical Microgroove/Nanopore Topography-Mediated Regulation of Stem Cell Differentiation | hPDLSCs | CD (0.2 μM) | ↑ |
| [66] | Role of p38, ERK1/2, focal adhesion kinase, RhoA/ROCK and cytoskeleton in the adipogenesis of human mesenchymal stem cells | hMSCs | CD (0.02 μg/mL) | ↑ |
| [67] | Cytoskeletal Disassembly and cell rounding promotes adipogenesis from ES cells | mESCs | CD (0.2–20 μM) | ↑ |
| [44] | Inhibition of actin polymerization decreases osteogeneic differentiation of mesenchymal stem cells through p38 MAPK pathway | hBM-MSCs | CD (100–1000 ng/mL) | ↑ |
| [45] | Modulating the Actin Cytoskeleton Affects Mechanically Induced Signal Transduction and Differentiation in Mesenchymal Stem Cells | hBM-MSCs | CD (0.5 μM) | ↑ |
| [50] | Effects of aspect ratios of stem cells on lineage commitments with and without induction media | rBM-MSCs | CD (0.25 mg/mL) | ↑ |
| [68] | Cytochalasin B Modulates Nanomechanical Patterning and Fate in Human Adipose-Derived Stem Cells | hASCs | CB (1–10 μM) | ↑ |
| [69] | Adipogenesis of adipose-derived stem cells may be regulated via the cytoskeleton at physiological oxygen levels in vitro | hASCs | CD (2 μM) | ↑ |
| [59] | Intranuclear Actin Structure Modulates Mesenchymal Stem Cell Differentiation | mBM-MSCs | CD (0.1 mg/mL) | ↑ |
| [46] | TGFβ1-Induced Differentiation of Human Bone Marrow-Derived MSCs Is Mediated by Changes to the Actin Cytoskeleton | hBM-MSCs | CD (not specified) | ↑ |
| [57] | Intranuclear Actin Regulates Osteogenesis | hBM-MSCs mBM-MSCs | CD (0.1 μg/mL) | ↑ |
| [70] | Early Committed Clockwise Cell Chirality Upregulates Adipogenic Differentiation of Mesenchymal Stem Cells | hMSCs | CD (0.02–0.2 μM) | ↑ |
| [52] #1 | Connective Tissue Growth Factor in Regulation of RhoA Mediated Cytoskeletal Tension Associated Osteogenesis of Mouse Adipose-Derived Stromal Cells | mASCs | CD (0.5 μg/mL) | ↓ |
| [62] #2 | Mesenchymal stem cell and chondrocyte fates in a multishear microdevice are regulated by Yes-associated protein | MSCs | CD (not specified) | ↑ |
| Reference Number | Title | Species and Cell Type | Molecular Treatment | Results * |
|---|---|---|---|---|
| [72] | Reorganization of actin filaments enhances chondrogenic differentiation of cells derived from murine embryonic stem cells | mEBCs | CD (0.5–10 μg/mL) | ↑ |
| [73] | Alpha-smooth muscle actin expression and structure integrity in chondrogenesis of human mesenchymal stem cells | hBM-MSCs | CD (10 μM) | cytotoxic dose |
| [71] | Interactions Between Integrin Ligand Density and Cytoskeletal Integrity Regulate BMSC Chondrogenesis | cBM-MSCs | CD (0.3 μM) | ↑ |
| [74] | Cell type dependent morphological adaptation in polyelectrolyte hydrogels governs chondrogenic fate | hBM-MSCs | CD (0.5 μM) | ↓ |
| [62] $ | Mesenchymal stem cell and chondrocyte fates in a multishear microdevice are regulated by Yes-associated protein | MSCs | CD (not specified) | ↑ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pampanella, L.; Petrocelli, G.; Abruzzo, P.M.; Zucchini, C.; Canaider, S.; Ventura, C.; Facchin, F. Cytochalasins as Modulators of Stem Cell Differentiation. Cells 2024, 13, 400. https://doi.org/10.3390/cells13050400
Pampanella L, Petrocelli G, Abruzzo PM, Zucchini C, Canaider S, Ventura C, Facchin F. Cytochalasins as Modulators of Stem Cell Differentiation. Cells. 2024; 13(5):400. https://doi.org/10.3390/cells13050400
Chicago/Turabian StylePampanella, Luca, Giovannamaria Petrocelli, Provvidenza Maria Abruzzo, Cinzia Zucchini, Silvia Canaider, Carlo Ventura, and Federica Facchin. 2024. "Cytochalasins as Modulators of Stem Cell Differentiation" Cells 13, no. 5: 400. https://doi.org/10.3390/cells13050400
APA StylePampanella, L., Petrocelli, G., Abruzzo, P. M., Zucchini, C., Canaider, S., Ventura, C., & Facchin, F. (2024). Cytochalasins as Modulators of Stem Cell Differentiation. Cells, 13(5), 400. https://doi.org/10.3390/cells13050400