
Glycogen Granules Are Degraded by Non-Selective Autophagy in Nitrogen-Starved Komagataella phaffii


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Plasmids
2.2. Iodine Staining for Glycogen
2.3. Biochemical Analysis
2.4. Fluorescence Microscopy
2.5. Statistical Analysis
3. Results
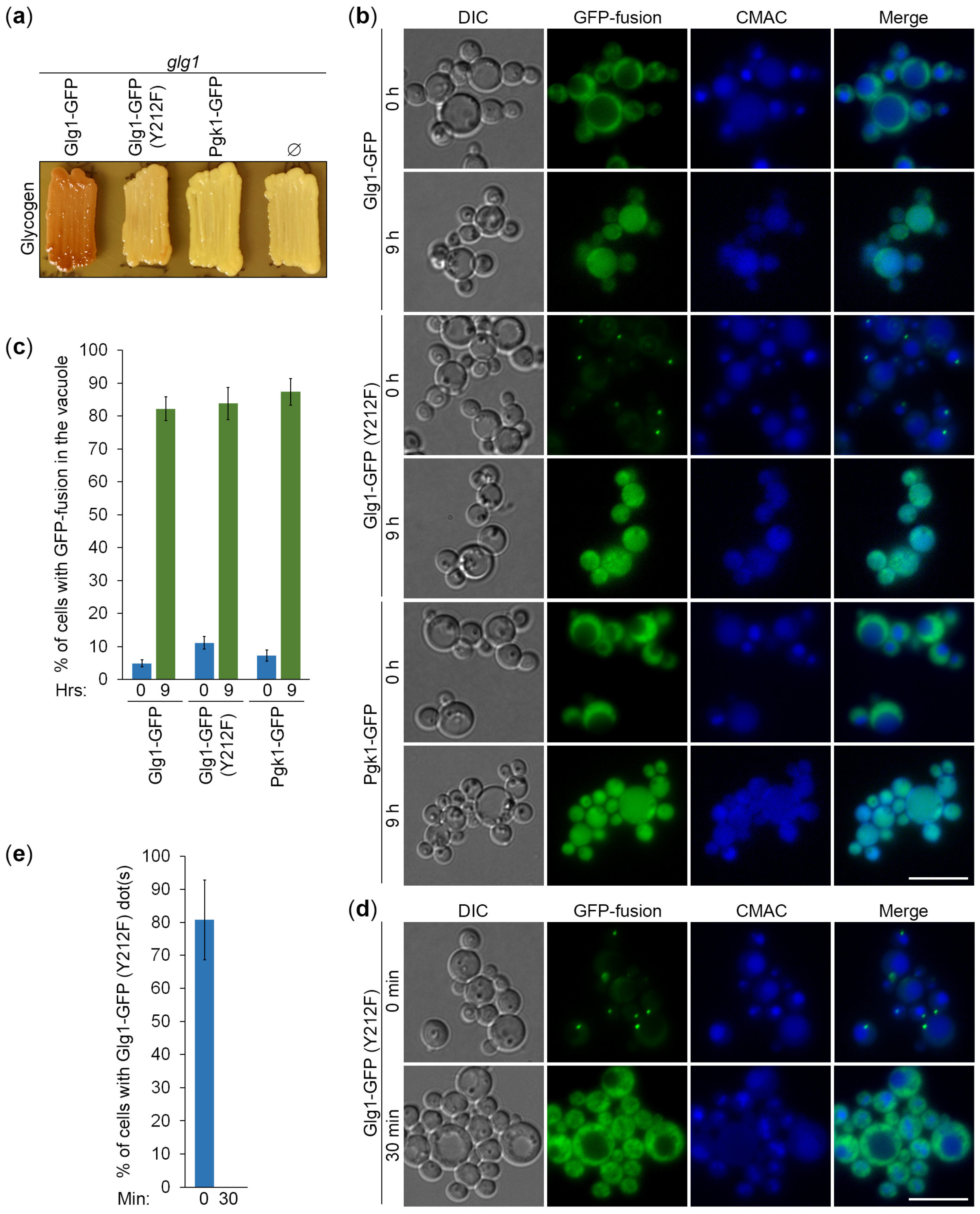
3.1. Glg1-GFP Fusion Protein Is Functional in Glycogen Synthesis
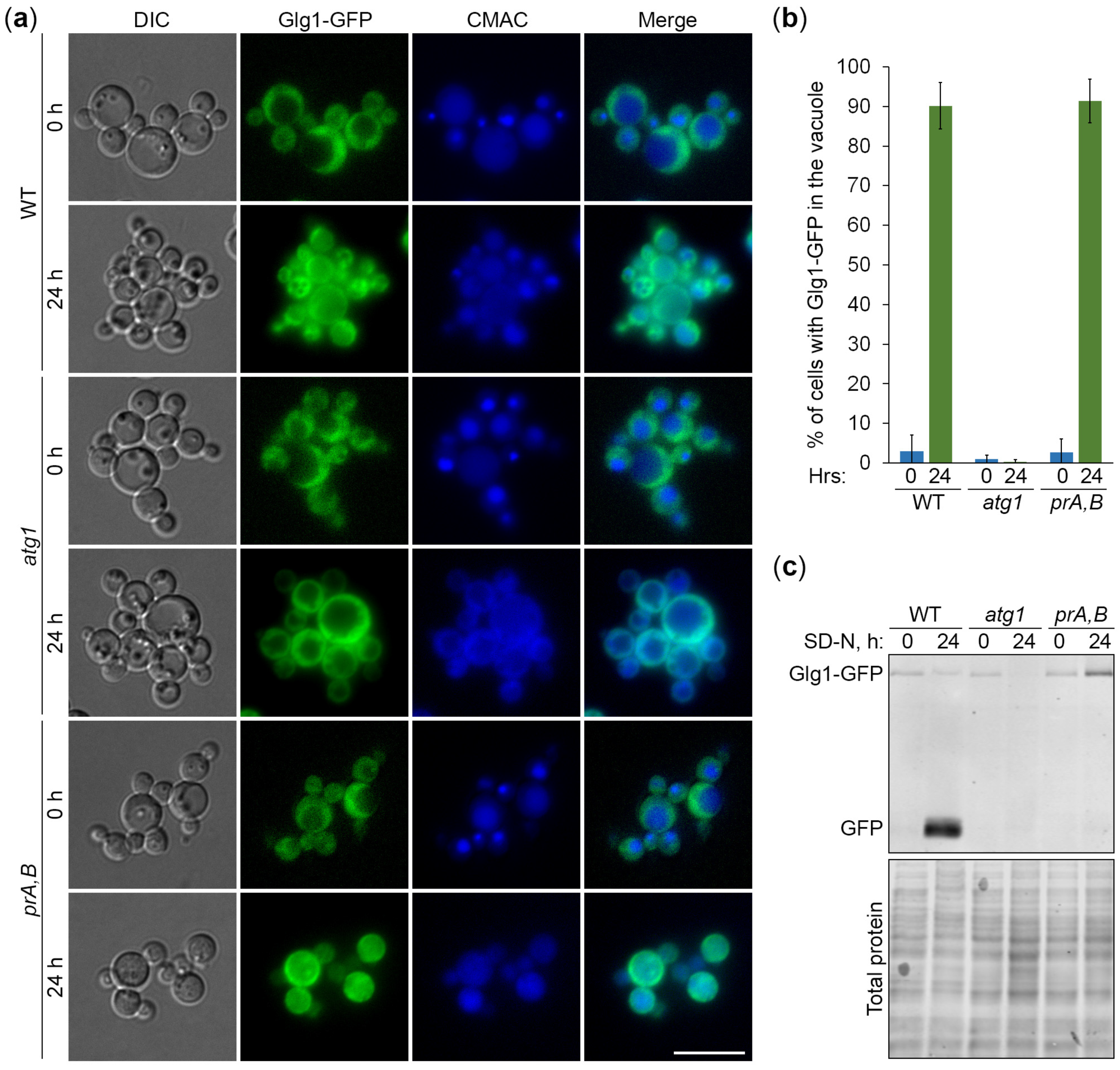
3.2. Degradation of Glycogen Granules Depends on Autophagy and Vacuole
3.3. Autophagy of Glycogen Granules Is Independent of Atg11
3.4. Autophagy of Glycogen Granules Is a Non-Selective Process
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Prats, C.; Graham, T.E.; Shearer, J. The dynamic life of the glycogen granule. J. Biol. Chem. 2018, 293, 7089–7098. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hu, Z. Is liver glycogen fragility a possible drug target for diabetes? FASEB J. 2020, 34, 3–15. [Google Scholar] [CrossRef]
- Liu, Q.H.; Tang, J.W.; Wen, P.B.; Wang, M.M.; Zhang, X.; Wang, L. From Prokaryotes to Eukaryotes: Insights Into the Molecular Structure of Glycogen Particles. Front. Mol. Biosci. 2021, 8, 673315. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Mu, J.; Farkas, I.; Huang, D.; Goebl, M.G.; Roach, P.J. Requirement of the self-glucosylating initiator proteins Glg1p and Glg2p for glycogen accumulation in Saccharomyces cerevisiae. Mol. Cell. Biol. 1995, 15, 6632–6640. [Google Scholar] [CrossRef]
- Mu, J.; Cheng, C.; Roach, P.J. Initiation of glycogen synthesis in yeast. Requirement of multiple tyrosine residues for function of the self-glucosylating Glg proteins in vivo. J. Biol. Chem. 1996, 271, 26554–26560. [Google Scholar] [CrossRef]
- Wilson, W.A.; Roach, P.J.; Montero, M.; Baroja-Fernández, E.; Muñoz, F.J.; Eydallin, G.; Viale, A.M.; Pozueta-Romero, J. Regulation of glycogen metabolism in yeast and bacteria. FEMS Microbiol. Rev. 2010, 34, 952–985. [Google Scholar] [CrossRef] [PubMed]
- Adeva-Andany, M.M.; González-Lucán, M.; Donapetry-García, C.; Fernández-Fernández, C.; Ameneiros-Rodríguez, E. Glycogen metabolism in humans. BBA Clin. 2016, 5, 85–100. [Google Scholar] [CrossRef]
- Raben, N.; Schreiner, C.; Baum, R.; Takikita, S.; Xu, S.; Xie, T.; Myerowitz, R.; Komatsu, M.; Van der Meulen, J.H.; Nagaraju, K.; et al. Suppression of autophagy permits successful enzyme replacement therapy in a lysosomal storage disorder--murine Pompe disease. Autophagy 2010, 6, 1078–1089. [Google Scholar] [CrossRef]
- Jiang, S.; Heller, B.; Tagliabracci, V.S.; Zhai, L.; Irimia, J.M.; DePaoli-Roach, A.A.; Wells, C.D.; Skurat, A.V.; Roach, P.J. Starch binding domain-containing protein 1/genethonin 1 is a novel participant in glycogen metabolism. J. Biol. Chem. 2010, 285, 34960–34971. [Google Scholar] [CrossRef]
- Jiang, S.; Wells, C.D.; Roach, P.J. Starch-binding domain-containing protein 1 (Stbd1) and glycogen metabolism: Identification of the Atg8 family interacting motif (AIM) in Stbd1 required for interaction with GABARAPL1. Biochem. Biophys. Res. Commun. 2011, 413, 420–425. [Google Scholar] [CrossRef]
- Yi, H.; Fredrickson, K.B.; Das, S.; Kishnani, P.S.; Sun, B. Stbd1 is highly elevated in skeletal muscle of Pompe disease mice but suppression of its expression does not affect lysosomal glycogen accumulation. Mol. Genet. Metab. 2013, 109, 312–314. [Google Scholar] [CrossRef]
- Sun, T.; Yi, H.; Yang, C.; Kishnani, P.S.; Sun, B. Starch Binding Domain-containing Protein 1 Plays a Dominant Role in Glycogen Transport to Lysosomes in Liver. J. Biol. Chem. 2016, 291, 16479–16484. [Google Scholar] [CrossRef]
- Cregg, J.M.; Russell, K.A. Transformation. Methods Mol. Biol. 1998, 103, 27–39. [Google Scholar] [CrossRef]
- Gould, S.J.; McCollum, D.; Spong, A.P.; Heyman, J.A.; Subramani, S. Development of the yeast Pichia pastoris as a model organism for a genetic and molecular analysis of peroxisome assembly. Yeast 1992, 8, 613–628. [Google Scholar] [CrossRef]
- Stromhaug, P.E.; Bevan, A.; Dunn, W.A., Jr. GSA11 encodes a unique 208-kDa protein required for pexophagy and autophagy in Pichia pastoris. J. Biol. Chem. 2001, 276, 42422–42435. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kamada, Y.; Stromhaug, P.E.; Guan, J.; Hefner-Gravink, A.; Baba, M.; Scott, S.V.; Ohsumi, Y.; Dunn, W.A., Jr.; Klionsky, D.J. Cvt9/Gsa9 functions in sequestering selective cytosolic cargo destined for the vacuole. J. Cell Biol. 2001, 153, 381–396. [Google Scholar] [CrossRef] [PubMed]
- Tuttle, D.L.; Dunn, W.A., Jr. Divergent modes of autophagy in the methylotrophic yeast Pichia pastoris. J. Cell Sci. 1995, 108 Pt 1, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Shroff, A.; Nazarko, T.Y. Komagataella phaffii Cue5 Piggybacks on Lipid Droplets for Its Vacuolar Degradation during Stationary Phase Lipophagy. Cells 2022, 11, 215. [Google Scholar] [CrossRef] [PubMed]
- Enjalbert, B.; Parrou, J.L.; Vincent, O.; François, J. Mitochondrial respiratory mutants of Saccharomyces cerevisiae accumulate glycogen and readily mobilize it in a glucose-depleted medium. Microbiology 2000, 146 Pt 10, 2685–2694. [Google Scholar] [CrossRef] [PubMed]
- Torija, M.J.; Novo, M.; Lemassu, A.; Wilson, W.; Roach, P.J.; François, J.; Parrou, J.L. Glycogen synthesis in the absence of glycogenin in the yeast Saccharomyces cerevisiae. FEBS Lett. 2005, 579, 3999–4004. [Google Scholar] [CrossRef] [PubMed]
- Baerends, R.J.; Faber, K.N.; Kram, A.M.; Kiel, J.A.; van der Klei, I.J.; Veenhuis, M. A stretch of positively charged amino acids at the N terminus of Hansenula polymorpha Pex3p is involved in incorporation of the protein into the peroxisomal membrane. J. Biol. Chem. 2000, 275, 9986–9995. [Google Scholar] [CrossRef]
- Stefan, C.J.; Blumer, K.J. A syntaxin homolog encoded by VAM3 mediates down-regulation of a yeast G protein-coupled receptor. J. Biol. Chem. 1999, 274, 1835–1841. [Google Scholar] [CrossRef]
- Kumar, R.; Rahman, M.A.; Nazarko, T.Y. Nitrogen Starvation and Stationary Phase Lipophagy Have Distinct Molecular Mechanisms. Int. J. Mol. Sci. 2020, 21, 9094. [Google Scholar] [CrossRef] [PubMed]
- Zientara-Rytter, K.; Subramani, S. Mechanistic Insights into the Role of Atg11 in Selective Autophagy. J. Mol. Biol. 2020, 432, 104–122. [Google Scholar] [CrossRef]
- Welter, E.; Thumm, M.; Krick, R. Quantification of nonselective bulk autophagy in S. cerevisiae using Pgk1-GFP. Autophagy 2010, 6, 794–797. [Google Scholar] [CrossRef] [PubMed]
- Case, L.E.; Kishnani, P.S. Physical therapy management of Pompe disease. Genet. Med. 2006, 8, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Koutsifeli, P.; Varma, U.; Daniels, L.J.; Annandale, M.; Li, X.; Neale, J.P.H.; Hayes, S.; Weeks, K.L.; James, S.; Delbridge, L.M.D.; et al. Glycogen-autophagy: Molecular machinery and cellular mechanisms of glycophagy. J. Biol. Chem. 2022, 298, 102093. [Google Scholar] [CrossRef]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutant | Strain | Background | Genotype and Plasmid | Source |
|---|---|---|---|---|
| WT | PPY12h | PPY12h | arg4 his4 | [14] |
| WT | SRK147 | PPY12h | his4::pRK22 (PGLG1-GLG1-GFP, HIS4) | This study |
| atg1 | R12 | GS115 | atg1-1::ZeocinR his4 | [15] |
| atg1 | SRK149 | R12 | his4::pRK22 (PGLG1-GLG1-GFP, HIS4) | This study |
| atg11 | R8 | GS115 | atg11-2::ZeocinR his4 | [16] |
| atg11 | SNW7 | R8 | his4::pRK22 (PGLG1-GLG1-GFP, HIS4) | This study |
| glg1 | SNW49 | PPY12h | ∆glg1::ZeocinR (pNW9) | This study |
| glg1 | SNW65 | SNW49 | his4::pRK22 (PGLG1-GLG1-GFP, HIS4) | This study |
| glg1 | SNW80 | SNW49 | his4::pNW11 (PGLG1-GLG1Y212F-GFP, HIS4) | This study |
| glg1 | SNW82 | SNW49 | his4::pNW10 (PGLG1-PGK1-GFP, HIS4) | This study |
| pep4 prb1 | SMD1163 | GS115 | pep4 prb1 his4 | [17] |
| pep4 prb1 | SRK151 | SMD1163 | his4::pRK22 (PGLG1-GLG1-GFP, HIS4) | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wijewantha, N.V.; Kumar, R.; Nazarko, T.Y. Glycogen Granules Are Degraded by Non-Selective Autophagy in Nitrogen-Starved Komagataella phaffii. Cells 2024, 13, 467. https://doi.org/10.3390/cells13060467
Wijewantha NV, Kumar R, Nazarko TY. Glycogen Granules Are Degraded by Non-Selective Autophagy in Nitrogen-Starved Komagataella phaffii. Cells. 2024; 13(6):467. https://doi.org/10.3390/cells13060467
Chicago/Turabian StyleWijewantha, Nimna V., Ravinder Kumar, and Taras Y. Nazarko. 2024. "Glycogen Granules Are Degraded by Non-Selective Autophagy in Nitrogen-Starved Komagataella phaffii" Cells 13, no. 6: 467. https://doi.org/10.3390/cells13060467