
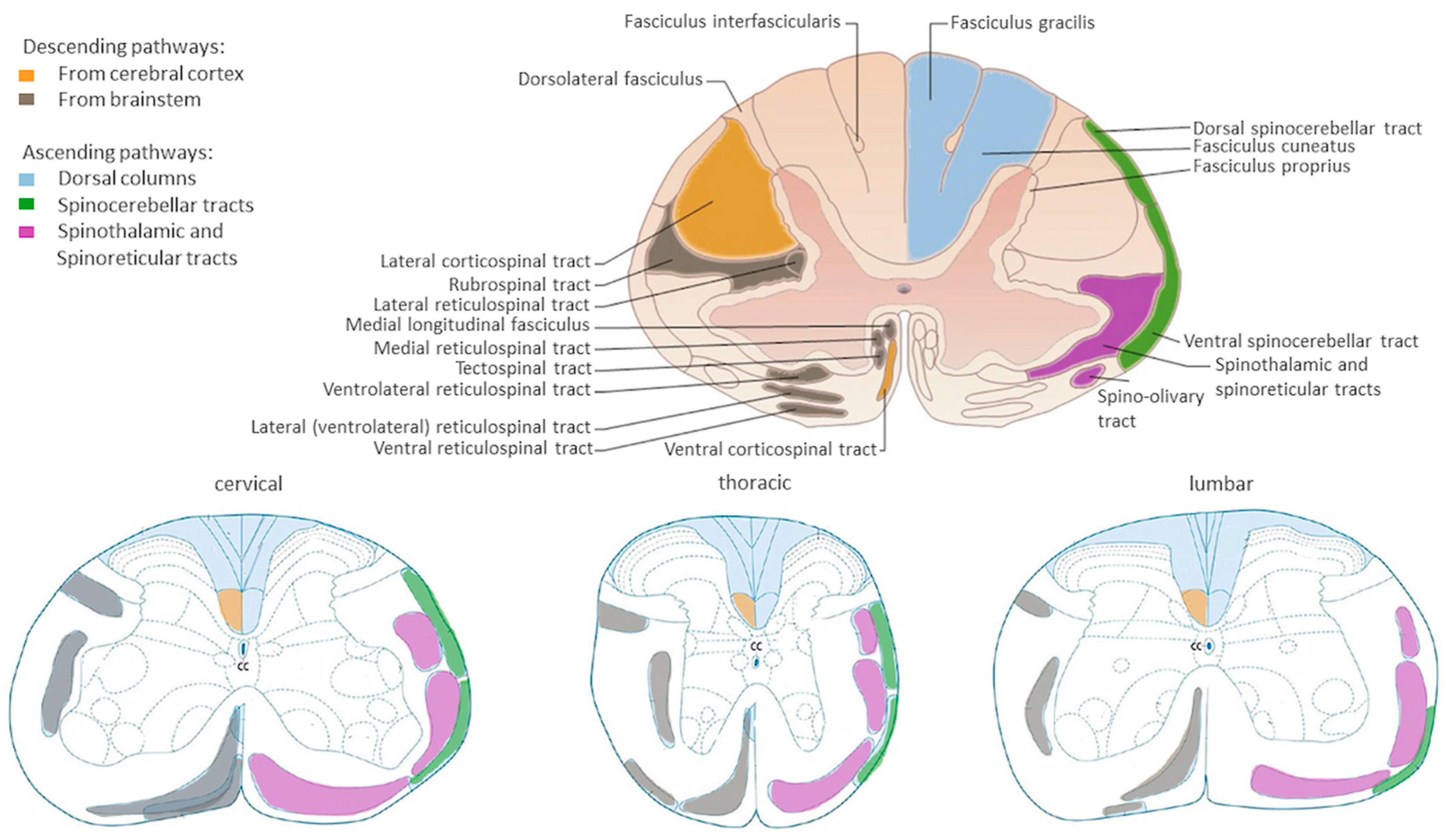
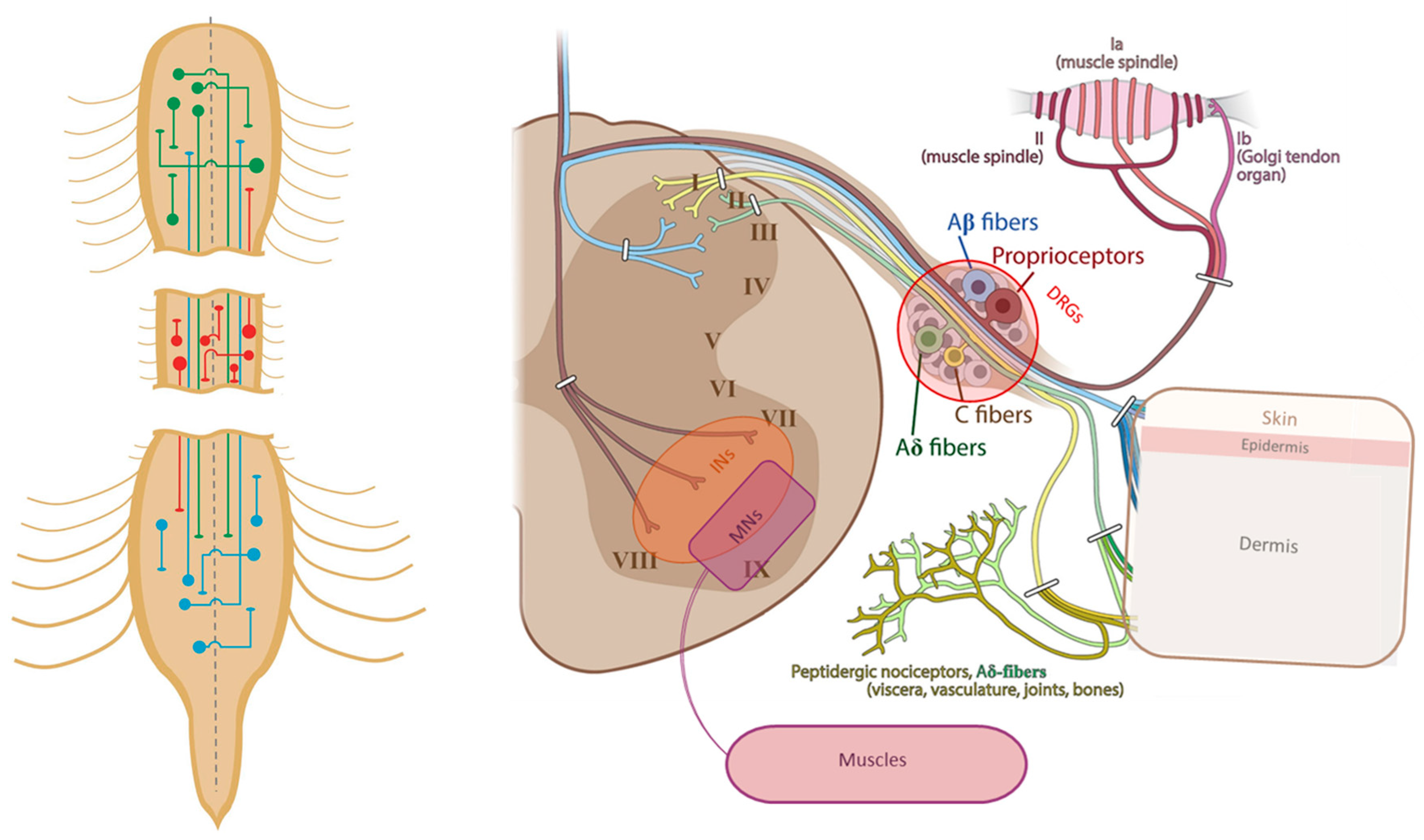
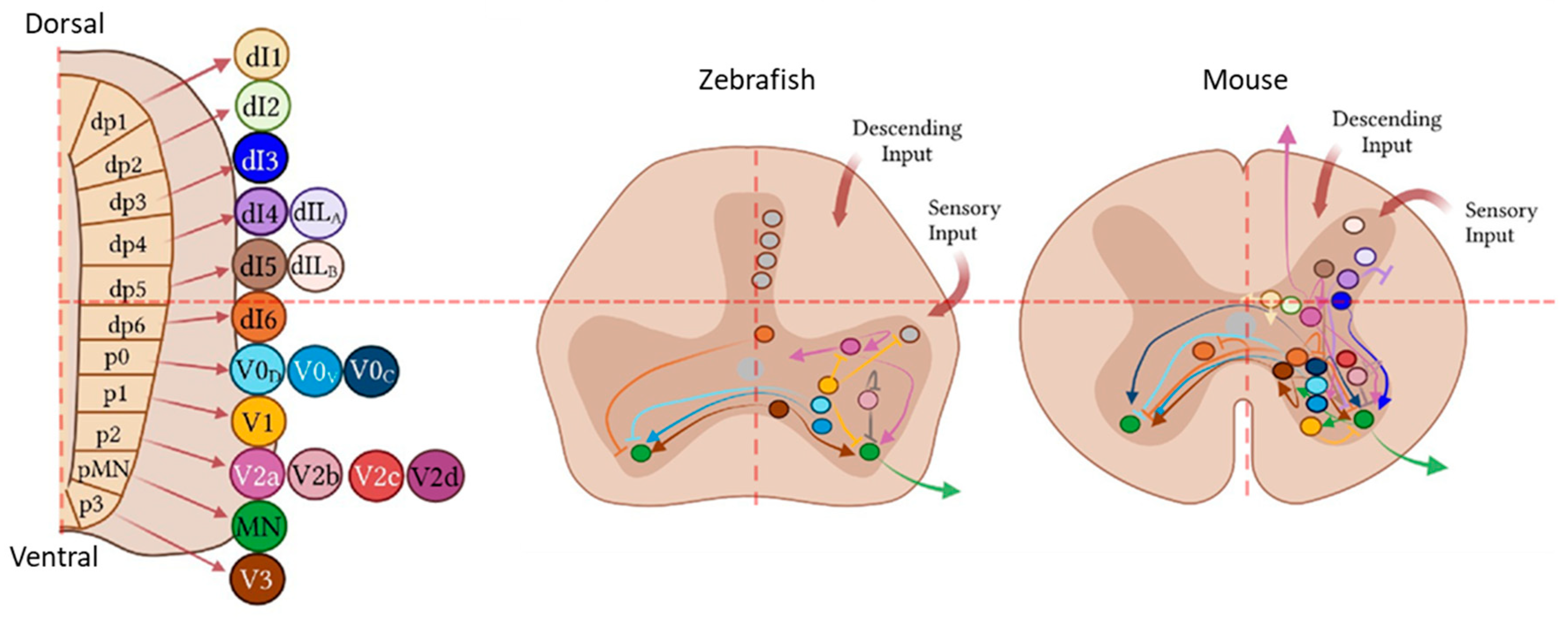
Potential Roles of Specific Subclasses of Premotor Interneurons in Spinal Cord Function Recovery after Traumatic Spinal Cord Injury in Adults


{kind=link}
{kind=link}
{kind=link}
Abstract
Share and Cite
Dominguez-Bajo, A.; Clotman, F. Potential Roles of Specific Subclasses of Premotor Interneurons in Spinal Cord Function Recovery after Traumatic Spinal Cord Injury in Adults. Cells 2024, 13, 652. https://doi.org/10.3390/cells13080652
Dominguez-Bajo A, Clotman F. Potential Roles of Specific Subclasses of Premotor Interneurons in Spinal Cord Function Recovery after Traumatic Spinal Cord Injury in Adults. Cells. 2024; 13(8):652. https://doi.org/10.3390/cells13080652
Chicago/Turabian StyleDominguez-Bajo, Ana, and Frédéric Clotman. 2024. "Potential Roles of Specific Subclasses of Premotor Interneurons in Spinal Cord Function Recovery after Traumatic Spinal Cord Injury in Adults" Cells 13, no. 8: 652. https://doi.org/10.3390/cells13080652
APA StyleDominguez-Bajo, A., & Clotman, F. (2024). Potential Roles of Specific Subclasses of Premotor Interneurons in Spinal Cord Function Recovery after Traumatic Spinal Cord Injury in Adults. Cells, 13(8), 652. https://doi.org/10.3390/cells13080652