
Sit4 Genetically Interacts with Vps27 to Regulate Mitochondrial Function and Lifespan in Saccharomyces cerevisiae

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Growth Conditions
2.2. Isolation of Vacuolar Membranes and Proteomic Analysis by HPLC-MS/MS
2.3. Chronological Lifespan
2.4. Glucose and Ethanol Determination
2.5. Glycogen Measurement
2.6. Western Blotting
2.7. Colony Immunoblot Assay
2.8. Iron Levels
2.9. Oxidative Stress Resistance
2.10. Growth Analysis
2.11. Oxygen Consumption
2.12. Alkaline Phosphatase Assay
2.13. Mitochondrial Mass
2.14. Analysis of Mitochondrial DNA by qPCR
2.15. Statistical Analysis
3. Results
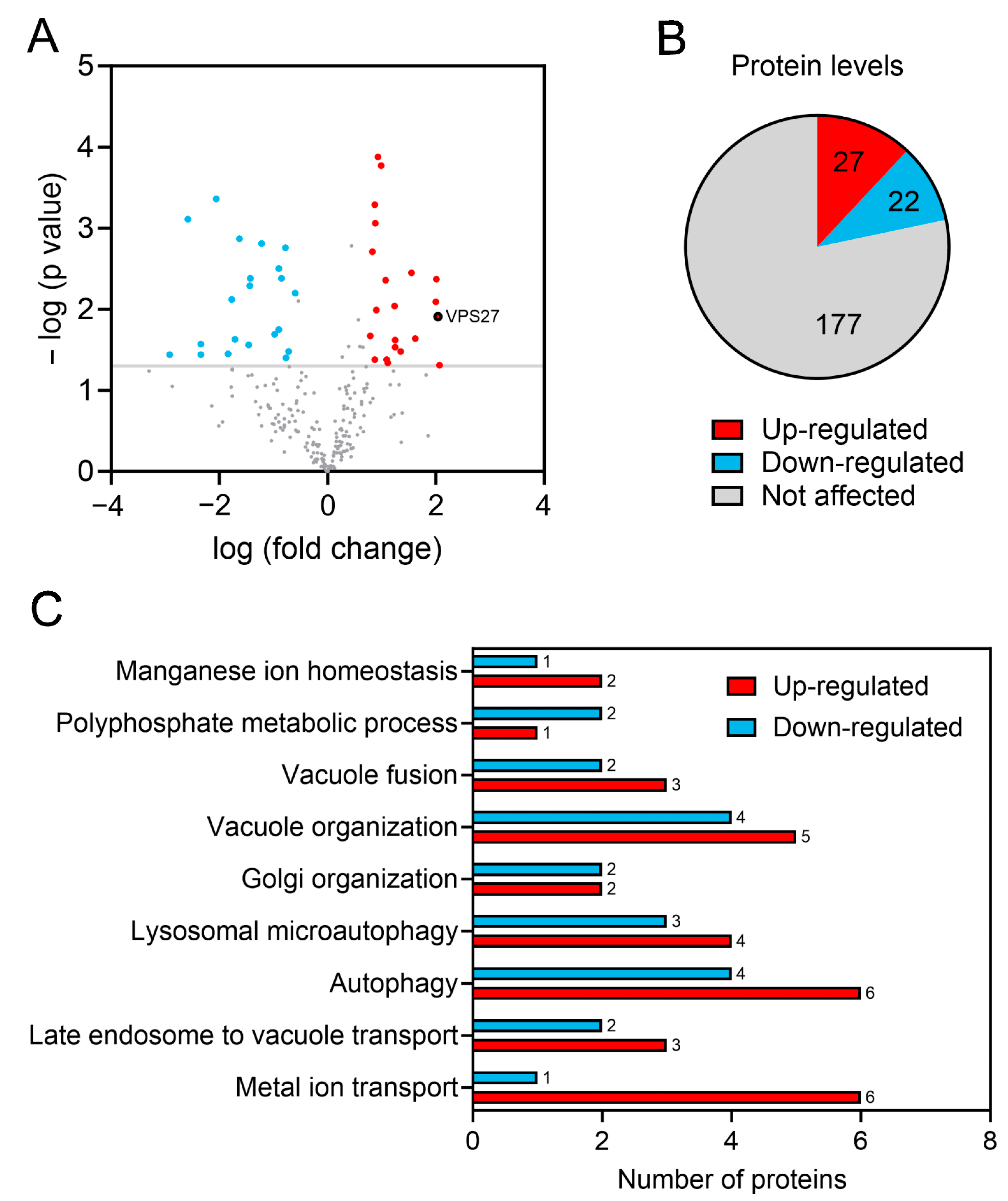
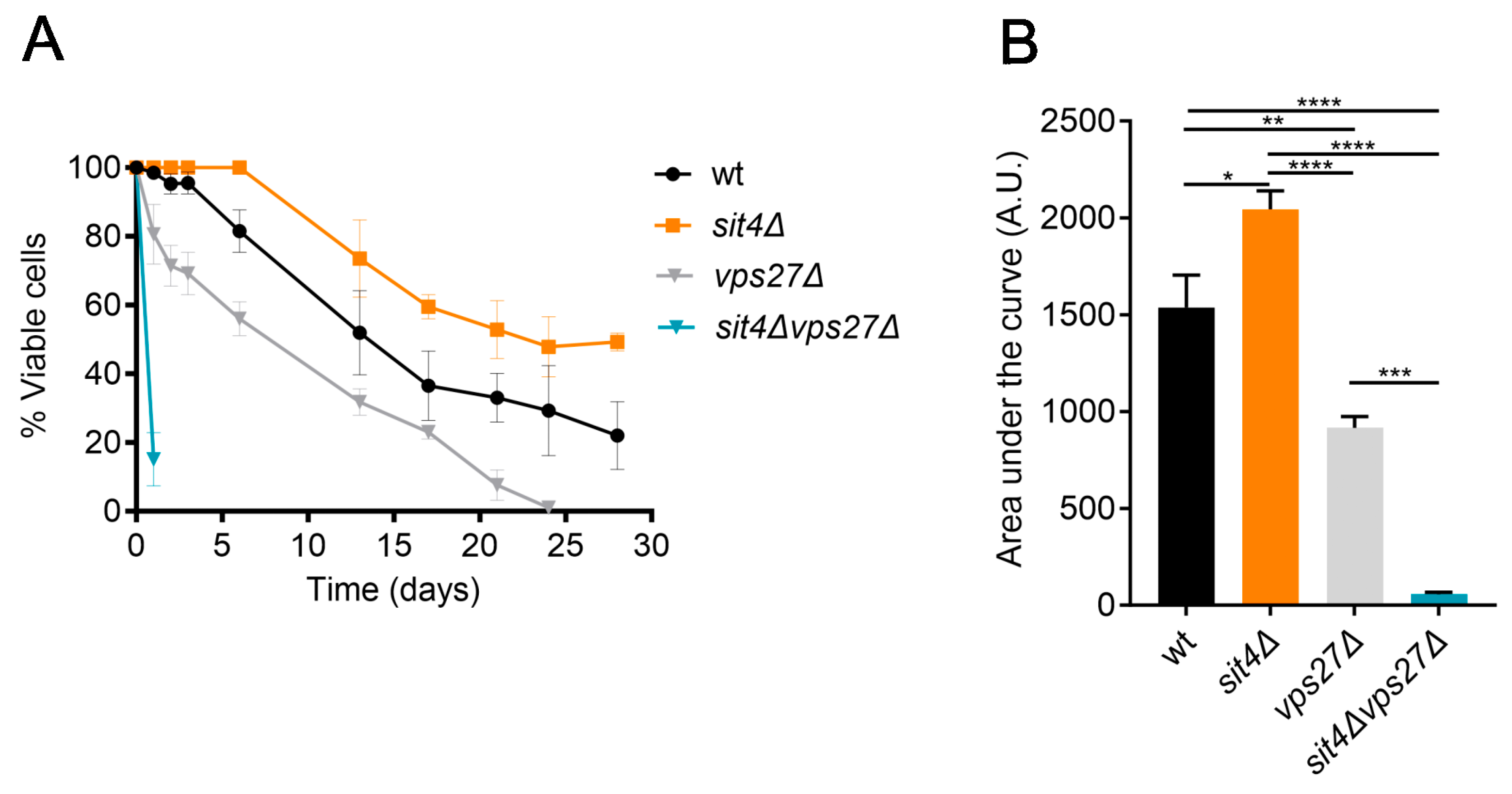
3.1. Vps27 Is Enriched in the Vacuolar Membranes and Mediates the Lifespan Extension of Cells Lacking the Phosphatase Sit4
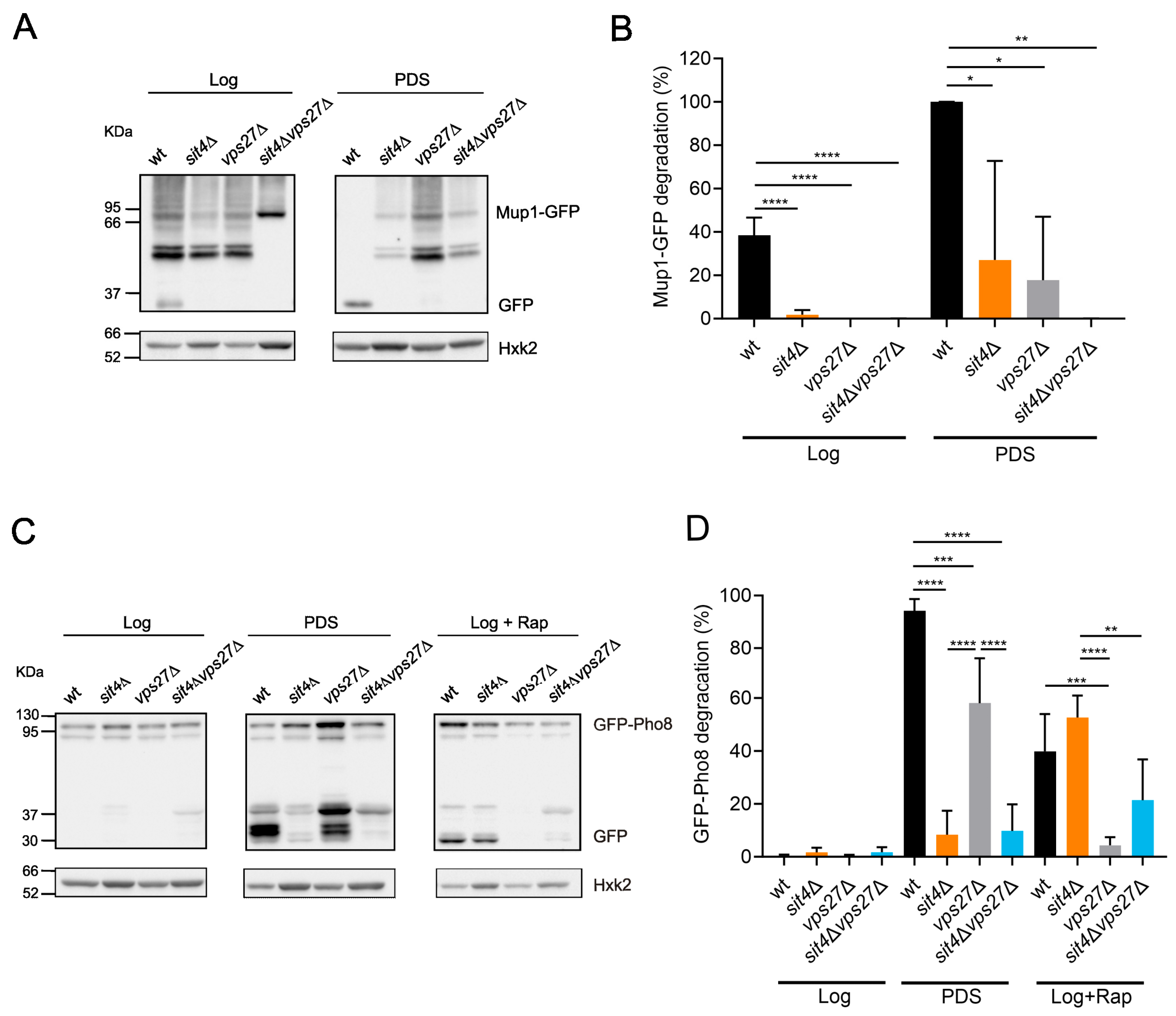
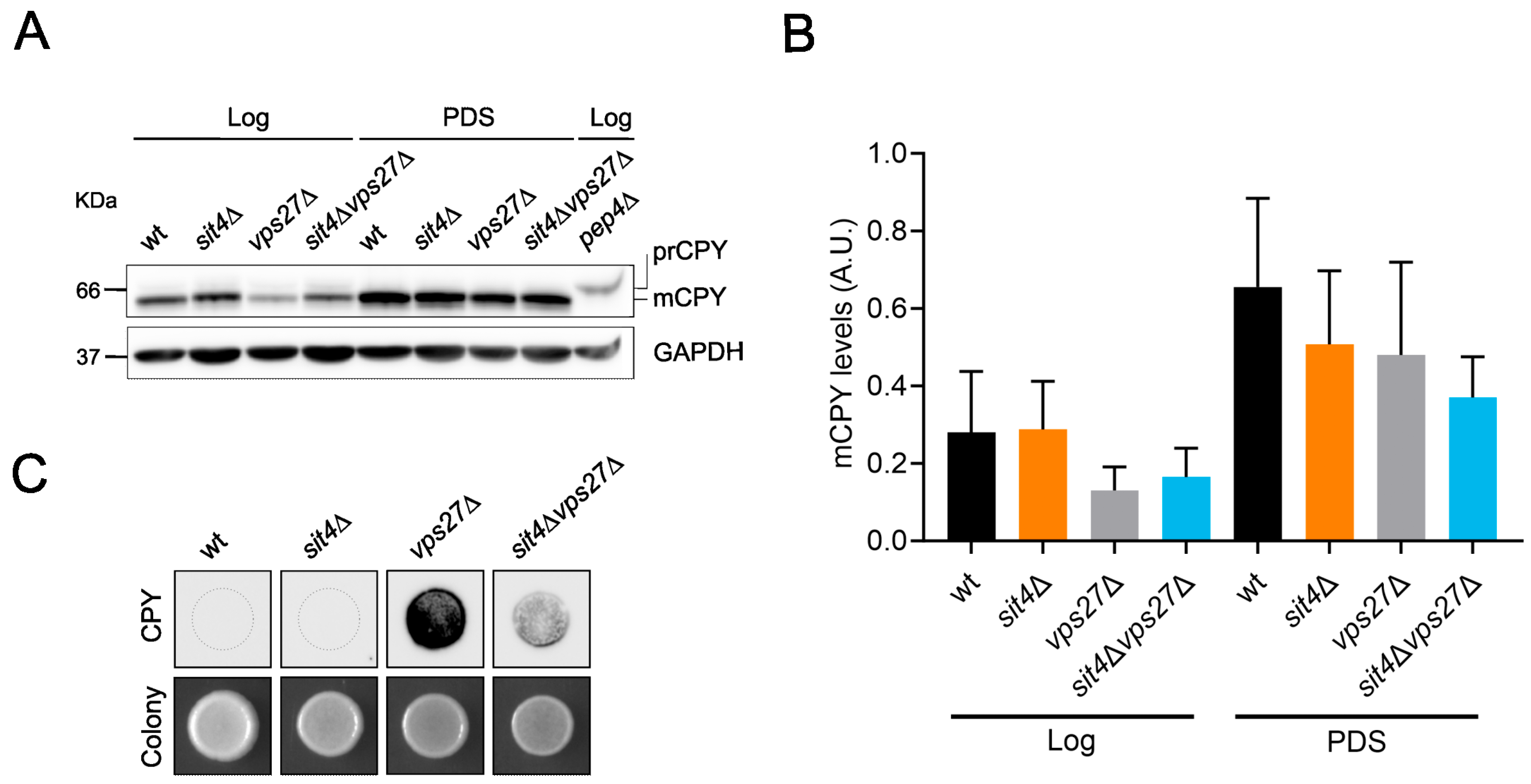
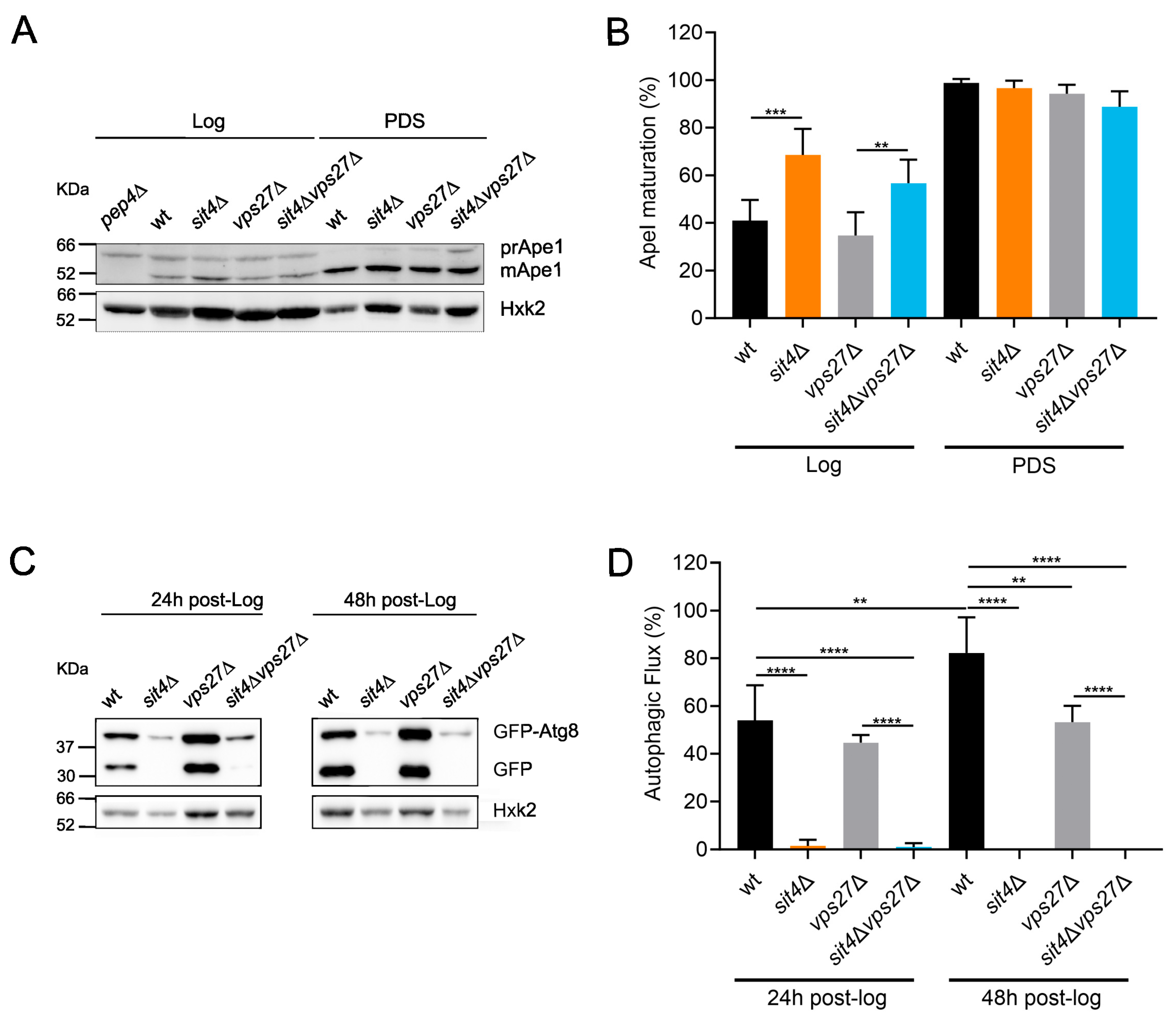
3.2. The Pathways for Vacuolar Degradation Are Downregulated in sit4Δ Cells in a Vps27-Independent Manner
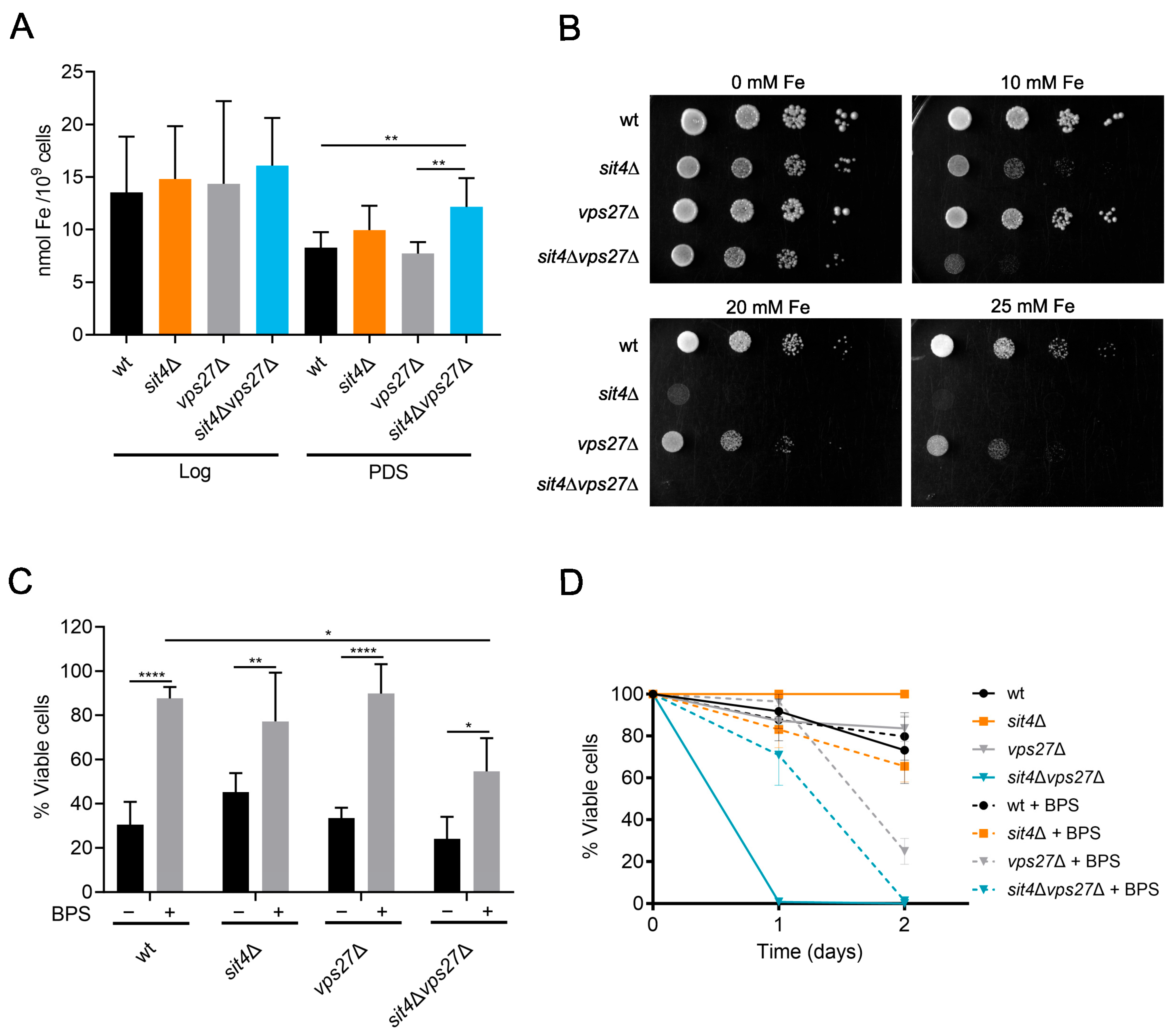
3.3. Loss of Iron Homeostasis in sit4∆vps27∆ Cells Does Not Contribute to Its Shortened Lifespan
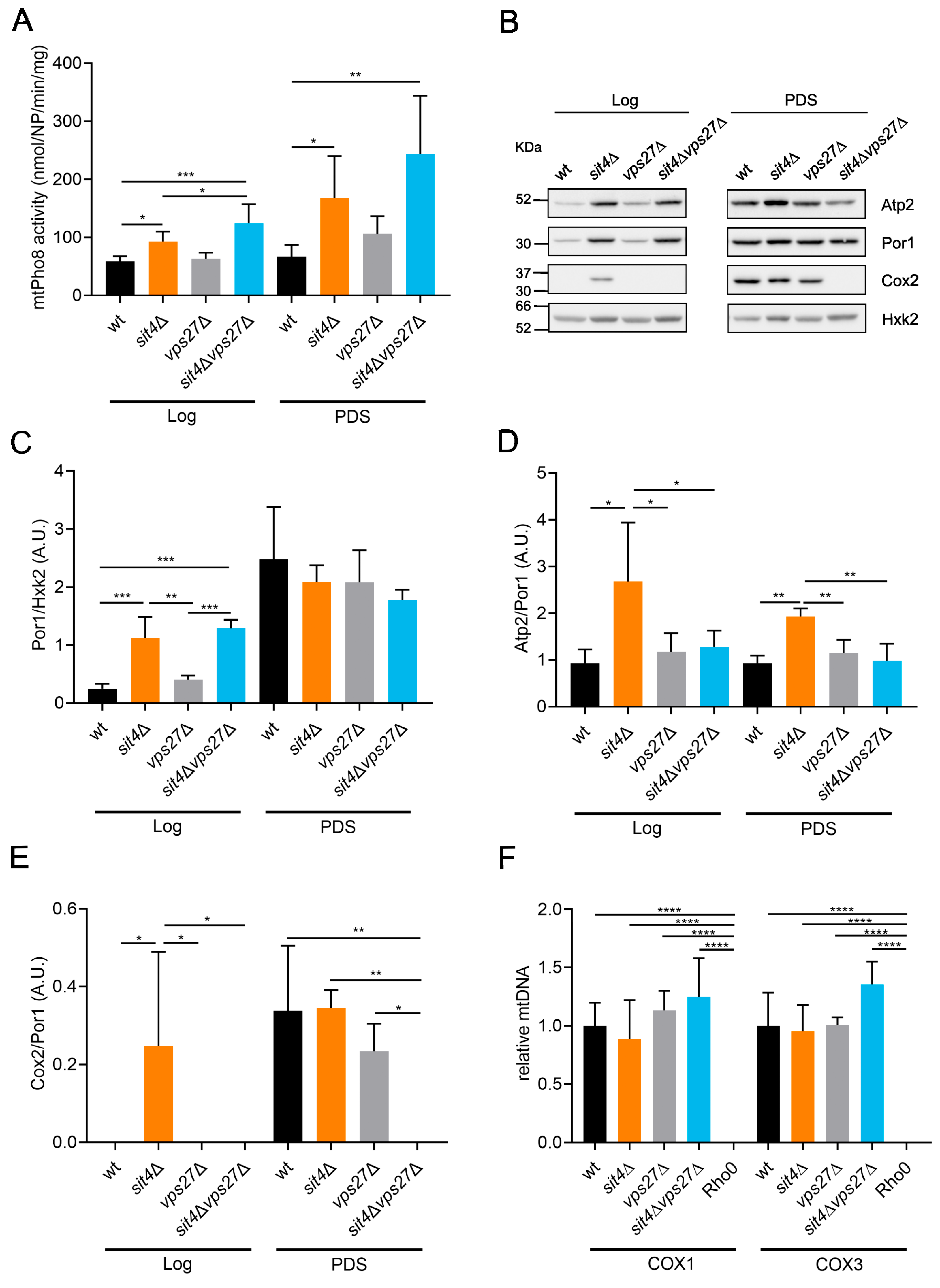
3.4. Vps27 Is Crucial for Mitochondrial Function in sit4∆ Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carmona-Gutierrez, D.; Hughes, A.L.; Madeo, F.; Ruckenstuhl, C. The Crucial Impact of Lysosomes in Aging and Longevity. Ageing Res. Rev. 2016, 32, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.-Y.; Zoncu, R. The Lysosome as a Command-and-Control Center for Cellular Metabolism. J. Cell Biol. 2016, 214, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Lamming, D.W.; Bar-Peled, L. Lysosome: The Metabolic Signaling Hub. Traffic 2019, 20, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, R.E.; Zoncu, R. The Lysosome as a Cellular Centre for Signalling, Metabolism and Quality Control. Nat. Cell Biol. 2019, 21, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Li, S.C.; Kane, P.M. The Yeast Lysosome-like Vacuole: Endpoint and Crossroads. Biochim. Biophys. Acta 2009, 1793, 650–663. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Evans, E.; Haslett, L.J. The Lysosomal Storage Disease Continuum with Ageing-Related Neurodegenerative Disease. Ageing Res. Rev. 2016, 32, 104–121. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Lopez, N.; Athonvarangkul, D.; Singh, R. Autophagy and Aging. Adv. Exp. Med. Biol. 2015, 847, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.L.; Gottschling, D.E. An Early Age Increase in Vacuolar pH Limits Mitochondrial Function and Lifespan in Yeast. Nature 2012, 492, 261–265. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Cuervo, A.M.; Dice, J.F. When Lysosomes Get Old. Exp. Gerontol. 2000, 35, 119–131. [Google Scholar] [CrossRef]
- Barbosa, M.C.; Grosso, R.A.; Fader, C.M. Hallmarks of Aging: An Autophagic Perspective. Front. Endocrinol. 2019, 9, 790. [Google Scholar] [CrossRef] [PubMed]
- Madeo, F.; Eisenberg, T.; Büttner, S.; Ruckenstuhl, C.; Kroemer, G. Spermidine: A Novel Autophagy Inducer and Longevity Elixir. Autophagy 2010, 6, 160–162. [Google Scholar] [CrossRef] [PubMed]
- Pyo, J.-O.; Yoo, S.-M.; Ahn, H.-H.; Nah, J.; Hong, S.-H.; Kam, T.-I.; Jung, S.; Jung, Y.-K. Overexpression of Atg5 in Mice Activates Autophagy and Extends Lifespan. Nat. Commun. 2013, 4, 2300. [Google Scholar] [CrossRef]
- Simonsen, A.; Cumming, R.C.; Brech, A.; Isakson, P.; Schubert, D.R.; Finley, K.D. Promoting Basal Levels of Autophagy in the Nervous System Enhances Longevity and Oxidant Resistance in Adult Drosophila. Autophagy 2008, 4, 176. [Google Scholar] [CrossRef] [PubMed]
- Tóth, M.L.; Sigmond, T.; Borsos, É.; Barna, J.; Erdélyi, P.; Takács-Vellai, K.; Orosz, L.; Kovács, A.L.; Csikós, G.; Sass, M.; et al. Longevity Pathways Converge on Autophagy Genes to Regulate Life Span in Caenorhabditis elegans. Autophagy 2008, 4, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Fabrizio, P.; Hu, J.; Ge, H.; Cheng, C.; Li, L.; Longo, V.D. Life Span Extension by Calorie Restriction Depends on Rim15 and Transcription Factors Downstream of Ras/PKA, Tor, and Sch9. PLoS Genet. 2008, 4, e13. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, H.; Suarez, J.A.; Longo, V.D. Protein and Amino Acid Restriction, Aging and Disease: From Yeast to Humans. Trends Endocrinol. Metab. 2014, 25, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Ruetenik, A.; Barrientos, A. Dietary Restriction, Mitochondrial Function and Aging: From Yeast to Humans. Biochim. Biophys. Acta—Bioenergy 2015, 1847, 1434–1447. [Google Scholar] [CrossRef]
- Hecht, K.A.; O’Donnell, A.F.; Brodsky, J.L. The Proteolytic Landscape of the Yeast Vacuole. Cell. Logist. 2014, 4, e28023. [Google Scholar] [CrossRef]
- Mehrpour, M.; Esclatine, A.; Beau, I.; Codogno, P. Overview of Macroautophagy Regulation in Mammalian Cells. Cell Res. 2010, 20, 748–762. [Google Scholar] [CrossRef]
- Wen, X.; Klionsky, D.J. An Overview of Macroautophagy in Yeast. J. Mol. Biol. 2016, 428, 1681–1699. [Google Scholar] [CrossRef] [PubMed]
- Youle, R.J.; Narendra, D.P. Mechanisms of Mitophagy. Nat. Rev. Mol. Cell Biol. 2011, 12, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Alao, J.P.; Legon, L.; Dabrowska, A.; Tricolici, A.M.; Kumar, J.; Rallis, C. Interplays of AMPK and TOR in Autophagy Regulation in Yeast. Cells 2023, 12, 519. [Google Scholar] [CrossRef] [PubMed]
- Aman, Y.; Schmauck-Medina, T.; Hansen, M.; Morimoto, R.I.; Simon, A.K.; Bjedov, I.; Palikaras, K.; Simonsen, A.; Johansen, T.; Tavernarakis, N.; et al. Autophagy in Healthy Aging and Disease. Nat. Aging 2021, 1, 634–650. [Google Scholar] [CrossRef] [PubMed]
- Luzio, J.P.; Parkinson, M.D.J.; Gray, S.R.; Bright, N.A. The Delivery of Endocytosed Cargo to Lysosomes. Biochem. Soc. Trans. 2009, 37, 1019–1021. [Google Scholar] [CrossRef] [PubMed]
- Feyder, S.; De Craene, J.-O.; Bär, S.; Bertazzi, D.; Friant, S. Membrane Trafficking in the Yeast Saccharomyces cerevisiae Model. Int. J. Mol. Sci. 2015, 16, 1509–1525. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Burd, C.G. Retrograde Trafficking and Plasma Membrane Recycling Pathways of the Budding Yeast Saccharomyces cerevisiae. Traffic 2020, 21, 45–59. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, M.J.; Lindsay, A.J. The Endosomal Recycling Pathway—At the Crossroads of the Cell. Int. J. Mol. Sci. 2020, 21, 6074. [Google Scholar] [CrossRef] [PubMed]
- Bowers, K.; Stevens, T.H. Protein Transport from the Late Golgi to the Vacuole in the Yeast Saccharomyces cerevisiae. Biochim. Biophys. Acta—Mol. Cell Res. 2005, 1744, 438–454. [Google Scholar] [CrossRef]
- Lynch-Day, M.A.; Klionsky, D.J. The Cvt Pathway as a Model for Selective Autophagy. FEBS Lett. 2010, 584, 1359–1366. [Google Scholar] [CrossRef]
- Galaris, D.; Barbouti, A.; Pantopoulos, K. Iron Homeostasis and Oxidative Stress: An Intimate Relationship. Biochim. Biophys. Acta—Mol. Cell Res. 2019, 1866, 118535. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, A.D.; Osório, H.; Sims, K.J.; Almeida, T.; Alves, M.; Bielawski, J.; Amorim, M.A.; Moradas-Ferreira, P.; Hannun, Y.A.; Costa, V. Role for Sit4p-Dependent Mitochondrial Dysfunction in Mediating the Shortened Chronological Lifespan and Oxidative Stress Sensitivity of Isc1p-Deficient Cells. Mol. Microbiol. 2011, 81, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, V.; Medeiros, T.C.; Vilaça, R.; Ferreira, J.; Moradas-Ferreira, P.; Costa, V. Ceramide Signaling Targets the PP2A-like Protein Phosphatase Sit4p to Impair Vacuolar Function, Vesicular Trafficking and Autophagy in Isc1p Deficient Cells. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2016, 1861, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Pereira, A.T.; Osório, H.; Moradas-Ferreira, P.; Costa, V. Sit4p-Mediated Dephosphorylation of Atp2p Regulates ATP Synthase Activity and Mitochondrial Function. Biochim. Biophys. Acta—Bioenergy 2018, 1859, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.; Pereira, A.T.; Costa, V. Activation of SNF1/AMPK Mediates the Mitochondrial Derepression, Resistance to Oxidative Stress and Increased Lifespan of Cells Lacking the Phosphatase Sit4p. Biochim. Biophys. Acta—Mol. Cell Res. 2020, 1867, 118660. [Google Scholar] [CrossRef] [PubMed]
- Bastians, H.; Ponstingl, H. The Novel Human Protein Serine/Threonine Phosphatase 6 Is a Functional Homologue of Budding Yeast Sit4p and Fission Yeast Ppe1, Which Are Involved in Cell Cycle Regulation. J. Cell Sci. 1996, 109, 2865–2874. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Sarabia, M.J.; Sutton, A.; Zhong, T.; Arndt, K.T. SIT4 Protein Phosphatase Is Required for the Normal Accumulation of SWI4, CLN1, CLN2, and HCS26 RNAs during Late G1. Genes Dev. 1992, 6, 2417–2428. [Google Scholar] [CrossRef] [PubMed]
- Nickels, J.T.; Broach, J.R. A Ceramide-Activated Protein Phosphatase Mediates Ceramide-Induced G1 Arrest of Saccharomyces cerevisiae. Genes Dev. 1996, 10, 382–394. [Google Scholar] [CrossRef] [PubMed]
- De La Torre-Ruiz, M.A.; Torres, J.; Ariño, J.; Herrero, E. Sit4 Is Required for Proper Modulation of the Biological Functions Mediated by Pkc1 and the Cell Integrity Pathway in Saccharomyces cerevisiae. J. Biol. Chem. 2002, 277, 33468–33476. [Google Scholar] [CrossRef]
- Di Como, C.J.; Arndt, K.T. Nutrients, via the Tor Proteins, Stimulate the Association of Tap42 with Type 2A Phosphatases. Genes Dev. 1996, 10, 1904–1916. [Google Scholar] [CrossRef]
- Zhang, W.; Du, G.; Zhou, J.; Chen, J. Regulation of Sensing, Transportation, and Catabolism of Nitrogen Sources in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2018, 82, e00040-17. [Google Scholar] [CrossRef] [PubMed]
- Jablonka, W.; Guzmán, S.; Ramírez, J.; Montero-Lomelí, M. Deviation of Carbohydrate Metabolism by the SIT4 Phosphatase in Saccharomyces cerevisiae. Biochim. Biophys. Acta—Gen. Subj. 2006, 1760, 1281–1291. [Google Scholar] [CrossRef] [PubMed]
- Bozaquel-Morais, B.L.; Madeira, J.B.; Maya-Monteiro, C.M.; Masuda, C.A.; Montero-Lomeli, M. A New Fluorescence-Based Method Identifies Protein Phosphatases Regulating Lipid Droplet Metabolism. PLoS ONE 2010, 5, e13692. [Google Scholar] [CrossRef] [PubMed]
- Masuda, C.A.; Ramírez, J.; Peña, A.; Montero-Lomelí, M. Regulation of Monovalent Ion Homeostasis and pH by the Ser-Thr Protein Phosphatase SIT4 in Saccharomyces cerevisiae. J. Biol. Chem. 2000, 275, 30957–30961. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, D.; Zhang, J.; Menon, S.; Lord, C.; Chen, S.; Helm, J.R.; Thorsen, K.; Corbett, K.D.; Hay, J.C.; Ferro-Novick, S. Sit4p/PP6 Regulates ER-to-Golgi Traffic by Controlling the Dephosphorylation of COPII Coat Subunits. Mol. Biol. Cell 2013, 24, 2727–2738. [Google Scholar] [CrossRef] [PubMed]
- Luke, M.M.; Della Seta, F.; Di Como, C.J.; Sugimoto, H.; Kobayashi, R.; Arndt, K.T. The SAPs, a New Family of Proteins, Associate and Function Positively with the SIT4 Phosphatase. Mol. Cell. Biol. 1996, 16, 2744–2755. [Google Scholar] [CrossRef] [PubMed]
- Woodacre, A.; Lone, M.A.; Jablonowski, D.; Schneiter, R.; Giorgini, F.; Schaffrath, R. A Novel Sit4 Phosphatase Complex Is Involved in the Response to Ceramide Stress in Yeast. Oxid. Med. Cell. Longev. 2013, 2013, 129645. [Google Scholar] [CrossRef] [PubMed]
- Tate, J.J.; Tolley, E.A.; Cooper, T.G. Sit4 and PP2A Dephosphorylate Nitrogen Catabolite Repression-Sensitive Gln3 When TorC1 Is up- as Well as Downregulated. Genetics 2019, 212, 1205–1225. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, V.; Medeiros, T.C.; Vilaça, R.; Pereira, A.T.; Chaves, S.R.; Côrte-Real, M.; Moradas-Ferreira, P.; Costa, V. Ceramide Signalling Impinges on Sit4p and Hog1p to Promote Mitochondrial Fission and Mitophagy in Isc1p-Deficient Cells. Cell Signal. 2015, 27, 1840–1849. [Google Scholar] [CrossRef]
- Barbosa, A.D.; Graça, J.; Mendes, V.; Chaves, S.R.; Amorim, M.A.; Mendes, M.V.; Moradas-Ferreira, P.; Côrte-Real, M.; Costa, V. Activation of the Hog1p Kinase in Isc1p-Deficient Yeast Cells Is Associated with Mitochondrial Dysfunction, Oxidative Stress Sensitivity and Premature Aging. Mech. Ageing Dev. 2012, 133, 317–330. [Google Scholar] [CrossRef]
- Gietz, R.D.; Schiestl, R.H. High-Efficiency Yeast Transformation Using the LiAc/SS Carrier DNA/PEG Method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Wiederhold, E.; Gandhi, T.; Permentier, H.P.; Breitling, R.; Poolman, B.; Slotboom, D.J. The Yeast Vacuolar Membrane Proteome. Mol. Cell Proteom. 2009, 8, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Martins, T.S.; Costa, R.S.; Vilaça, R.; Lemos, C.; Teixeira, V.; Pereira, C.; Costa, V. Iron Limitation Restores Autophagy and Increases Lifespan in the Yeast Model of Niemann–Pick Type C1. Int. J. Mol. Sci. 2023, 24, 6221. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.S.; Moggridge, S.; Müller, T.; Sorensen, P.H.; Morin, G.B.; Krijgsveld, J. Single-Pot, Solid-Phase-Enhanced Sample Preparation for Proteomics Experiments. Nat. Protoc. 2019, 14, 68–85. [Google Scholar] [CrossRef] [PubMed]
- Osório, H.; Silva, C.; Ferreira, M.; Gullo, I.; Máximo, V.; Barros, R.; Mendonça, F.; Oliveira, C.; Carneiro, F. Proteomics Analysis of Gastric Cancer Patients with Diabetes Mellitus. J. Clin. Med. 2021, 10, 407. [Google Scholar] [CrossRef] [PubMed]
- Fabrizio, P.; Longo, V.D. The Chronological Life Span of Saccharomyces cerevisiae. Aging Cell 2003, 2, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Chun, K.T.; Wek, R.C. Mitochondrial Respiratory Mutants in Yeast Inhibit Glycogen Accumulation by Blocking Activation of Glycogen Synthase. J. Biol. Chem. 1998, 273, 31337–31344. [Google Scholar] [CrossRef]
- Golden, C.K.; Kazmirchuk, T.D.D.; McNally, E.K.; El Eissawi, M.; Gokbayrak, Z.D.; Richard, J.D.; Brett, C.L. A Two-Tiered System for Selective Receptor and Transporter Protein Degradation. PLoS Genet. 2022, 18, e1010446. [Google Scholar] [CrossRef]
- Martins, T.S.; Pereira, C.; Canadell, D.; Vilaça, R.; Teixeira, V.; Moradas-Ferreira, P.; de Nadal, E.; Posas, F.; Costa, V. The Hog1p Kinase Regulates Aft1p Transcription Factor to Control Iron Accumulation. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2018, 1863, 61–70. [Google Scholar] [CrossRef]
- Vowinckel, J.; Hartl, J.; Butler, R.; Ralser, M. MitoLoc: A Method for the Simultaneous Quantification of Mitochondrial Network Morphology and Membrane Potential in Single Cells. Mitochondrion 2015, 24, 77–86. [Google Scholar] [CrossRef]
- Westermann, B.; Neupert, W. Mitochondria-Targeted Green Fluorescent Proteins: Convenient Tools for the Study of Organelle Biogenesis in Saccharomyces cerevisiae. Yeast 2000, 16, 1421–1427. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hochstrasser, M. Selective Microautophagy of Proteasomes Is Initiated by ESCRT-0 and Is Promoted by Proteasome Ubiquitylation. J. Cell Sci. 2022, 135, jcs259393. [Google Scholar] [CrossRef] [PubMed]
- Oku, M.; Maeda, Y.; Kagohashi, Y.; Kondo, T.; Yamada, M.; Fujimoto, T.; Sakai, Y. Evidence for ESCRT- and Clathrin-Dependent Microautophagy. J. Cell Biol. 2017, 216, 3263–3274. [Google Scholar] [CrossRef] [PubMed]
- Bilodeau, P.S.; Urbanowski, J.L.; Winistorfer, S.C.; Piper, R.C. The Vps27p-Hse1p Complex Binds Ubiquitin and Mediates Endosomal Protein Sorting. Nat. Cell Biol. 2002, 4, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Katzmann, D.J.; Stefan, C.J.; Babst, M.; Emr, S.D. Vps27 Recruits ESCRT Machinery to Endosomes during MVB Sorting. J. Cell Biol. 2003, 162, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Marek, A.; Korona, R. Restricted Pleiotropy Facilitates Mutational Erosion of Major Life-History Traits. Evolution 2013, 67, 3077–3086. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Tang, Y.; Quan, Z.; Zhang, Z.; Oliver, S.G.; Zhang, N. Chronological Lifespan in Yeast Is Dependent on the Accumulation of Storage Carbohydrates Mediated by Yak1, Mck1 and Rim15 Kinases. PLoS Genet. 2016, 12, e1006458. [Google Scholar] [CrossRef]
- Müller, M.; Schmidt, O.; Angelova, M.; Faserl, K.; Weys, S.; Kremser, L.; Pfaffenwimmer, T.; Dalik, T.; Kraft, C.; Trajanoski, Z.; et al. The Coordinated Action of the MVB Pathway and Autophagy Ensures Cell Survival during Starvation. eLife 2015, 4, e07736. [Google Scholar] [CrossRef]
- Ebrahimi, M.; Habernig, L.; Broeskamp, F.; Aufschnaiter, A.; Diessl, J.; Atienza, I.; Matz, S.; Ruiz, F.A.; Büttner, S. Phosphate Restriction Promotes Longevity via Activation of Autophagy and the Multivesicular Body Pathway. Cells 2021, 10, 3161. [Google Scholar] [CrossRef]
- Ivashov, V.; Zimmer, J.; Schwabl, S.; Kahlhofer, J.; Weys, S.; Gstir, R.; Jakschitz, T.; Kremser, L.; Bonn, G.K.; Lindner, H.; et al. Complementary A-Arrestin-Ubiquitin Ligase Complexes Control Nutrient Transporter Endocytosis in Response to Amino Acids. eLife 2020, 9, e58246. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, R.; De Virgilio, C. TORC1 Specifically Inhibits Microautophagy through ESCRT-0. Curr. Genet. 2019, 65, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Morshed, S.; Tasnin, M.N.; Ushimaru, T. ESCRT Machinery Plays a Role in Microautophagy in Yeast. BMC Mol. Cell Biol. 2020, 21, 70. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, R.; Péli-Gulli, M.P.; Hu, Z.; Jaquenoud, M.; Garcia Osuna, G.M.; Sardu, A.; Dengjel, J.; De Virgilio, C. Spatially Distinct Pools of TORC1 Balance Protein Homeostasis. Mol. Cell 2019, 73, 325–338.e8. [Google Scholar] [CrossRef]
- Bonangelino, C.J.; Chavez, E.M.; Bonifacino, J.S. Genomic Screen for Vacuolar Protein Sorting Genes in Saccharomyces cerevisiae. Mol. Biol. Cell 2002, 13, 2486–2501. [Google Scholar] [CrossRef] [PubMed]
- Piper, R.C.; Cooper, A.A.; Yang, H.; Stevens, T.H. VPS27 Controls Vacuolar and Endocytic Traffic through a Prevacuolar Compartment in Saccharomyces cerevisiae. J. Cell Biol. 1995, 131, 603–617. [Google Scholar] [CrossRef] [PubMed]
- Raymond, C.K.; Howald-Stevenson, I.; Vater, C.A.; Stevens, T.H. Morphological Classification of the Yeast Vacuolar Protein Sorting Mutants: Evidence for a Prevacuolar Compartment in Class E Vps Mutants. Mol. Biol. Cell 1992, 3, 1389–1402. [Google Scholar] [CrossRef]
- Liang, Y. Phagophore-Lysosome/Vacuole Fusion in Mutant Yeast and Mammalian Cells. Autophagy 2023, 19, 2595–2600. [Google Scholar] [CrossRef]
- Sun, L.X.; Qian, H.; Liu, M.Y.; Wu, M.H.; Wei, Y.Y.; Zhu, X.M.; Lu, J.P.; Lin, F.C.; Liu, X.H. Endosomal Sorting Complexes Required for Transport-0 (ESCRT-0) Are Essential for Fungal Development, Pathogenicity, Autophagy and ER-Phagy in Magnaporthe oryzae. Environ. Microbiol. 2022, 24, 1076–1092. [Google Scholar] [CrossRef]
- Hansen, M.; Rubinsztein, D.C.; Walker, D.W. Autophagy as a Promoter of Longevity: Insights from Model Organisms. Nat. Rev. Mol. Cell Biol. 2018, 19, 579–593. [Google Scholar] [CrossRef]
- Nair, U.; Thumm, M.; Klionsky, D.J.; Krick, R. GFP-Atg8 Protease Protection as a Tool to Monitor Autophagosome Biogenesis. Autophagy 2011, 7, 1546–1550. [Google Scholar] [CrossRef] [PubMed]
- Yorimitsu, T.; He, C.; Wang, K.; Klionsky, D.J. Tap42-Associated Protein Phosphatase Type 2A Negatively Regulates Induction of Autophagy. Autophagy 2009, 5, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Strochlic, T.I.; Schmiedekamp, B.C.; Lee, J.; Katzmann, D.J.; Burd, C.G. Opposing Activities of the Snx3-Retromer Complex and ESCRT Proteins Mediate Regulated Cargo Sorting at a Common Endosome. Mol. Biol. Cell 2008, 19, 4694–4706. [Google Scholar] [CrossRef] [PubMed]
- Ahuatzi, D.; Riera, A.; Peláez, R.; Herrero, P.; Moreno, F. Hxk2 Regulates the Phosphorylation State of Mig1 and Therefore Its Nucleocytoplasmic Distribution. J. Biol. Chem. 2007, 282, 4485–4493. [Google Scholar] [CrossRef] [PubMed]
- Moreno, F.; Herrero, P. The Hexokinase 2-Dependent Glucose Signal Transduction Pathway of Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2002, 26, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, A.D.; Pereira, C.; Osório, H.; Moradas-Ferreira, P.; Costa, V. The Ceramide-Activated Protein Phosphatase Sit4p Controls Lifespan, Mitochondrial Function and Cell Cycle Progression by Regulating Hexokinase 2 Phosphorylation. Cell Cycle 2016, 15, 1620–1630. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Barrientos, A.; Epstein, C.B.; Butow, R.A.; Tzagoloff, A. SIT4 Regulation of Mig1p-Mediated Catabolite Repression in Saccharomyces cerevisiae. FEBS Lett. 2007, 581, 5658–5663. [Google Scholar] [CrossRef]
- Ruiz, A.; Xu, X.; Carlson, M. Roles of Two Protein Phosphatases, Reg1-Glc7 and Sit4, and Glycogen Synthesis in Regulation of SNF1 Protein Kinase. Proc. Natl. Acad. Sci. USA 2011, 108, 6349–6354. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, A.; Liu, Y.; Xu, X.; Carlson, M. Heterotrimer-Independent Regulation of Activation-Loop Phosphorylation of Snf1 Protein Kinase Involves Two Protein Phosphatases. Proc. Natl. Acad. Sci. USA 2012, 109, 8652–8657. [Google Scholar] [CrossRef]
- Treitel, M.A.; Kuchin, S.; Carlson, M. Snf1 Protein Kinase Regulates Phosphorylation of the Mig1 Repressor in Saccharomyces cerevisiae. Mol. Cell. Biol. 1998, 18, 6273–6280. [Google Scholar] [CrossRef]
- Barros, M.H.; Bandy, B.; Tahara, E.B.; Kowaltowski, A.J. Higher Respiratory Activity Decreases Mitochondrial Reactive Oxygen Release and Increases Life Span in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 49883–49888. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Schroeder, E.A.; Ocampo, A.; Barrientos, A.; Shadel, G.S. Regulation of Yeast Chronological Life Span by TORC1 via Adaptive Mitochondrial ROS Signaling. Cell Metab. 2011, 13, 668–678. [Google Scholar] [CrossRef] [PubMed]
- Ng, M.Y.W.; Wai, T.; Simonsen, A. Quality Control of the Mitochondrion. Dev. Cell 2021, 56, 881–905. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Chiang, W. Mitophagy in Aging and Longevity. IUBMB Life 2022, 74, 296–316. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, R.A.; Bernstein, D. Mitochondrial Remodeling: Rearranging, Recycling, and Reprogramming. Cell Calcium 2016, 60, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Innokentev, A.; Kanki, T. Mitophagy in Yeast: Molecular Mechanism and Regulation. Cells 2021, 10, 3569. [Google Scholar] [CrossRef]
- Campbell, C.L.; Thorsness, P.E. Escape of Mitochondrial DNA to the Nucleus in Yme1 Yeast Is Mediated by Vacuolar-Dependent Turnover of Abnormal Mitochondrial Compartments. J. Cell Sci. 1998, 111, 2455–2464. [Google Scholar] [CrossRef]
- Seo, A.Y.; Joseph, A.-M.; Dutta, D.; Hwang, J.C.Y.; Aris, J.P.; Leeuwenburgh, C. New Insights into the Role of Mitochondria in Aging: Mitochondrial Dynamics and More. J. Cell Sci. 2010, 123, 2533–2542. [Google Scholar] [CrossRef] [PubMed]
- Madeo, F.; Zimmermann, A.; Maiuri, M.C.; Kroemer, G. Essential Role for Autophagy in Life Span Extension. J. Clin. Investig. 2015, 125, 85–93. [Google Scholar] [CrossRef]
- Tang, F.; Watkins, J.W.; Bermudez, M.; Gray, R.; Gaban, A.; Portie, K.; Grace, S.; Kleve, M.; Craciun, G. A Life-Span Extending Form of Autophagy Employs the Vacuole-Vacuole Fusion Machinery. Autophagy 2008, 4, 874–886. [Google Scholar] [CrossRef]
- Athané, A.; Buisson, A.; Challier, M.; Beaumatin, F.; Manon, S.; Bhatia-Kiššová, I.; Camougrand, N. Insights into the Relationship between the Proteasome and Autophagy in Human and Yeast Cells. Int. J. Biochem. Cell Biol. 2015, 64, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Nam, T.; Han, J.H.; Devkota, S.; Lee, H.-W. Emerging Paradigm of Crosstalk between Autophagy and the Ubiquitin-Proteasome System. Mol. Cells 2017, 40, 897–905. [Google Scholar] [CrossRef]
- Ji, C.H.; Kwon, Y.T. Crosstalk and Interplay between the Ubiquitin-Proteasome System and Autophagy. Mol. Cells 2017, 40, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Dikic, I. Proteasomal and Autophagic Degradation Systems. Annu. Rev. Biochem. 2017, 86, 193–224. [Google Scholar] [CrossRef] [PubMed]
- Singer, T.; Haefner, S.; Hoffmann, M.; Fischer, M.; Ilyina, J.; Hilt, W. Sit4 Phosphatase Is Functionally Linked to the Ubiquitin-Proteasome System. Genetics 2003, 164, 1305–1321. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE Database Resources in 2022: A Hub for Mass Spectrometry-Based Proteomics Evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Source |
|---|---|---|
| BY4741 b,e,f | Mata, his3Δ1, leu2Δ0, met15Δ0, ura3Δ0 | EUROSCARF |
| sit4Δ b,e,f | BY4741 sit4::MXHIS3 | [34] |
| vps27∆ a,b,e,f | BY4741 vps27::KanMX4 | This study |
| sit4∆vps27∆ a,b,e,f | BY4741 sit4::MXHIS3 vps27::KanMX4 | This study |
| pep4Δ | BY4741 pep4::KanMX4 | EUROSCARF |
| pho8Δ c | BY4741 pho8::HPH | [49] |
| sit4Δpho8Δ c | BY4741 sit4::KanMX4 pho8::HPH | [49] |
| vps27Δpho8Δ c | BY4741 vps27::MXHIS3 pho8::HPH | This study |
| sit4Δvps27Δpho8Δ c | BY4741 sit4::KanMX4 vps27::MXHIS3 pho8::HPH | This study |
| BY4742 d | Matα his3Δ1, leu2Δ0, lys2Δ0, ura3Δ0 | EUROSCARF |
| sit4Δ d | BY4742 sit4::KanMX4 | EUROSCARF |
| vps27∆ d | BY4742 vps27::MXHIS3 | This study |
| sit4∆vps27∆ d | BY4742 sit4::KanMX4 vps27::MXHIS3 | This study |
| BY4741 Rho0 | BY4741 Rho0 | [50] |
| Primer | Sequence |
|---|---|
| COX1 fw | CTACAGATACAGCATTTCCAAGA |
| COX1 rv | GTGCCTGAATAGATGATAATGGT |
| COX3 fw | TTGAAGCTGTACAACCTACC |
| COX3 rv | CCTGCGATTAAGGCATGATG |
| ACT1 fw | GTATGTGTAAAGCCGGTTTTG |
| ACT1 rv | CATGATACCTTGGTGTCTTGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins, T.S.; Correia, M.; Pinheiro, D.; Lemos, C.; Mendes, M.V.; Pereira, C.; Costa, V. Sit4 Genetically Interacts with Vps27 to Regulate Mitochondrial Function and Lifespan in Saccharomyces cerevisiae. Cells 2024, 13, 655. https://doi.org/10.3390/cells13080655
Martins TS, Correia M, Pinheiro D, Lemos C, Mendes MV, Pereira C, Costa V. Sit4 Genetically Interacts with Vps27 to Regulate Mitochondrial Function and Lifespan in Saccharomyces cerevisiae. Cells. 2024; 13(8):655. https://doi.org/10.3390/cells13080655
Chicago/Turabian StyleMartins, Telma S., Miguel Correia, Denise Pinheiro, Carolina Lemos, Marta Vaz Mendes, Clara Pereira, and Vítor Costa. 2024. "Sit4 Genetically Interacts with Vps27 to Regulate Mitochondrial Function and Lifespan in Saccharomyces cerevisiae" Cells 13, no. 8: 655. https://doi.org/10.3390/cells13080655
APA StyleMartins, T. S., Correia, M., Pinheiro, D., Lemos, C., Mendes, M. V., Pereira, C., & Costa, V. (2024). Sit4 Genetically Interacts with Vps27 to Regulate Mitochondrial Function and Lifespan in Saccharomyces cerevisiae. Cells, 13(8), 655. https://doi.org/10.3390/cells13080655