

Finerenone Alleviates Over-Activation of Complement C5a-C5aR1 Axis of Macrophages by Regulating G Protein Subunit Alpha i2 to Improve Diabetic Nephropathy

Abstract
1. Introduction
2. Materials and Methods
2.1. Human Renal Biopsy Samples
2.2. Animal Model
2.3. HFD/STZ-Induced DN Model in Global C5aR1 Knockout Mice
2.4. Cell Culture
2.5. Biochemical Analysis of Serum/Urine/Cell Supernatant Samples
2.6. Renal Histology
2.7. Transmission Electron Microscopy
2.8. Tissue RNA Isolation and RT-qPCR
2.9. Western Blot
2.10. Immunofluorescence and Immunohistochemistry
2.11. Flow Cytometry
2.12. Bioinformatic Analyses
2.13. Statistics Analyses
3. Results
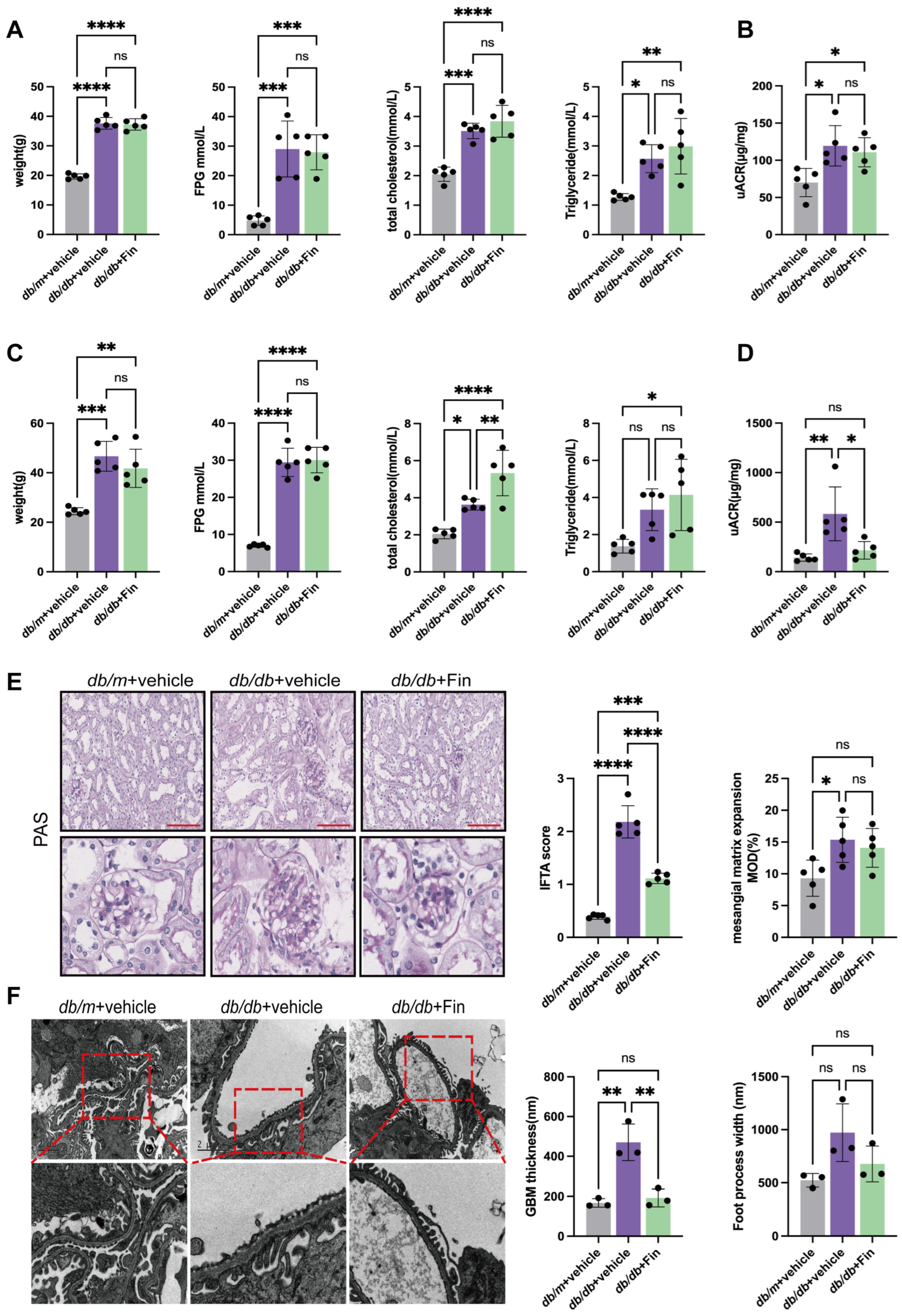
3.1. Finerenone Alleviated Renal Injury in db/db Mice
3.2. Finerenone Affected the C5aR1/Gnαi2 Axis by Inhibiting the Transcriptional Regulation of Gnαi2 by the MR
3.3. Finerenone Alleviated C5a-C5aR1-Associated Chemotaxis in db/db Mice
3.4. C5aR1 Deficiency Alleviated Renal Injury in HFD/STZ-Induced Diabetic Mice
3.5. Finerenone Alleviated HG-Induced Injuries and M1 Polarization of Macrophage via Inhibiting Gnαi2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, L.; Peng, W.; Zhao, Z.; Zhang, M.; Shi, Z.; Song, Z.; Zhang, X.; Li, C.; Huang, Z.; Sun, X.; et al. Prevalence and Treatment of Diabetes in China, 2013–2018. JAMA 2021, 326, 2498–2506. [Google Scholar] [CrossRef] [PubMed]
- Gaede, P.; Vedel, P.; Larsen, N.; Jensen, G.V.; Parving, H.H.; Pedersen, O. Multifactorial intervention and cardiovascular disease in patients with type 2 diabetes. N. Engl. J. Med. 2003, 348, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Gæde, P.; Oellgaard, J.; Carstensen, B.; Rossing, P.; Lund-Andersen, H.; Parving, H.H.; Pedersen, O. Years of life gained by multifactorial intervention in patients with type 2 diabetes mellitus and microalbuminuria: 21 years follow-up on the Steno-2 randomised trial. Diabetologia 2016, 59, 2298–2307. [Google Scholar] [CrossRef] [PubMed]
- Gaddam, K.K.; Pimenta, E.; Husain, S.; Calhoun, D.A. Aldosterone and cardiovascular disease. Curr. Probl. Cardiol. 2009, 34, 51–84. [Google Scholar] [CrossRef]
- Fuller, P.J.; Young, M.J. Mechanisms of mineralocorticoid action. Hypertension 2005, 46, 1227–1235. [Google Scholar] [CrossRef]
- Thomas, W.; Harvey, B.J. Mechanisms underlying rapid aldosterone effects in the kidney. Annu. Rev. Physiol. 2011, 73, 335–357. [Google Scholar] [CrossRef]
- Gorini, S.; Marzolla, V.; Mammi, C.; Armani, A.; Caprio, M. Mineralocorticoid Receptor and Aldosterone-Related Biomarkers of End-Organ Damage in Cardiometabolic Disease. Biomolecules 2018, 8, 96. [Google Scholar] [CrossRef]
- Belden, Z.; Deiuliis, J.A.; Dobre, M.; Rajagopalan, S. The Role of the Mineralocorticoid Receptor in Inflammation: Focus on Kidney and Vasculature. Am. J. Nephrol. 2017, 46, 298–314. [Google Scholar] [CrossRef]
- Barrera-Chimal, J.; Lima-Posada, I.; Bakris, G.L.; Jaisser, F. Mineralocorticoid receptor antagonists in diabetic kidney disease—Mechanistic and therapeutic effects. Nat. Rev. Nephrol. 2022, 18, 56–70. [Google Scholar] [CrossRef]
- Bakris, G.L.; Agarwal, R.; Anker, S.D.; Pitt, B.; Ruilope, L.M.; Rossing, P.; Kolkhof, P.; Nowack, C.; Schloemer, P.; Joseph, A.; et al. Effect of Finerenone on Chronic Kidney Disease Outcomes in Type 2 Diabetes. N. Engl. J. Med. 2020, 383, 2219–2229. [Google Scholar] [CrossRef]
- Pitt, B.; Filippatos, G.; Agarwal, R.; Anker, S.D.; Bakris, G.L.; Rossing, P.; Joseph, A.; Kolkhof, P.; Nowack, C.; Schloemer, P.; et al. Cardiovascular Events with Finerenone in Kidney Disease and Type 2 Diabetes. N. Engl. J. Med. 2021, 385, 2252–2263. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.M.; Ziemann, M.; Thallas-Bonke, V.; Snelson, M.; Kumar, V.; Laskowski, A.; Nguyen, T.V.; Huynh, K.; Clarke, M.V.; Libianto, R.; et al. Complement C5a Induces Renal Injury in Diabetic Kidney Disease by Disrupting Mitochondrial Metabolic Agility. Diabetes 2020, 69, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.M.; Snelson, M.; Østergaard, J.A.; Coughlan, M.T. The Complement Pathway: New Insights into Immunometabolic Signaling in Diabetic Kidney Disease. Antioxid. Redox Signal. 2022, 37, 781–801. [Google Scholar] [CrossRef] [PubMed]
- Gerard, N.P.; Gerard, C. The chemotactic receptor for human C5a anaphylatoxin. Nature 1991, 349, 614–617. [Google Scholar] [CrossRef]
- Ohno, M.; Hirata, T.; Enomoto, M.; Araki, T.; Ishimaru, H.; Takahashi, T.A. A putative chemoattractant receptor, C5L2, is expressed in granulocyte and immature dendritic cells, but not in mature dendritic cells. Mol. Immunol. 2000, 37, 407–412. [Google Scholar] [CrossRef]
- Klos, A.; Tenner, A.J.; Johswich, K.O.; Ager, R.R.; Reis, E.S.; Köhl, J. The role of the anaphylatoxins in health and disease. Mol. Immunol. 2009, 46, 2753–2766. [Google Scholar] [CrossRef]
- Li, L.; Wei, T.; Liu, S.; Wang, C.; Zhao, M.; Feng, Y.; Ma, L.; Lu, Y.; Fu, P.; Liu, J. Complement C5 activation promotes type 2 diabetic kidney disease via activating STAT3 pathway and disrupting the gut-kidney axis. J. Cell. Mol. Med. 2021, 25, 960–974. [Google Scholar] [CrossRef]
- Lake, B.B.; Menon, R.; Winfree, S.; Hu, Q.; Melo Ferreira, R.; Kalhor, K.; Barwinska, D.; Otto, E.A.; Ferkowicz, M.; Diep, D.; et al. An atlas of healthy and injured cell states and niches in the human kidney. Nature 2023, 619, 585–594. [Google Scholar] [CrossRef]
- Zarantonello, A.; Revel, M.; Grunenwald, A.; Roumenina, L.T. C3-dependent effector functions of complement. Immunol. Rev. 2023, 313, 120–138. [Google Scholar] [CrossRef]
- Gasque, P.; Chan, P.; Fontaine, M.; Ischenko, A.; Lamacz, M.; Götze, O.; Morgan, B.P. Identification and characterization of the complement C5a anaphylatoxin receptor on human astrocytes. J. Immunol. 1995, 155, 4882–4889. [Google Scholar] [CrossRef]
- Pandey, S.; Maharana, J.; Li, X.X.; Woodruff, T.M.; Shukla, A.K. Emerging Insights into the Structure and Function of Complement C5a Receptors. Trends Biochem. Sci. 2020, 45, 693–705. [Google Scholar] [CrossRef] [PubMed]
- Weis, W.I.; Kobilka, B.K. The Molecular Basis of G Protein-Coupled Receptor Activation. Annu. Rev. Biochem. 2018, 87, 897–919. [Google Scholar] [CrossRef] [PubMed]
- Katritch, V.; Cherezov, V.; Stevens, R.C. Structure-function of the G protein-coupled receptor superfamily. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 531–556. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.C.; Condon, N.D.; Hill, T.A.; Reid, R.C.; Fairlie, D.P.; Lim, J. Ras-Related Protein Rab5a Regulates Complement C5a Receptor Trafficking, Chemotaxis, and Chemokine Secretion in Human Macrophages. J. Innate Immun. 2023, 15, 468–484. [Google Scholar] [CrossRef]
- Li, X.; Chen, X.; Gong, S.; Zhao, J.; Yao, C.; Zhu, H.; Xiao, R.; Qin, Y.; Li, R.; Sun, N.; et al. Platelets promote CRC by activating the C5a/C5aR1 axis via PSGL-1/JNK/STAT1 signaling in tumor-associated macrophages. Theranostics 2023, 13, 2040–2056. [Google Scholar] [CrossRef]
- Desai, J.V.; Kumar, D.; Freiwald, T.; Chauss, D.; Johnson, M.D.; Abers, M.S.; Steinbrink, J.M.; Perfect, J.R.; Alexander, B.; Matzaraki, V.; et al. C5a-licensed phagocytes drive sterilizing immunity during systemic fungal infection. Cell 2023, 186, 2802–2822.e22. [Google Scholar] [CrossRef]
- van den Bos, E.; Ambrosy, B.; Horsthemke, M.; Walbaum, S.; Bachg, A.C.; Wettschureck, N.; Innamorati, G.; Wilkie, T.M.; Hanley, P.J. Knockout mouse models reveal the contributions of G protein subunits to complement C5a receptor-mediated chemotaxis. J. Biol. Chem. 2020, 295, 7726–7742. [Google Scholar] [CrossRef]
- Yao, L.; Liang, X.; Liu, Y.; Li, B.; Hong, M.; Wang, X.; Chen, B.; Liu, Z.; Wang, P. Non-steroidal mineralocorticoid receptor antagonist finerenone ameliorates mitochondrial dysfunction via PI3K/Akt/eNOS signaling pathway in diabetic tubulopathy. Redox Biol. 2023, 68, 102946. [Google Scholar] [CrossRef]
- Wu, P.; Wang, Y.; Davis, M.E.; Zuckerman, J.E.; Chaudhari, S.; Begg, M.; Ma, R. Store-Operated Ca2+ Channels in Mesangial Cells Inhibit Matrix Protein Expression. J. Am. Soc. Nephrol. 2015, 26, 2691–2702. [Google Scholar] [CrossRef]
- Tervaert, T.W.; Mooyaart, A.L.; Amann, K.; Cohen, A.H.; Cook, H.T.; Drachenberg, C.B.; Ferrario, F.; Fogo, A.B.; Haas, M.; de Heer, E.; et al. Pathologic classification of diabetic nephropathy. J. Am. Soc. Nephrol. 2010, 21, 556–563. [Google Scholar] [CrossRef]
- Kelly, D.J.; Wilkinson-Berka, J.L.; Allen, T.J.; Cooper, M.E.; Skinner, S.L. A new model of diabetic nephropathy with progressive renal impairment in the transgenic (mRen-2)27 rat (TGR). Kidney Int. 1998, 54, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Jensen, E.B.; Gundersen, H.J.; Osterby, R. Determination of membrane thickness distribution from orthogonal intercepts. J. Microsc. 1979, 115, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhang, Q.Y.; Xu, L.; Li, M.; Sun, Q.Y. Icariin ameliorates LPS-induced acute lung injury in mice via complement C5a-C5aR1 and TLR4 signaling pathways. Int. Immunopharmacol. 2024, 131, 111802. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.; Brodermann, M.H.; McCaffary, D.; Iqbal, A.J.; Greaves, D.R. Netrin-1 Reduces Monocyte and Macrophage Chemotaxis towards the Complement Component C5a. PLoS ONE 2016, 11, e0160685. [Google Scholar] [CrossRef]
- Calle, P.; Hotter, G. Macrophage Phenotype and Fibrosis in Diabetic Nephropathy. Int. J. Mol. Sci. 2020, 21, 2806. [Google Scholar] [CrossRef]
- Sun, Z.J.; Li, X.Q.; Chang, D.Y.; Wang, S.X.; Liu, G.; Chen, M.; Zhao, M.H. Complement deposition on renal histopathology of patients with diabetic nephropathy. Diabetes Metab. 2019, 45, 363–368. [Google Scholar] [CrossRef]
- Li, M.R.; Sun, Z.J.; Chang, D.Y.; Yu, X.J.; Wang, S.X.; Chen, M.; Zhao, M.H. C3c deposition predicts worse renal outcomes in patients with biopsy-proven diabetic kidney disease in type 2 diabetes mellitus. J. Diabetes 2022, 14, 291–297. [Google Scholar] [CrossRef]
- Li, X.Q.; Chang, D.Y.; Chen, M.; Zhao, M.H. Complement activation in patients with diabetic nephropathy. Diabetes Metab. 2019, 45, 248–253. [Google Scholar] [CrossRef]
- Huang, L.; Wu, W.; Wang, X. Analysis of the microecological mechanism of diabetic kidney disease based on the theory of “gut-kidney axis”: A systematic review. Open Life Sci. 2024, 19, 20220909. [Google Scholar] [CrossRef]
- Kumar Pasupulati, A.; Chitra, P.S.; Reddy, G.B. Advanced glycation end products mediated cellular and molecular events in the pathology of diabetic nephropathy. Biomol. Concepts 2016, 7, 293–309. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence 5′ to 3′ | |
|---|---|---|
| Forward | Reverse | |
| Mouse | ||
| C5ar1 | ATGGACCCCATAGATAACAGCA | GAGTAGATGATAAGGGCTGCAAC |
| Gnαi2 | CAGAGGAACAAGGGATGCTTC | TAAGCGGCTGAGTCATTGAGC |
| Cxcl15 | CAAGGCTGGTCCATGCTCC | TGCTATCACTTCCTTTCTGTTGC |
| Ccl2 | TTAAAAACCTGGATCGGAACCAA | GCATTAGCTTCAGATTTACGGGT |
| 18 s | GTAACCCGTTGAACCCCATTC | GCCTCACTAAACCATCCAATCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.-H.; Sun, Z.-J.; Tang, S.C.W.; Zhao, M.-H.; Chen, M.; Chang, D.-Y. Finerenone Alleviates Over-Activation of Complement C5a-C5aR1 Axis of Macrophages by Regulating G Protein Subunit Alpha i2 to Improve Diabetic Nephropathy. Cells 2025, 14, 337. https://doi.org/10.3390/cells14050337
Li Z-H, Sun Z-J, Tang SCW, Zhao M-H, Chen M, Chang D-Y. Finerenone Alleviates Over-Activation of Complement C5a-C5aR1 Axis of Macrophages by Regulating G Protein Subunit Alpha i2 to Improve Diabetic Nephropathy. Cells. 2025; 14(5):337. https://doi.org/10.3390/cells14050337
Chicago/Turabian StyleLi, Zi-Han, Zi-Jun Sun, Sydney C. W. Tang, Ming-Hui Zhao, Min Chen, and Dong-Yuan Chang. 2025. "Finerenone Alleviates Over-Activation of Complement C5a-C5aR1 Axis of Macrophages by Regulating G Protein Subunit Alpha i2 to Improve Diabetic Nephropathy" Cells 14, no. 5: 337. https://doi.org/10.3390/cells14050337
APA StyleLi, Z.-H., Sun, Z.-J., Tang, S. C. W., Zhao, M.-H., Chen, M., & Chang, D.-Y. (2025). Finerenone Alleviates Over-Activation of Complement C5a-C5aR1 Axis of Macrophages by Regulating G Protein Subunit Alpha i2 to Improve Diabetic Nephropathy. Cells, 14(5), 337. https://doi.org/10.3390/cells14050337