

Therapeutic Targets in Glioblastoma: Molecular Pathways, Emerging Strategies, and Future Directions


Abstract
1. Introduction
2. Molecular Pathogenesis of GBM
3. Emerging Therapeutic Targets Under Investigation
3.1. Targeting Growth Factor Receptors
3.2. Signal Transduction Pathway Inhibitors
3.3. Epigenetic Modulators
3.4. Immunotherapy
3.5. Immune Checkpoint Inhibitors (ICIs)
3.5.1. PD-1/PD-L1 Blockade
3.5.2. CTLA-4 Inhibition
3.5.3. Next-Generation Checkpoints (LAG-3, TIM-3, and Others)
3.6. Chimeric Antigen Receptor (CAR) T Cell Therapy
Established Targets
3.7. Vaccines and Peptide-Based Immunotherapies Under Investigation
3.7.1. Peptide-Based Vaccines
3.7.2. Dendritic Cell (DC) Vaccines
3.7.3. Cell-Penetrating and Tumor-Targeting Peptides
3.8. Oncolytic Virus Therapies
3.8.1. Virus Platforms
3.8.2. Mechanistic Synergies
3.9. Cytokine-Based Therapies
3.10. Targeting the Tumor Microenvironment: Tumor-Associated Macrophages (TAMs)
3.11. Targeting Tumor Metabolism
3.12. Bypassing the Blood–Brain Barrier
3.13. Drug Repurposing and Combination Therapies
3.14. Oncolytic Viruses and Gene-Based Approaches
3.15. Nanotechnology and Drug Delivery Systems
3.16. Molecular Profiling and Biomarkers
3.17. Combination Therapies
4. Challenges and Future Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berger, T.R.; Wen, P.Y.; Lang-Orsini, M.; Chukwueke, U.N. World Health Organization 2021 Classification of Central Nervous System Tumors and Implications for Therapy for Adult-Type Gliomas: A Review. JAMA Oncol. 2022, 8, 1493–1501. [Google Scholar] [PubMed]
- Wen, P.Y.; Kesari, S. Malignant gliomas in adults. N. Engl. J. Med. 2008, 359, 492–507. [Google Scholar] [PubMed]
- Sottoriva, A.; Spiteri, I.; Piccirillo, S.G.M.; Touloumis, A.; Collins, V.P.; Marioni, J.C.; Curtis, C.; Watts, C.; Tavaré, S. Intratumor heterogeneity in human glioblastoma reflects cancer evolutionary dynamics. Proc. Natl. Acad. Sci. USA 2013, 110, 4009–4014. [Google Scholar] [PubMed]
- Wesseling, P.; Capper, D. WHO 2016 Classification of gliomas. Neuropathol. Appl. Neurobiol. 2018, 44, 139–150. [Google Scholar]
- Ostrom, Q.T.; Gittleman, H.; Truitt, G.; Boscia, A.; Kruchko, C.; Barnholtz-Sloan, J. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2011–2015. Neuro Oncol. 2018, 20 (Suppl. S4), iv1–iv86. [Google Scholar]
- Thakkar, J.P.; Dolecek, T.A.; Horbinski, C.; Ostrom, Q.T.; Lightner, D.D.; Barnholtz-Sloan, J.S.; Villano, J.L. Epidemiologic and molecular prognostic review of glioblastoma. Cancer Epidemiol. Biomark. Prev. 2014, 23, 1985–1996. [Google Scholar]
- Ostrom, Q.T.; Bauchet, L.; Davis, F.G.; Deltour, I.; Fisher, J.L.; Eastman Langer, C.; Pekmezci, M.; Schwartzbaum, J.A.; Turner, M.C.; Walsh, K.M.; et al. The epidemiology of glioma in adults: A “state of the science” review. Neuro Oncol. 2014, 16, 896–913. [Google Scholar]
- Bondy, M.L.; Scheurer, M.E.; Malmer, B.; Barnholtz-Sloan, J.S.; Davis, F.G.; Il’yasova, D.; Kruchko, C.; McCarthy, B.J.; Rajaraman, P.; Schwartzbaum, J.A.; et al. Brain tumor epidemiology: Consensus from the Brain Tumor Epidemiology Consortium. Cancer 2008, 113, 1953–1968. [Google Scholar]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar]
- Shaw, R.; Basu, M.; Karmakar, S.; Ghosh, M.K. MGMT in TMZ-based glioma therapy: Multifaceted insights and clinical trial perspectives. Biochim. Biophys. Acta Mol. Cell Res. 2024, 1871, 119673. [Google Scholar] [CrossRef]
- Weller, M.; Stupp, R.; Hegi, M.E.; van den Bent, M.; Tonn, J.C.; Sanson, M.; Wick, W.; Reifenberger, G. Personalized care in neuro-oncology coming of age: Why we need MGMT and 1p/19q testing in glioma patients in clinical practice. Neuro Oncol. 2012, 14 (Suppl. S4), iv100–iv108. [Google Scholar] [CrossRef] [PubMed]
- Sanai, N.; Berger, M.S. Surgical oncology for gliomas: The state of the art. Nat. Rev. Clin. Oncol. 2018, 15, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, T.; Kawauchi, D.; Ohno, M.; Miyakita, Y.; Takahashi, M.; Yanagisawa, S.; Osawa, S.; Fujita, S.; Omura, T.; Narita, Y. Risk factors of distant recurrence and dissemination of IDH wild-type glioblastoma: A single-center study and meta-analysis. Cancers 2024, 16, 2873. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M. The blood-brain barrier: Bottleneck in brain drug development. NeuroRx 2005, 2, 3–14. [Google Scholar] [CrossRef]
- Almeida, J.P.; Chaichana, K.L.; Rincon-Torroella, J.; Quinones-Hinojosa, A. The value of extent of resection of glioblastomas: Clinical evidence and current approach. Curr. Neurol. Neurosci. Rep. 2015, 15, 517. [Google Scholar]
- Keime-Guibert, F.; Chinot, O.; Taillandier, L.; Cartalat-Carel, S.; Frenay, M.; Kantor, G.; Guillamo, J.-S.; Jadaud, E.; Colin, P.; Bondiau, P.-Y.; et al. Radiotherapy for glioblastoma in the elderly. N. Engl. J. Med. 2007, 356, 1527–1535. [Google Scholar] [CrossRef]
- Hegi, M.E.; Diserens, A.-C.; Gorlia, T.; Hamou, M.-F.; de Tribolet, N.; Weller, M.; Kros, J.M.; Hainfellner, J.A.; Mason, W.; Mariani, L.; et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. N. Engl. J. Med. 2005, 352, 997–1003. [Google Scholar]
- Brandes, A.A.; Franceschi, E.; Tosoni, A.; Benevento, F.; Scopece, L.; Mazzocchi, V.; Bacci, A.; Agati, R.; Calbucci, F.; Ermani, M. Temozolomide concomitant and adjuvant to radiotherapy in elderly patients with glioblastoma: Correlation with MGMT promoter methylation status. Cancer 2009, 115, 3512–3518. [Google Scholar] [CrossRef]
- Friedman, H.S.; Kerby, T.; Calvert, H. Temozolomide and treatment of malignant glioma. Clin. Cancer Res. 2000, 6, 2585–2597. [Google Scholar]
- Stummer, W.; Pichlmeier, U.; Meinel, T.; Wiestler, O.D.; Zanella, F.; Reulen, H.-J. ALA-Glioma Study Group Fluorescence-guided surgery with 5-aminolevulinic acid for resection of malignant glioma: A randomized controlled multicentre phase III trial. Lancet Oncol. 2006, 7, 392–401. [Google Scholar] [CrossRef]
- Chanbour, H.; Chotai, S. Review of Intraoperative Adjuncts for Maximal Safe Resection of Gliomas and Its Impact on Outcomes. Cancers 2022, 14, 5705. [Google Scholar] [CrossRef] [PubMed]
- Kubben, P.L.; ter Meulen, K.J.; Schijns, O.E.M.G.; ter Laak-Poort, M.P.; van Overbeeke, J.J.; van Santbrink, H. Intraoperative MRI-guided resection in glioblastoma: Impact on extent of resection and survival. Acta Neurochir. 2010, 152, 1337–1345. [Google Scholar]
- Armstrong, T.S.; Lacour, N.; Gilbert, M.R. Mental health in glioblastoma patients: Is it time to look beyond physical function? Neuro Oncol. 2013, 15, 875–876. [Google Scholar]
- Tan, A.C.; Ashley, D.M.; López, G.Y.; Malinzak, M.; Friedman, H.S.; Khasraw, M. Management of glioblastoma: State of the art and future directions. CA Cancer J. Clin. 2020, 70, 299–312. [Google Scholar]
- Lacroix, M.; Abi-Said, D.; Fourney, D.R.; Gokaslan, Z.L.; Shi, W.; DeMonte, F.; Lang, F.F.; McCutcheon, I.E.; Hassenbusch, S.J.; Holland, E.; et al. A multivariate analysis of 416 patients with glioblastoma multiforme: Prognosis, extent of resection, and survival. J. Neurosurg. 2001, 95, 190–198. [Google Scholar]
- Verhaak, R.G.W.; Hoadley, K.A.; Purdom, E.; Wang, V.; Qi, Y.; Wilkerson, M.D.; Miller, C.R.; Ding, L.; Golub, T.; Mesirov, J.P.; et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010, 17, 98–110. [Google Scholar]
- Furnari, F.B.; Fenton, T.; Bachoo, R.M.; Mukasa, A.; Stommel, J.M.; Stegh, A.; Hahn, W.C.; Ligon, K.L.; Louis, D.N.; Brennan, C.; et al. Malignant astrocytic glioma: Genetics, biology, and paths to treatment. Genes Dev. 2007, 21, 2683–2710. [Google Scholar]
- Brennan, C.W.; Verhaak, R.G.; McKenna, A.; Campos, B.; Noushmehr, H.; Salama, S.R.; Zheng, S.; Chakravarty, D.; Sanborn, J.Z.; Berman, S.H.; et al. The somatic genomic landscape of glioblastoma. Cell 2013, 155, 462–477. [Google Scholar]
- Li, R.; Li, H.; Yan, W.; Yang, P.; Bao, Z.; Zhang, C.; Jiang, T.; You, Y. Genetic and clinical characteristics of primary and secondary glioblastoma is associated with differential molecular subtype distribution. Oncotarget 2015, 6, 7318–7324. [Google Scholar]
- Cancer Genome Atlas Research Network. Comprehensive genomic characterization defines human glioblastoma genes and core pathways. Nature 2008, 455, 1061–1068. [Google Scholar]
- Gan, H.K.; Cvrljevic, A.N.; Johns, T.G. The epidermal growth factor receptor variant III (EGFRvIII): Where wild things are altered. Am. J. Cancer Res. 2013, 3, 221–237. [Google Scholar]
- Wanis, H.A.; Møller, H.; Ashkan, K.; Davies, E.A. The incidence of major subtypes of primary brain tumors in adults in England, 1995–2017. Neuro Oncol. 2021, 23, 1371–1382. [Google Scholar] [CrossRef] [PubMed]
- Grochans, S.; Cybulska, A.M.; Simińska, D.; Korbecki, J.; Kojder, K.; Chlubek, D.; Baranowska-Bosiacka, I. Epidemiology of Glioblastoma Multiforme–Literature Review. Cancers 2022, 14, 2412. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization classification of tumors of the central nervous system. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar]
- Ohgaki, H.; Kleihues, P. Genetic pathways to primary and secondary glioblastoma. Am. J. Pathol. 2007, 170, 1445–1453. [Google Scholar] [CrossRef]
- Yan, H.; Parsons, D.W.; Jin, G.; McLendon, R.; Rasheed, B.A.; Yuan, W.; Kos, I.; Batinic-Haberle, I.; Jones, S.; Riggins, G.J.; et al. IDH1 and IDH2 mutations in gliomas. N. Engl. J. Med. 2009, 360, 765–773. [Google Scholar]
- Wang, C.J.; Choe, K.S. Genomic Landscape of Glioblastoma and the Potential Clinical Utility. CNS Oncol. 2014, 3, 169–172. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas Research Network. Comprehensive, integrative genomic analysis of diffuse lower-grade gliomas. N. Engl. J. Med. 2015, 372, 2481–2498. [Google Scholar]
- Brown, T.J.; Brennan, M.C.; Li, M.; Church, E.W.; Brandmeir, N.J.; Rakszawski, K.L.; Patel, A.S.; Rizk, E.B.; Suki, D.; Sawaya, R.; et al. Association of the extent of resection with survival in glioblastoma: A systematic review and meta-analysis. JAMA Oncol. 2016, 2, 1460–1469. [Google Scholar] [CrossRef]
- Komotar, R.J.; Otten, M.L.; Moise, G.; Connolly, E.S., Jr. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma-a critical review. Clin. Med. Oncol. 2008, 2, 421–422. [Google Scholar]
- Kamson, D.O.; Grossman, S.A. The Role of Temozolomide in Patients with Newly Diagnosed Wild-Type IDH, Unmethylated MGMTp Glioblastoma During the COVID-19 Pandemic. JAMA Oncol. 2021, 7, 675–676. [Google Scholar] [PubMed]
- Maragkos, G.A.; Schüpper, A.J.; Lakomkin, N.; Sideras, P.; Price, G.; Baron, R.; Hamilton, T.; Haider, S.; Lee, I.Y.; Hadjipanayis, C.G.; et al. Fluorescence-guided high-grade glioma surgery more than four hours after 5-aminolevulinic acid administration. Front Neurol. 2021, 12, 644804. [Google Scholar] [PubMed]
- Moiraghi, A.; Roux, A.; Peeters, S.; Pelletier, J.B.; Baroud, M.; Trancart, B.; Oppenheim, C.; Lechapt, E.; Benevello, C.; Parraga, E.; et al. Feasibility, Safety and Impact on Overall Survival of Awake Resection for Newly Diagnosed Supratentorial IDH-Wildtype Glioblastomas in Adults. Cancers 2021, 13, 2911. [Google Scholar] [CrossRef]
- Zhao, Y.-H.; Wang, Z.-F.; Pan, Z.-Y.; Péus, D.; Delgado-Fernandez, J.; Pallud, J.; Li, Z.-Q. A meta-analysis of survival outcomes following reoperation in recurrent glioblastoma: Time to consider the timing of reoperation. Front. Neurol. 2019, 10, 286. [Google Scholar]
- Stupp, R.; Taillibert, S.; Kanner, A.; Read, W.; Steinberg, D.M.; Lhermitte, B.; Toms, S.; Idbaih, A.; Ahluwalia, M.S.; Fink, K.; et al. Effect of tumor-treating fields plus maintenance temozolomide vs temozolomide alone on survival in glioblastoma: A randomized clinical trial. JAMA 2017, 318, 2306–2316. [Google Scholar]
- Chinot, O.L.; Wick, W.; Mason, W.; Henriksson, R.; Saran, F.; Nishikawa, R.; Carpentier, A.F.; Hoang-Xuan, K.; Kavan, P.; Cernea, D.; et al. Bevacizumab plus radiotherapy–temozolomide for newly diagnosed glioblastoma. N. Engl. J. Med. 2014, 370, 709–722. [Google Scholar]
- Reardon, D.A.; Brandes, A.A.; Omuro, A.; Mulholland, P.; Lim, M.; Wick, A.; Baehring, J.; Ahluwalia, M.S.; Roth, P.; Bähr, O.; et al. Effect of nivolumab vs. bevacizumab in patients with recurrent glioblastoma (CheckMate 143): A randomized clinical trial. JAMA Oncol. 2020, 6, 1003–1012. [Google Scholar]
- Weller, M.; Butowski, N.A.; Tran, D.D.; Recht, L.D.; O’Rourke, D.M.; Finocchiaro, G.; Perry, J.R.; Wick, W.; Stupp, R.; Sampson, J.H.; et al. Rindopepimut with temozolomide for patients with newly diagnosed, EGFRvIII-expressing glioblastoma (ACT IV): A randomized, double-blind, international phase 3 trial. Lancet Oncol. 2017, 18, 1373–1385. [Google Scholar]
- O’Rourke, D.M.; Nasrallah, M.P.; Desai, A.; Melenhorst, J.J.; Mansfield, K.; Morrissette, J.J.D.; Martinez-Lage, M.; Brem, S.; Maloney, E.; Shen, A.; et al. A single dose of peripherally infused EGFRvIII-directed CAR T cells mediates antigen loss and induces adaptive resistance in patients with recurrent glioblastoma. Sci. Transl. Med. 2017, 9, eaao984. [Google Scholar]
- Ahmed, N.; Brawley, V.; Hegde, M.; Bielamowicz, K.; Kalra, M.; Landi, D.; Robertson, C.; Gray, T.L.; Diouf, O.; Wakefield, A.; et al. HER2-specific chimeric antigen receptor–modified virus-specific T cells for progressive glioblastoma: A phase 1 dose-escalation trial. JAMA Oncol. 2017, 3, 1094–1101. [Google Scholar]
- Quail, D.F.; Joyce, J.A. The microenvironmental landscape of brain tumors. Cancer Cell 2017, 31, 326–341. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Hawkins, C.; Clarke, I.D.; Squire, J.A.; Bayani, J.; Hide, T.; Henkelman, R.M.; Cusimano, M.D.; Dirks, P.B. Identification of human brain tumour initiating cells. Nature 2004, 432, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Perry, J.R.; Laperriere, N.; O’Callaghan, C.J.; Brandes, A.A.; Menten, J.; Phillips, C.; Fay, M.; Nishikawa, R.; Cairncross, J.G.; Roa, W.; et al. Short-course radiation plus temozolomide in elderly patients with glioblastoma. N. Engl. J. Med. 2017, 376, 1027–1037. [Google Scholar] [CrossRef] [PubMed]
- Desjardins, A.; Gromeier, M.; Herndon, J.E.; Beaubier, N.; Bolognesi, D.P.; Friedman, A.H.; Friedman, H.S.; McSherry, F.; Muscat, A.M.; Nair, S.; et al. Recurrent glioblastoma treated with recombinant poliovirus. N. Engl. J. Med. 2018, 379, 150–161. [Google Scholar] [CrossRef]
- Mathur, R.; Wang, Q.; Schupp, P.G.; Nikolic, A.; Hilz, S.; Hong, C.; Grishanina, N.R.; Kwok, D.; Stevers, N.O.; Jin, Q.; et al. Glioblastoma evolution and heterogeneity from a 3D whole-tumor perspective. Cell 2024, 187, 446–463.e16. [Google Scholar] [CrossRef]
- Patel, A.P.; Tirosh, I.; Trombetta, J.J.; Shalek, A.K.; Gillespie, S.M.; Wakimoto, H.; Cahill, D.P.; Nahed, B.V.; Curry, W.T.; Martuza, R.L.; et al. Single-cell RNA-seq highlights intratumoral heterogeneity in primary glioblastoma. Science 2014, 344, 1396–1401. [Google Scholar] [CrossRef]
- Lang, F.F.; Conrad, C.; Gomez-Manzano, C.; Yung WK, A.; Sawaya, R.; Weinberg, J.S.; Prabhu, S.S.; Rao, G.; Fuller, G.N.; Aldape, K.D.; et al. Phase I study of DNX-2401 (Delta-24-RGD) oncolytic adenovirus: Replication immunotherapeutic effects in recurrent malignant glioma. J. Clin. Oncol. 2018, 36, 1419–1429. [Google Scholar] [CrossRef]
- Todo, T.; Ito, H.; Ino, Y.; Ohtsu, H.; Ota, Y.; Shibahara, J.; Tanaka, M. Intratumoral oncolytic herpes virus G47∆ for residual or recurrent glioblastoma: A phase 2 trial. Nat. Med. 2022, 28, 1610–1619. [Google Scholar] [CrossRef]
- Brown, C.E.; Alizadeh, D.; Starr, R.; Weng, L.; Wagner, J.R.; Naranjo, A.; Ostberg, J.R.; Blanchard, M.S.; Kilpatrick, J.; Simpson, J.; et al. Regression of glioblastoma after chimeric antigen receptor T-cell therapy. N. Engl. J. Med. 2016, 375, 2561–2569. [Google Scholar] [CrossRef]
- Lerner, A.; Palmer, K.; Campion, T.; Millner, T.O.; Scott, E.; Lorimer, C.; Paraskevopoulos, D.; McKenna, G.; Marino, S.; Lewis, R.; et al. Gliomas in adults: Guidance on investigations, diagnosis, treatment and surveillance. Clin. Med. 2024, 24, 100240. [Google Scholar] [CrossRef]
- Lapointe, S.; Perry, A.; Butowski, N.A. Primary brain tumours in adults. Lancet 2018, 392, 432–446. [Google Scholar] [CrossRef] [PubMed]
- Cloughesy, T.F.; Mochizuki, A.Y.; Orpilla, J.R.; Hugo, W.; Lee, A.H.; Davidson, T.B.; Wang, A.C.; Ellingson, B.M.; Rytlewski, J.A.; Sanders, C.M.; et al. Neoadjuvant anti–PD-1 immunotherapy promotes a survival benefit with intratumoral and systemic immune responses in recurrent glioblastoma. Nat. Med. 2019, 25, 477–486. [Google Scholar] [PubMed]
- Ostrom, Q.T.; Cioffi, G.; Waite, K.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS statistical report: Primary brain and other central nervous system tu-mors diagnosed in the United States in 2014–2018. Neuro Oncol. 2021, 23 (Suppl. S3), iii1–iii105. [Google Scholar] [PubMed]
- Huse, J.T.; Holland, E.C. Targeting brain cancer: Advances in the understanding of brain tumor biology and their impact on patient care. Nat. Rev. Cancer 2010, 10, 319–326. [Google Scholar]
- Adamson, C.; Kanu, O.O.; Mehta, A.I.; Di, C.; Lin, N.; Mattox, A.K.; Bigner, D.D. Glioblastoma multiforme: A review of where we have been and where we are going. Expert Opin. Investig. Drugs 2009, 18, 1061–1083. [Google Scholar] [CrossRef]
- Lowe, S.; Bhat, K.P.; Olar, A. Current clinical management of patients with glioblastoma. Cancer Rep. 2019, 2, e1216. [Google Scholar]
- Tan, N.; Wong, M.; Nannini, M.A.; Hong, R.; Lee, L.B.; Price, S.; Williams, K.; Savy, P.P.; Yue, P.; Sampath, D.; et al. Bcl-2/Bcl-xL inhibition increases the efficacy of MEK inhibition alone and in combination with PI3 kinase inhibition in lung and pancreatic tumor models. Clin. Cancer Res. 2013, 19, 2984–2996. [Google Scholar]
- Lim, M.; Xia, Y.; Bettegowda, C.; Weller, M. Current state of immunotherapy for glioblastoma. Nat. Rev. Clin. Oncol. 2018, 15, 422–442. [Google Scholar]
- Wick, W.; Platten, M.; Meisner, C.; Felsberg, J.; Tabatabai, G.; Simon, M.; Nikkhah, G.; Papsdorf, K.; Steinbach, J.P.; Sabel, M.; et al. Temozolomide chemotherapy alone versus radiotherapy alone for malignant astrocytoma in the elderly: The NOA-08 randomized, phase 3 trial. Lancet Oncol. 2012, 13, 707–715. [Google Scholar]
- Gilbert, M.R.; Wang, M.; Aldape, K.D.; Stupp, R.; Hegi, M.E.; Jaeckle, K.A.; Armstrong, T.S.; Wefel, J.S.; Won, M.; Blumenthal, D.T.; et al. Dose-dense temozolomide for newly diagnosed glioblastoma: A randomized phase III clinical trial. J. Clin. Oncol. 2013, 31, 4085–4091. [Google Scholar]
- Friedman, H.S.; Prados, M.D.; Wen, P.Y.; Mikkelsen, T.; Schiff, D.; Abrey, L.E.; Yung, W.K.A.; Paleologos, N.; Nicholas, M.K.; Jensen, R.; et al. Bevacizumab alone and in combination with irinotecan in recurrent glioblastoma. J. Clin. Oncol. 2009, 27, 4733–4740. [Google Scholar] [PubMed]
- Westphal, M.; Hilt, D.C.; Bortey, E.; Delavault, P.; Olivares, R.; Warnke, P.C.; Whittle, I.R.; Jääskeläinen, J.; Ram, Z. A phase 3 trial of local chemotherapy with biodegradable carmustine (BCNU) wafers (Gliadel wafers) in patients with primary malignant glioma. Neuro-Oncology 2003, 5, 79–88. [Google Scholar] [PubMed]
- Herrlinger, U.; Tzaridis, T.; Mack, F.; Steinbach, J.P.; Schlegel, U.; Sabel, M.; Hau, P.; Kortmann, R.-D.; Krex, D.; Grauer, O.; et al. Lomustine-temozolomide combination therapy vs standard temozolomide in newly diagnosed glioblastoma with methylated MGMT promoter (CeTeG/NOA-09): A randomized, multicentre, open-label, phase 3 trial. Lancet 2019, 393, 678–688. [Google Scholar] [PubMed]
- Gilbert, M.R.; Dignam, J.J.; Armstrong, T.S.; Wefel, J.S.; Blumenthal, D.T.; Vogelbaum, M.A.; Colman, H.; Chakravarti, A.; Pugh, S.; Won, M.; et al. A randomized trial of bevacizumab for newly diagnosed glioblastoma. N. Engl. J. Med. 2014, 370, 699–708. [Google Scholar]
- Sanai, N.; Polley, M.-Y.; McDermott, M.W.; Parsa, A.T.; Berger, M.S. An extent of resection threshold for glioblastoma survival. J. Neurosurg. 2011, 115, 567–573. [Google Scholar]
- Giese, A.; Bjerkvig, R.; Berens, M.E.; Westphal, M. Cost of migration: Invasion of malignant gliomas and implications for treatment. J. Clin. Oncol. 2003, 21, 1624–1636. [Google Scholar]
- Weller, M.; Cloughesy, T.; Perry, J.R.; Wick, W. Standards of care for treatment of recurrent glioblastoma—Are we there yet? Neuro-Oncology 2013, 15, 4–27. [Google Scholar]
- Schell, M.; Pflüger, I.; Brugnara, G.; Isensee, F.; Neuberger, U.; Foltyn, M.; Kessler, T.; Sahm, F.; Wick, A.; Nowosielski, M.; et al. Bevacizumab plus CCNU in recurrent glioblastoma (EORTC 26101): A randomized, open-label, multicentre phase 3 trial. Lancet Oncol. 2017, 18, 1305–1316. [Google Scholar]
- Liau, L.M.; Ashkan, K.; Brem, S.; Campian, J.L.; Trusheim, J.E.; Iwamoto, F.M.; Tran, D.D.; Ansstas, G.; Cobbs, C.S.; Heth, J.A.; et al. Association of autologous tumor lysate-loaded dendritic cell vaccination with extension of survival among patients with newly diagnosed and recurrent glioblastoma: A phase 3 prospective externally controlled cohort trial. JAMA Oncol. 2023, 9, 112–121. [Google Scholar]
- Bao, S.; Wu, Q.; McLendon, R.E.; Hao, Y.; Shi, Q.; Hjelmeland, A.B.; Dewhirst, M.W.; Bigner, D.D.; Rich, J.N. Stem cell-like glioma cells promote radioresistance by preferential activation of the DNA damage response. Nature 2006, 444, 756–760. [Google Scholar]
- Berghoff, A.S.; Kiesel, B.; Widhalm, G.; Rajky, O.; Ricken, G.; Wöhrer, A.; Dieckmann, K.; Filipits, M.; Brandstetter, A.; Weller, M.; et al. Programmed death ligand 1 expression and tumor-infiltrating lymphocytes in glioblastoma. Neuro-Oncology 2015, 17, 1064–1075. [Google Scholar] [CrossRef] [PubMed]
- Van Meir, E.G.; Hadjipanayis, C.G.; Norden, A.D.; Shu, H.-K.; Wen, P.Y.; Olson, J.J. Exciting new advances in neuro-oncology: The avenue to a cure for malignant glioma. CA Cancer J. Clin. 2010, 60, 166–193. [Google Scholar]
- Sulman, E.P.; Ismaila, N.; Armstrong, T.S.; Tsien, C.; Batchelor, T.T.; Cloughesy, T.; Galanis, E.; Gilbert, M.; Gondi, V.; Lovely, M.; et al. Radiotherapy for glioblastoma: American Society of Clinical Oncology Clinical Practice Guideline. J. Clin. Oncol. 2017, 35, 2432–2438. [Google Scholar] [CrossRef] [PubMed]
- Obrador, E.; Moreno-Murciano, P.; Oriol-Caballo, M.; López-Blanch, R.; Pineda, B.; Gutiérrez-Arroyo, J.L.; Loras, A.; Gonzalez-Bonet, L.G.; Martinez-Cadenas, C.; Estrela, J.M.; et al. Glioblastoma Therapy: Past, Present and Future. Int. J. Mol. Sci. 2024, 25, 2529. [Google Scholar] [CrossRef]
- Johnson, D.R.; O’Neill, B.P. Glioblastoma survival in the United States before and during the temozolomide era. J. Neurooncol. 2012, 107, 359–364. [Google Scholar]
- Yasumoto, Y.; Miyazaki, H.; Vaidyan, L.K.; Kagawa, Y.; Ebrahimi, M.; Yamamoto, Y.; Ogata, M.; Katsuyama, Y.; Sadahiro, H.; Suzuki, M.; et al. Inhibition of fatty acid synthase decreases expression of stemness markers in glioma stem cells. PLoS ONE 2016, 11, e0147717. [Google Scholar]
- Duraj, T.; Kalamian, M.; Zuccoli, G.; Maroon, J.C.; D’Agostino, D.P.; Scheck, A.C.; Poff, A.; Winter, S.F.; Hu, J.; Klement, R.J.; et al. Clinical research framework proposal for ketogenic metabolic therapy in glioblastoma. BMC Med. 2024, 22, 578. [Google Scholar]
- Di Stefano, A.L.; Fucci, A.; Frattini, V.; Labussiere, M.; Mokhtari, K.; Zoppoli, P.; Marie, Y.; Bruno, A.; Boisselier, B.; Giry, M.; et al. Detection, characterisation, and inhibition of FGFR-TACC fusions in IDH wild-type glioma. Clin. Cancer Res. 2015, 21, 3307–3315. [Google Scholar]
- Zhuang, Q.; Yang, H.; Mao, Y. Oncogenesis of diffuse gliomas: What have we learned from clinical, pathological, and molecular features of protracted (>5 year) survivors. Brain Pathol. 2010, 20, 84–91. [Google Scholar]
- Li, H.; Jin, X.; Zhang, Z.; Xing, Y.; Kong, X. Inhibition of autophagy potentiated the apoptosis induced by the PI3K/mTOR inhibitor NVP-BEZ235 in renal cell carcinoma cells. Cell Death Dis. 2013, 4, e848. [Google Scholar]
- Taal, W.; Oosterkamp, H.M.; Walenkamp AM, E.; Dubbink, H.J.; Beerepoot, L.V.; Hanse MC, J.; Buter, J.; Honkoop, A.H.; Boerman, D.; de Vos, F.Y.F.; et al. Single-agent bevacizumab or lomustine in recurrent glioblastoma: Results of the randomized BELOB trial. JAMA Oncol. 2015, 1, 230–237. [Google Scholar]
- Weller, M.; van den Bent, M.; Hopkins, K.; Tonn, J.C.; Stupp, R.; Falini, A.; Cohen-Jonathan-Moyal, E.; Frappaz, D.; Henriksson, R.; Balana, C.; et al. EANO guideline for the diagnosis and treatment of anaplastic gliomas and glioblastoma. Lancet Oncol. 2014, 15, e395–e403. [Google Scholar] [CrossRef]
- Liang, A.; Zhou, B.; Sun, W. Integrated genomic characterization of cancer genes in glioma. Cancer Cell Int. 2017, 17, 90. [Google Scholar] [PubMed]
- Gimple, R.C.; Yang, K.; Halbert, M.E.; Agnihotri, S.; Rich, J.N. Brain cancer stem cells: Resilience through adaptive plasticity and hierarchical heterogeneity. Nat. Rev. Cancer 2022, 22, 497–514. [Google Scholar] [PubMed]
- Sampson, J.H.; Maus, M.V.; June, C.H. Immunotherapy for brain tumors. J. Clin. Oncol. 2017, 35, 2450–2456. [Google Scholar]
- Weller, M.; van den Bent, M.; Hopkins, K.; Tonn, J.C.; Stupp, R.; Falini, A.; Cohen-Jonathan-Moyal, E.; Frappaz, D.; Henriksson, R.; Balana, C.; et al. Glioblastoma: ESMO consensus recommendations on diagnosis and treatment. Ann. Oncol. 2021, 32, 1428–1447. [Google Scholar]
- Zong, H.; Verhaak, R.G.; Canoll, P. The cellular origin for malignant glioma and prospects for clinical advancements. Expert Rev. Mol. Diagn. 2012, 12, 383–394. [Google Scholar]
- Killela, P.J.; Reitman, Z.J.; Jiao, Y.; Bettegowda, C.; Agrawal, N.; Diaz LA, J.r.; Friedman, A.H.; Friedman, H.; Gallia, G.L.; Giovanella, B.C.; et al. TERT promoter mutations occur frequently in gliomas and a subset of tumors derived from cells with low rates of self-renewal. Proc. Natl. Acad. Sci. USA 2013, 110, 6021–6026. [Google Scholar]
- Dapash, M.; Castro, B.; Hou, D.; Lee-Chang, C. Current Immunotherapeutic Strategies for the Treatment of Glioblastoma. Cancers 2021, 13, 4548. [Google Scholar] [CrossRef]
- Marchini, A.; Daeffler, L.; Pozdeev, V.I.; Angelova, A.; Rommelaere, J. Immune conversion of tumor microenvironment by oncolytic viruses: The protoparvovirus H-1PV case study. Front. Immunol. 2019, 10, 1848. [Google Scholar]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug repurposing: Progress, challenges and recommendations. Nat. Rev. Drug Discov. 2019, 18, 41. [Google Scholar] [PubMed]
- Sesen, J.; Dahan, P.; Scotland, S.J.; Saland, E.; Dang, V.-T.; Lemarié, A.; Tyler, B.M.; Brem, H.; Toulas, C.; Cohen-Jonathan Moyal, E.; et al. Metformin inhibits growth of human glioblastoma cells and enhances therapeutic response. PLoS ONE 2015, 10, e0123721. [Google Scholar]
- Wang, Y.; Malik, S.; Suh, H.-W.; Xiao, Y.; Deng, Y.; Fan, R.; Huttner, A.; Bindra, R.S.; Singh, V.; Saltzman, W.M.; et al. Anti-seed PNAs targeting multiple oncomiRs for brain tumor therapy. Sci. Adv. 2023, 9, eabq7459. [Google Scholar] [PubMed]
- Marino, A.; Almici, E.; Migliorin, S.; Tapeinos, C.; Battaglini, M.; Cappello, V.; Marchetti, M.; de Vito, G.; Cicchi, R.; Pavone, F.S.; et al. Piezoelectric Barium Titanate Nanostimulators for the Treatment of Glioblastoma Multiforme. J. Colloid Interface Sci. 2019, 538, 449–461. [Google Scholar]
- Vahab, S.A.; Vyshma, K.V.; Kumar, V.S. Exosome-based drug delivery systems for enhanced neurological therapeutics. Drug Deliv. Transl. Res. 2024, 14, 2193–2211. [Google Scholar]
- Ghaznavi, H.; Afzalipour, R.; Khoei, S.; Sargazi, S.; Shirvalilou, S.; Sheervalilou, R. New insights into targeted therapy of glioblastoma using smart nanoparticles. Cancer Cell Int. 2024, 24, 160. [Google Scholar]
- Devi, C.M.; Deka, K.; Das, A.K.; Talukdar, A.; Sola, P. Recent advances in marine-derived nanoformulation for the management of glioblastoma. Mol. Biotechnol. 2024, 66, 830–843. [Google Scholar]
- Harris, M.A.; Kuang, H.; Schneiderman, Z.; Shiao, M.L.; Crane, A.T.; Chrostek, M.R.; Tăbăran, A.-F.; Pengo, T.; Liaw, K.; Xu, B.; et al. ssDNA nanotubes for selective targeting of glioblastoma and delivery of doxorubicin for enhanced survival. Sci. Adv. 2021, 7, eabl5872. [Google Scholar]
- Siddiqui, B.; Islam, M.R.; Islam, M.M.; Siddique MA, B.; Hossain, N.; Amin, M.R.; Ahmed, S.F. Targeted Delivery of Nanomedicines to Glioblastoma: Overcoming the Clinical Barrier. J. Drug Deliv. Sci. Technol. 2024, 99, 105980. [Google Scholar]
- Zhu, W.; Huang, W.; Ivanov, I.V.; Xia, H.; Moore, M.; Niederfellner, M.J.; Mellman, I.; Schreiber, R.A. CSF-1R inhibition alters macrophage polarization and blocks glioma progression. Nat. Med. 2013, 19, 873–881. [Google Scholar]
- Butowski, N.; Colman, H.; De Groot, J.F.; Omuro, A.M.; Nayak, L.; Wen, P.Y.; Cloughesy, T.F.; Marimuthu, A.; Haidar, S.; Perry, A.; et al. Orally administered colony stimulating factor 1 receptor inhibitor PLX3397 in recurrent glioblastoma: An Ivy Foundation Early Phase Clinical Trials Consortium phase II study. Neuro Oncol. 2016, 18, 557–564. [Google Scholar] [PubMed]
- Mitchell, D.A.; Batich, K.A.; Gunn, M.D.; Huang, M.-N.; Sanchez-Perez, L.; Nair, S.K.; Congdon, K.L.; Reap, E.A.; Healy, A.; Archer, D.E.; et al. Tetanus toxoid and CCL3 improve dendritic cell vaccines in mice and glioblastoma patients. Nature 2015, 519, 366–369. [Google Scholar] [PubMed]
- Laurent, S.; Forge, D.; Port, M.; Roch, A.; Robic, C.; Vander Elst, L.; Muller, R.N. Magnetic iron oxide nanoparticles: Synthesis, stabilization, vectorization, physicochemical characterizations, and biological applications. Chem. Rev. 2008, 108, 2064–2110. [Google Scholar] [CrossRef] [PubMed]
- Liau, L.M.; Ashkan, K.; Tran, D.D.; Campian, J.L.; Trusheim, J.E.; Cobbs, C.S.; Heth, J.A.; Salacz, M.; Taylor, S.; D’Andre, S.D.; et al. First results on survival from a large phase 3 clinical trial of an autologous dendritic cell vaccine in newly diagnosed glioblastoma. J. Transl. Med. 2018, 16, 142. [Google Scholar] [CrossRef]
- van Kuppevelt, T.H.; Dennissen, M.A.; van Venrooij, W.J.; Hoet, R.M.; Veerkamp, J.H. Generation and application of type-specific anti-heparan sulfate antibodies using phage display technology: Novel tools in exploring structural-functional relationships in heparan sulfate. J. Neurosci. 1998, 18, 10112–10123. [Google Scholar]
- Takeuchi, T.; Futaki, S. Current understanding of direct translocation of arginine-rich cell-penetrating peptides and its internalization mechanisms. Chem. Pharm. Bull. 2016, 64, 1431–1437. [Google Scholar]
- Banks, W.A. From blood–brain barrier to blood–brain interface: New opportunities for CNS drug delivery. Nat. Rev. Drug Discov. 2016, 15, 275–292. [Google Scholar]
- Bulbake, U.; Doppalapudi, S.; Kommineni, N.; Khan, W. Liposomal formulations in clinical use: An updated review. Pharmaceutics 2017, 9, 12. [Google Scholar] [CrossRef]
- Dadfar, S.M.; Roemhild, K.; Drude, N.I.; von Stillfried, S.; Knüchel, R.; Kiessling, F.; Lammers, T. Iron oxide nanoparticles: Diagnostic, therapeutic and theranostic applications. Adv. Drug Deliv. Rev. 2019, 138, 302–325. [Google Scholar]
- Wu, L.; Mendoza-Garcia, A.; Li, Q.; Sun, S. Organic phase syntheses of magnetic nanoparticles and their applications. Chem. Rev. 2016, 116, 10473–10512. [Google Scholar]
- Kumar, C.S.; Mohammad, F. Magnetic nanomaterials for hyperthermia-based therapy and controlled drug delivery. Adv. Drug Deliv. Rev. 2011, 63, 789–808. [Google Scholar] [CrossRef] [PubMed]
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the clinic: An update. Bioeng. Transl. Med. 2019, 4, e10143. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Fu, X.; Fu, W.; Li, Z. Biodegradable stimuli-responsive polypeptide materials for biomedical applications. Macromol. Rapid. Commun. 2015, 36, 142–157. [Google Scholar]
- Danhier, F.; Ansorena, E.; Silva, J.M.; Coco, R.; Le Breton, A. Préat VPLGA-based nanoparticles: An overview of biomedical applications. J. Control. Release 2012, 161, 505–522. [Google Scholar] [CrossRef]
- Brem, H.; Piantadosi, S.; Burger, P.C.; Walker, M.; Selker, R.; Vick, N.A.; Black, K.; Sisti, M.; Brem, S.; Mohr, G. Placebo-controlled trial of safety and efficacy of intraoperative controlled delivery by biodegradable polymers of chemotherapy for recurrent gliomas. Lancet 1995, 345, 1008–1012. [Google Scholar] [CrossRef]
- Nguyen, M.K.; Lee, D.S. Injectable biodegradable hydrogels. Macromol. Biosci. 2010, 10, 563–579. [Google Scholar] [CrossRef]
- Xu, H.; Das, M.; Reilly, M.A.; Iyer, P.; Venkatraman, S.S.; Kuang, L. Injectable biodegradable hybrid hydrogels based on a poly(amino acid) and poly(ethylene glycol) for locally targeted drug delivery. Macromolecules 2010, 43, 4305–4313. [Google Scholar]
- Sousa, F.; Lee, H.; Almeida, M.; Bazzoni, A.; Rothen-Rutishauser, B.; Petri-Fink, A. Immunostimulatory nanoparticles delivering cytokines as a novel cancer nanoadjuvant to empower glioblastoma immunotherapy. Drug Deliv. Transl. Res. 2024, 14, 2655–2667. [Google Scholar] [CrossRef]
- He, H.; Chen, S.; Zhou, J.; Li, M.; Huang, S.; Zheng, H.; Zhang, W.; Yang, F.; Wang, Z.; Wei, Y. Smart nano-delivery systems for glioblastoma treatment. Front. Bioeng. Biotechnol. 2020, 8, 724. [Google Scholar]
- Sun, Q.; Zhou, Z.; Qiu, N.; Shen, Y. Rational design of cancer nanomedicine: Nanoproperty integration and synchronization. Adv. Mater. 2017, 29, 1606628. [Google Scholar]
- Cheng, R.; Meng, F.; Deng, C.; Klok, H.-A.; Zhong, Z. Dual-responsive mesoporous silica nanoparticle-based multicore/shell nanogels for targeted and triggered intracellular drug release. Biomaterials 2013, 34, 3647–3657. [Google Scholar] [PubMed]
- Fares, J.; Wan, Y.; Mair, R.; Price, S.J. Molecular diversity in isocitrate dehydrogenase-wild-type glioblastoma. Brain Commun. 2024, 6, fcae108. [Google Scholar] [PubMed]
- Crespo, I.; Vital, A.L.; Gonzalez-Tablas, M.; Patino Mdel, C.; Otero, A.; Lopes, M.C.; de Oliveira, C.; Domingues, P.; Orfao, A.; Tabernero, M.D. Molecular and genomic alterations in glioblastoma multiforme. Am. J. Pathol. 2015, 185, 1449–1462. [Google Scholar]
- van de Geer, W.S.; Hoogstrate, Y.; Draaisma, K.; Robe, P.A.; Bins, S.; Mathijssen, R.H.J.; Kros, J.M.; French, P.J.; van den Bent, M.J.; Dirven, C.M.F. Landscape of driver gene events, biomarkers, and druggable targets identified by whole-genome sequencing of glioblastomas. Neuro-Oncol. Adv. 2022, 4, vdab177. [Google Scholar]
- Omuro, A.; DeAngelis, L.M. Glioblastoma and other malignant gliomas: A clinical review. JAMA 2013, 310, 1842–1850. [Google Scholar]
- Yousefi, Y.; Nejati, R.; Eslahi, A.; Alizadeh, F.; Farrokhi, S.; Asoodeh, A.; Mojarrad, M. Enhancing Temozolomide (TMZ) chemosensitivity using CRISPR-dCas9-mediated downregulation of O6-methylguanine DNA methyltransferase (MGMT). J. Neurooncol. 2024, 169, 129–135. [Google Scholar]
- Wong, E.T.; Yamaguchi, N.H. Treatment advances for glioblastoma. Expert Rev. Neurother. 2011, 11, 1343–1345. [Google Scholar]
- Guo, C.; Yang, Q.; Xu, P.; Ji, N.; Li, J.; Sun, T.; Wang, J.; Wang, L.; Li, C.; Wang, G.; et al. Adjuvant Temozolomide Chemotherapy with or Without Interferon Alfa Among Patients with Newly Diagnosed High-grade Gliomas: A Randomized Clinical Trial. JAMA Netw. Open 2023, 6, e2253285. [Google Scholar]
- Olivet, M.M.; Brown, M.C.; Reitman, Z.J.; Ashley, D.M.; Grant, G.A.; Yang, Y.; Markert, J.M. Clinical Applications of Immunotherapy for Recurrent Glioblastoma in Adults. Cancers 2023, 15, 3901. [Google Scholar] [CrossRef]
- Batchelor, T.T.; Duda, D.G.; di Tomaso, E.; Ancukiewicz, M.; Plotkin, S.R.; Chi, A.S.; Eichler, A.F.; Drappatz, J.; Gerard, M.; Louis, D.N.; et al. Phase II study of cediranib, an oral pan-VEGF receptor tyrosine kinase inhibitor, in patients with recurrent glioblastoma. J. Clin. Oncol. 2010, 28, 2817–2823. [Google Scholar]
- Gomez-Manzano, C.; Fueyo, J. Targeting multiple pathways in glioblastoma. J. Clin. Oncol. 2012, 30, 4058–4059. [Google Scholar]
- Piccirillo, S.G.M.; Colman, S.; Potter, N.E.; van Delft, F.W.; Lillis, S.; Carnicer, M.-J.; Kearney, L.; Watts, C.; Greaves, M. Genetic and functional diversity of propagating cells in glioblastoma. Stem Cell Rep. 2015, 4, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Carlsen, L.; El-Deiry, W.S. Anti-cancer immune responses to DNA damage response inhibitors: Molecular mechanisms and progress toward clinical translation. Front. Oncol. 2022, 12, 998388. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.R.; Zhong, X.; Sandoval, J.A.; Hickey, R.J.; Malkas, L.H. Applications of emerging molecular technologies in glioblastoma multiforme. Expert Rev. Mol. Diagn. 2014, 8, 1497–1506. [Google Scholar]
- Hasan, I.; Roy, S.; Ehexige, E.; Wu, R.; Chen, Y.; Gao, Z.; Guo, B.; Chang, C.-C. A state-of-the-art liposome technology for glioblastoma treatment. Nanoscale 2023, 15, 18108–18138. [Google Scholar]
- Liu, Y.; Lang, F.; Chou, F.-J.; Zaghloul, K.A.; Yang, C. Isocitrate Dehydrogenase Mutations in Glioma: Genetics, Biochemistry, and Clinical Indications. Biomed. 2020, 8, 294. [Google Scholar]
- Bi, W.L.; Hosny, A.; Schabath, M.B.; Giger, M.L.; Birkbak, N.J.; Mehrtash, A.; Nguyentat, P.P.; Velazquez, M.A.; Yankeelov, T.E.; van den Abbeele, A.D.; et al. Artificial intelligence in cancer imaging: Clinical challenges and applications. CA Cancer J. Clin. 2019, 69, 127–157. [Google Scholar]
- Touat, M.; Idbaih, A.; Sanson, M.; Ligon, K.L. Glioblastoma targeted therapy: Updated approaches from recent biological insights. Ann. Oncol. 2017, 28, 1457–1472. [Google Scholar]
- Taphoorn, M.J.B.; Henriksson, R.; Bottomley, A.; Cloughesy, T.; Wick, W.; Mason, W.P.; Saran, F.; Nishikawa, R.; Hilton, M.; Theodore-Oklota, C.; et al. Health-related quality of life in a randomized phase III study of bevacizumab with temozolomide and radiotherapy in newly diagnosed glioblastoma. J. Clin. Oncol. 2015, 33, 2166–2175. [Google Scholar]
- Song, I.-W.; Vo, H.H.; Chen, Y.-S.; Baysal, M.A.; Kahle, M.; Johnson, A.; Tsimberidou, A.M. Precision Oncology: Evolving Clinical Trials across Tumor Types. Cancers 2023, 15, 1967. [Google Scholar] [CrossRef]
- Aldape, K.; Brindle, K.M.; Chesler, L.; Chopra, R.; Gajjar, A.; Gilbert, M.R.; Gottardo, N.; Gutmann, D.H.; Hargrave, D.; Holland, E.C.; et al. Challenges to curing primary brain tumours. Nat. Rev. Clin. Oncol. 2019, 16, 509–520. [Google Scholar]


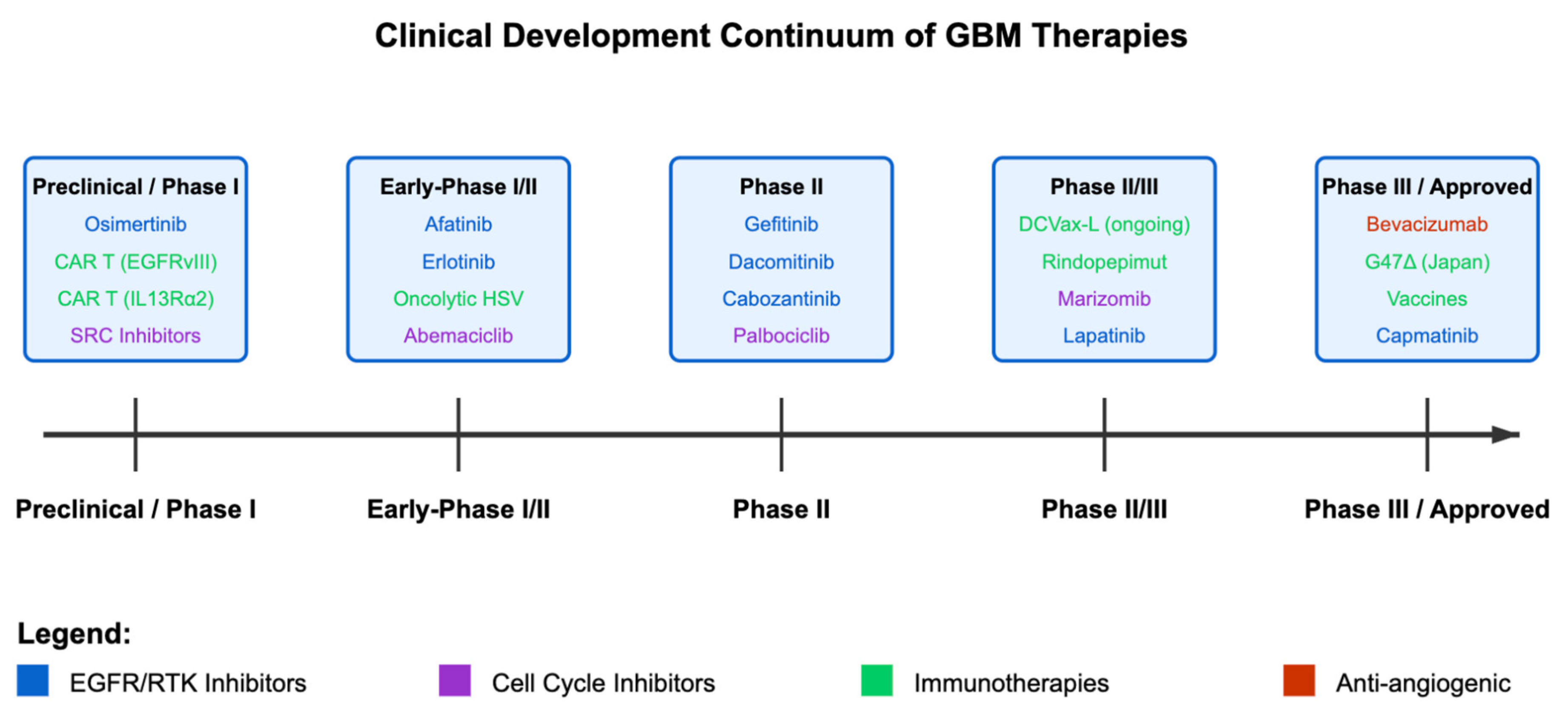

{kind=link}
{kind=link}
{kind=link}
| Genetic Mutation/Epigenetic Modification | Frequency in GBM (% of Cases) | Impact on Tumor Biology | Potential Approaches Under Investigation |
|---|---|---|---|
| TP53 Mutations | 31–38% overall; up to 65% in secondary GBMs | Disrupts cell cycle control and apoptosis | Potential for therapies targeting p53 pathways |
| PTEN Mutations | 24–37% (mainly in primary GBMs) | Activates PI3K/Akt signaling, promoting proliferation and survival | Use of PI3K/Akt pathway inhibitors |
| EGFR Amplification and Mutations | 36–60% in primary GBMs; EGFRvIII in 20–50% of amplified cases | Enhances cell growth via receptor activation | EGFR inhibitors and antibodies targeting EGFRvIII variant |
| NF1 Mutations or Deletions | 15–17% | Affects RAS/MAPK signaling pathways | Therapies targeting RAS/MAPK components |
| PIK3CA and PIK3R1 Mutations | PIK3CA: 7–10%; PIK3R1: 7–8% | Activates PI3K/Akt pathway | PI3K inhibitors |
| RB1 Mutations | 8–13% | Impairs cell cycle regulation via retinoblastoma pathway | CDK inhibitors targeting cell cycle dysregulation |
| CDKN2A Deletion (p16INK4) and p14ARF) | 31–78% in primary GBMs | Loss of cell cycle inhibition, increased proliferation | CDK4/6 inhibitors; restoring cell cycle checkpoints |
| ATRX Mutations | Common in secondary GBMs and lower-grade gliomas | Involved in telomere maintenance | Targeting telomere elongation mechanisms |
| TERT Promoter Mutations | 58% in primary; 28% in secondary GBMs | Increases telomerase activity (anti-senescence), | Telomerase inhibitors |
| MGMT Promoter Hypermethylation | 36% in primary; 75% in secondary GBMs | Reduces DNA repair capacity; better response to alkylating agents | Predictive biomarker for temozolomide efficacy |
| Hypermethylation of Tumor Suppressor Genes | RB1: 14% primary, 43% secondary; CDKN2A-p14ARF: 6% primary, 31% secondary | Silencing of genes critical for cell cycle and apoptosis | Use of demethylating agents to reactivate tumor suppressor genes |
| Loss of Heterozygosity (LOH) on Chromosome 10 | Up to 70% in primary GBMs | Associated with PTEN loss; contributes to tumor progression | Important to target PTEN pathway |
| Chromosome 9p21 Deletion | 31–78% in primary GBMs | Loss of CDKN2A locus, leading to cell cycle dysregulation | Need for therapies targeting cell cycle control |
| Signaling Pathway | Key Components | Role in GBM Progression | Potential Targeted Therapies |
|---|---|---|---|
| p53 Pathway | TP53 gene, MDM2, p21 | Regulates cell cycle and apoptosis; mutations lead to uncontrolled cell proliferation and impaired cell death | MDM2 inhibitors (e.g., RG7112), compounds restoring p53 function (e.g., PRIMA-1) |
| PI3K/AKT/mTOR Pathway | PI3K (PIK3CA), AKT, mTOR, PTEN | Promotes cellular growth, survival, and metabolism; frequently activated due to PTEN loss or PIK3CA mutations | PI3K inhibitors (e.g., BKM120), AKT inhibitors (e.g., perifosine), mTOR inhibitors (e.g., everolimus) |
| EGFR Pathway | EGFR, EGFRvIII mutant, downstream effectors (RAS, AKT) | Enhances tumor cell proliferation and survival; EGFR amplification/mutation leads to constitutive activation | EGFR tyrosine kinase inhibitors (e.g., erlotinib), monoclonal antibodies, vaccines targeting EGFRvIII |
| NF-κB Pathway | NF-κB proteins (p65, p50), IκB kinase (IKK) complex | Drives inflammation, promotes tumor growth and resistance to apoptosis | NF-κB inhibitors (e.g., parthenolide, BAY 11-7082) |
| Wnt Signaling Pathway | Wnt ligands, frizzled receptors, β-catenin | Regulates cell proliferation and differentiation; aberrant activation contributes to tumor aggressiveness | Wnt pathway inhibitors (under investigation) |
| TERT Pathway | Telomerase reverse transcriptase (TERT) | Maintains telomere length, allowing unlimited cell division | Telomerase inhibitors, TERT-targeted therapies |
| CDKN2A/pRB Pathway | CDKN2A gene (p16INK4A, p14ARF), RB1 protein | Controls cell cycle progression; loss leads to unchecked proliferation | CDK4/6 inhibitors (e.g., palbociclib), strategies to restore pathway function |
| c-Met Pathway | c-Met receptor, hepatocyte growth factor (HGF) | Promotes cell growth, invasion, and angiogenesis | c-Met inhibitors (e.g., crizotinib, cabozantinib), monoclonal antibodies (e.g., onartuzumab) |
| FGFR Pathway | FGFR receptors, FGF ligands | Involved in cell proliferation and survival; less commonly altered in GBM | FGFR inhibitors (e.g., futibatinib, pemigatinib) |
| BRAF Pathway | BRAF kinase (V600E mutation) | Activates MAPK/ERK pathway, promoting growth | BRAF inhibitors (e.g., dabrafenib, vemurafenib) |
| Src Pathway | Src family kinases | Facilitates proliferation and invasion | Src inhibitors (e.g., dasatinib) |
| RAS/MAPK Pathway | RAS proteins, RAF, MEK, ERK | Controls cell proliferation and differentiation; overactivation leads to tumor growth | MEK inhibitors, oncolytic viruses targeting RAS pathway |
| MGMT | O6-Methylguanine-DNA methyltransferase | Repairs DNA damage from alkylating agents | MGMT inhibitors |
| VEGF Signaling | Vascular endothelial growth factor (VEGF), VEGF receptors | Stimulates angiogenesis, supporting tumor vascularization | Anti-VEGF therapies (e.g., bevacizumab) |
| TGF-β Pathway | Transforming growth factor-beta (TGF-β) | Promotes invasion and immunosuppression | TGF-β inhibitors (e.g., galunisertib) |
| HDAC Pathway | Histone deacetylases | Epigenetic regulation; overactivity leads to aberrant gene expression | HDAC inhibitors (e.g., vorinostat, panobinostat) |
| Notch Pathway | Receptors (Notch1–4), ligands (Dll1, Dll3, Dll4, Jagged1–2), γ-secretase, RBPJK | Maintains GSCs, promotes treatment resistance, drives tumor growth, angiogenesis, and stemness under hypoxia | GSIs (DAPT, RO4929097), ASIs (INCB3619), miRNAs (miR-34a, miR-181c), arsenic trioxide, tipifarnib, CB-103 |
| Hedgehog Pathway | Sonic Hedgehog (SHH), patched (PTCH1/2), smoothened (SMO), GLI1/2/3 | Regulates tumor growth, stem cell maintenance, drug resistance, and promotes angiogenesis and invasion | SMO inhibitors (e.g., vismodegib, sonidegib), GLI inhibitors (e.g., GANT-61), combination therapies to overcome resistance |
| MAPK Pathway | EGFR, PDGFRA, BRAF, MAPK | Promotes cell proliferation, survival, and therapy resistance via pathway hyperactivation (high MAPK activity correlates with poor survival and increased tumor aggressiveness) | MAPK inhibitors (e.g., BRAF inhibitors); potential for combination therapies targeting MAPK and PI3K/AKT pathways |
| (a) | |||
|---|---|---|---|
| Agent | Mechanism | Clinical Phase/Population | Findings |
| Gefitinib | First-generation EGFR tyrosine kinase inhibitor (TKI) | Phase II (recurrent GBM) (e.g., NCT01520870) | - Poor BBB penetration - EGFR alterations in GBM are heterogeneous; not all tumors rely on EGFR signaling |
| Dacomitinib | Pan-EGFR TKI (inhibits EGFR, HER2, HER4) | Phase II (recurrent GBM) (e.g., NCT02447419) | - Still challenged by BBB penetration - Broader than gefitinib, but GBM evolves alternate pathways |
| Osimertinib | Third-generation EGFR TKI, better BBB permeability | Early-phase/preclinical (recurrent GBM) | - Promising in preclinical models due to improved BBB penetration - Further phase I/II trials needed to determine safety and efficacy |
| Nimotuzumab | Anti-EGFR monoclonal antibody (mAb) | Phase II/III (various GBM populations) | - Mixed results: some modest improvements in specific subgroups - Reduced toxicity vs. other anti-EGFR mAbs because of intermediate affinity |
| Depatux-M (ABT-414) | Antibody–drug conjugate targeting EGFR; delivers cytotoxic agent | Phase II/III (EGFR-amplified GBM) (e.g., NCT02573324) | - Some efficacy in EGFR-amplified GBM - Ocular toxicity reported; highlights the need for careful dosing and patient selection |
Challenges:
| |||
| (b) | |||
| Agent | Mechanism | Clinical Phase/Population | Findings |
| Cabozantinib | Inhibits MET and VEGFR2 (angiogenesis) | Phase II (recurrent GBM) (e.g., NCT00704288) | - Modest activity in heavily pretreated patients - Notable toxicities (hypertension, fatigue, etc.) |
| Capmatinib (INC280) | Selective MET inhibitor | Phase II (recurrent GBM) (e.g., NCT01870726) | - Limited efficacy overall - Possible benefit in tumors with MET amplification or alterations |
| Erdafitinib | Pan-FGFR inhibitor (incl. FGFR3-TACC3 fusions) | Phase II (recurrent GBM) (e.g., NCT01703481) | - Partial responses in some patients with FGFR alterations - Ongoing trials with biomarker selection |
Challenges:
| |||
| (c) | |||
| Agent | Mechanism | Clinical Phase/Population | Findings |
| Palbociclib | CDK4/6 inhibitor; blocks G1 → S phase transition | Phase II (recurrent GBM) (e.g., NCT01227434) | - No significant efficacy as monotherapy - Ongoing combinations with radiation or targeted agents |
| Ribociclib | CDK4/6 inhibitor | Phase I/II (recurrent GBM) (e.g., NCT02345824) | - Limited single-agent benefit - Potential synergy with other pathways (e.g., mTOR inhibitors) |
Challenges:
| |||
| (d) | |||
| Agent | Mechanism | Clinical Phase/Population | Findings |
| Bortezomib/Marizomib | Proteasome inhibitors (alter proteostasis) | Bortezomib: phase I/II; marizomib: phase III (e.g., NCT03345095) | - Bortezomib limited by BBB and toxicity - Marizomib under combination of trials (TMZ + RT), aiming for synergy |
| Bevacizumab | Anti-VEGF mAb (angiogenesis blockade) | Approved for Recurrent GBM | - Improves progression-free survival, less proven benefit in overall survival - Combined with chemo or RT |
Challenges:
| |||
| (a) | |||
|---|---|---|---|
| Agent | Target | Clinical Phase/Population | Key Findings and Rationale |
| EGFRvIII-targeted CAR T Cells | EGFRvIII mutation (common in GBM) | Early-phase (e.g., NCT02209376) | - Safe but limited efficacy due to antigen loss and immunosuppressive microenvironment |
| IL13Rα2-targeted CAR T Cells | IL13Rα2 (overexpressed in GBM) | Phase I (case reports) | - Dramatic regression in a single case report - Studies ongoing to confirm broad efficacy and overcome tumor heterogeneity |
| HER2-targeted CAR T Cells | HER2 receptor | Early-phase | - Preliminary safety established; potential synergy with other immunotherapies |
Challenges:
| |||
| (b) | |||
| Agent | Mechanism | Clinical Phase/Population | Key Findings and Rationale |
| Rindopepimut | Peptide vaccine targeting EGFRvIII | Phase III (ACT IV; NCT01480479) | - Did not improve OS vs. control - Trial halted; underscores how GBM escapes single-target therapies |
| DCVax®-L | Dendritic cell vaccine with autologous tumor lysate | Phase III (NCT00045968) | - Interim data suggest possible survival benefit - Full results pending; likely works best in low tumor burden |
Challenges:
| |||
| Agent | Virus Type/Target | Clinical Phase/Population | Key Findings and Rationale |
|---|---|---|---|
| PVSRIPO | Engineered poliovirus targeting CD155 | Phase I/II (recurrent GBM) | - Demonstrated safety; some patients have prolonged survival - Requires strong anti-tumor immune response |
| DNX-2401 | Oncolytic adenovirus selectively replicating in GBM | Phase I (recurrent GBM) (NCT00805376) | - Induces immune response; some durable remissions - Combining with other immunotherapies is under investigation |
| G47Δ | Genetically engineered herpes simplex virus | Phase II (Japan) | - Conditional approval in Japan for recurrent GBM - Showed improved survival vs. historical controls |
Challenges:
| |||
| Agent | Mechanism | Clinical Phase/Population | Key Findings and Rationale |
|---|---|---|---|
| Vorinostat | HDAC inhibitor; alters gene expression, induces apoptosis | Phase II | - Limited efficacy as monotherapy - Combining with RT or chemotherapy being explored |
| Azacitidine | DNMT inhibitor; demethylates DNA to restore tumor suppressor genes | Phase II (NCT03666559) | - Ongoing; rationale is that epigenetic changes in GBM may re-sensitize to therapy |
Challenges:
| |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, J.; Karbhari, N.; Campian, J.L. Therapeutic Targets in Glioblastoma: Molecular Pathways, Emerging Strategies, and Future Directions. Cells 2025, 14, 494. https://doi.org/10.3390/cells14070494
Tang J, Karbhari N, Campian JL. Therapeutic Targets in Glioblastoma: Molecular Pathways, Emerging Strategies, and Future Directions. Cells. 2025; 14(7):494. https://doi.org/10.3390/cells14070494
Chicago/Turabian StyleTang, Justin, Nishika Karbhari, and Jian L. Campian. 2025. "Therapeutic Targets in Glioblastoma: Molecular Pathways, Emerging Strategies, and Future Directions" Cells 14, no. 7: 494. https://doi.org/10.3390/cells14070494
APA StyleTang, J., Karbhari, N., & Campian, J. L. (2025). Therapeutic Targets in Glioblastoma: Molecular Pathways, Emerging Strategies, and Future Directions. Cells, 14(7), 494. https://doi.org/10.3390/cells14070494