Exploring MicroRNA Biomarkers for Parkinson’s Disease from mRNA Expression Profiles



Abstract

1. Introduction
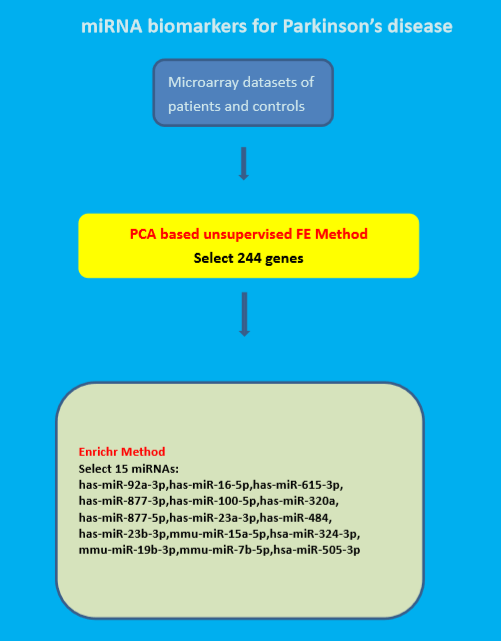
2. Materials and Methods
2.1. Gene Expression
2.2. Principal Component Analysis Based Unsupervised Feature Extraction
2.3. Validation of Obtained mRNAs
3. Results
4. Discussion
More Brain Synapse-Related Biological Terms Are Enriched
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- DeMaagd, G.; Philip, A. Parkinson’s disease and its management: Part 1: Disease entity, risk factors, pathophysiology, clinical presentation, and diagnosis. Pharm. Ther. 2015, 40, 504. [Google Scholar]
- Drui, G.; Carnicella, S.; Carcenac, C.; Favier, M.; Bertrand, A.; Boulet, S.; Savasta, M. Loss of dopaminergic nigrostriatal neurons accounts for the motivational and affective deficits in Parkinson’s disease. Mol. Psychiatry 2014, 19, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Neikrug, A.B.; Maglione, J.E.; Liu, L.; Natarajan, L.; Avanzino, J.A.; Corey-Bloom, J.; Palmer, B.W.; Loredo, J.S.; Ancoli-Israel, S. Effects of sleep disorders on the non-motor symptoms of Parkinson disease. J. Clin. Sleep Med. 2013, 9, 1119–1129. [Google Scholar] [CrossRef] [PubMed]
- Marsh, L. Depression and Parkinson’s disease: Current knowledge. Curr. Neurol. Neurosci. Rep. 2013, 13, 409. [Google Scholar] [CrossRef] [PubMed]
- Watson, G.S.; Leverenz, J.B. Profile of cognitive impairment in Parkinson’s disease. Brain Pathol. 2010, 20, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Levine, C.B.; Fahrbach, K.R.; Siderowf, A.D.; Estok, R.P.; Ludensky, V.M.; Ross, S.D. Diagnosis and treatment of Parkinson’s disease: A systematic review of the literature. Evid. Rep. Technol. Assess. 2003, 57, 1–4. [Google Scholar]
- Tan, E.-K.; Jankovic, J. Genetic testing in Parkinson disease: Promises and pitfalls. Arch. Neurol. 2006, 63, 1232–1237. [Google Scholar] [CrossRef]
- Zimprich, A.; Biskup, S.; Leitner, P.; Lichtner, P.; Farrer, M.; Lincoln, S.; Kachergus, J.; Hulihan, M.; Uitti, R.J.; Calne, D.B.; et al. Mutations in LRRK2 cause autosomal-dominant parkinsonism with pleomorphic pathology. Neuron 2004, 44, 601–607. [Google Scholar] [CrossRef]
- Gasser, T. Usefulness of genetic testing in PD and PD trials: A balanced review. J. Park. Dis. 2015, 5, 209–215. [Google Scholar] [CrossRef]
- Stefanis, L. α-Synuclein in Parkinson’s disease. Cold Spring Harbor Perspect. Med. 2012, 2, a009399. [Google Scholar] [CrossRef]
- Dawson, T.M.; Dawson, V.L. The role of parkin in familial and sporadic Parkinson’s disease. Mov. Disord. 2010, 25, S32–S39. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.K.; Puong, K.Y.; Fook-Chong, S.; Chua, E.; Shen, H.; Yuen, Y.; Pavanni, R.; Wong, M.-C.; Puvan, K.; Zhao, Y. Case-control study of UCHL1 S18Y variant in Parkinson’s disease. Mov. Disord. 2010, 25, S32–S39. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.A.; Andruska, K.M.; Benitez, B.A.; Racette, B.A.; Perlmutter, J.S.; Cruchaga, C. Variants in GBA, SNCA, and MAPT influence Parkinson disease risk, age at onset, and progression. Neurobiol. Aging 2016, 37, 209. e1–209. e7. [Google Scholar] [CrossRef] [PubMed]
- Pinho, R.; Guedes, L.C.; Soreq, L.; Lobo, P.P.; Mestre, T.; Coelho, M.; Rosa, M.M.; Gonçalves, N.; Wales, P.; Mendes, T. Gene expression differences in peripheral blood of Parkinson’s disease patients with distinct progression profiles. PLoS ONE 2016, 11, e0157852. [Google Scholar] [CrossRef]
- Aguiar, P.M.C.; Severino, P. Biomarkers in Parkinson Disease: global gene expression analysis in peripheral blood from patients with and without mutations in PARK2 and PARK8. Einstein 2010, 8, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, Y.H.; Wang, H. Exploring microRNA Biomarker for Amyotrophic Lateral Sclerosis. Int. J. Mol. Sci. 2018, 19, 1318. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, Y.H.; Wang, H. Genetic association between amyotrophic lateral sclerosis and cancer. Genes 2017, 8, 243. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinf. 2013, 14, 128. [Google Scholar] [CrossRef]
- Leggio, L.; Vivarelli, S.; L’Episcopo, F.; Tirolo, C.; Caniglia, S.; Testa, N.; Marchetti, B.; Iraci, N. microRNAs in Parkinson’s disease: From pathogenesis to novel diagnostic and therapeutic approaches. Int. J. Mol. Sci. 2017, 18, 2698. [Google Scholar] [CrossRef]
- Chatterjee, P.; Bhattacharyya, M.; Bandyopadhyay, S.; Roy, D. Studying the system-level involvement of microRNAs in Parkinson’s disease. PLoS ONE 2014, 9, e93751. [Google Scholar] [CrossRef] [PubMed]
- Hoss, A.G.; Labadorf, A.; Beach, T.G.; Latourelle, J.C.; Myers, R.H. microRNA profiles in Parkinson’s disease prefrontal cortex. Front. Aging Neurosci. 2016, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Hoss, A.G.; Kartha, V.K.; Dong, X.; Latourelle, J.C.; Dumitriu, A.; Hadzi, T.C.; Macdonald, M.E.; Gusella, J.F.; Akbarian, S.; Chen, J.F.; et al. MicroRNAs located in the Hox gene clusters are implicated in huntington’s disease pathogenesis. PLoS Genet. 2014, 10, e1004188. [Google Scholar] [CrossRef] [PubMed]
- Sibley, C.R.; Seow, Y.; Curtis, H.; Weinberg, M.S.; Wood, M.J. Silencing of Parkinson’s disease-associated genes with artificial mirtron mimics of miR-1224. Nucleic Acids Res. 2012, 40, 9863–9875. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, J.; Lu, J.; Cao, S.; Zhao, Q.; Yu, Z. Identification of aberrant circulating miRNAs in Parkinson’s disease plasma samples. Brain Behav. 2018, 8, e00941. [Google Scholar] [CrossRef]
- Heman-Ackah, S.M.; Hallegger, M.; Rao, M.S.; Wood, M.J. RISC in PD: The impact of microRNAs in Parkinson’s disease cellular and molecular pathogenesis. Front. Mol. Neurosci. 2013, 6, 40. [Google Scholar] [CrossRef]
- Prajapati, P.; Sripada, L.; Singh, K.; Bhatelia, K.; Singh, R.; Singh, R. TNF-α regulates miRNA targeting mitochondrial complex-I and induces cell death in dopaminergic cells. Biochim. Biophys. Acta Mol. Basis Dis. 2015, 1852, 451–461. [Google Scholar] [CrossRef]
- Vallelunga, A.; Ragusa, M.; Di Mauro, S.; Iannitti, T.; Pilleri, M.; Biundo, R.; Weis, L.; Di Pietro, C.; De Iuliis, A.; Nicoletti, A. Identification of circulating microRNAs for the differential diagnosis of Parkinson’s disease and Multiple System Atrophy. Front. Cell. Neurosci. 2014, 8, 156. [Google Scholar] [CrossRef]
- Khoo, S.K.; Petillo, D.; Kang, U.J.; Resau, J.H.; Berryhill, B.; Linder, J.; Forsgren, L.; Neuman, L.A.; Tan, A.C. Plasma-based circulating MicroRNA biomarkers for Parkinson’s disease. J. Park. Dis. 2012, 2, 321–331. [Google Scholar]
- Martin, I.; Kim, J.W.; Lee, B.D.; Kang, H.C.; Xu, J.C.; Jia, H.; Stankowski, J.; Kim, M.S.; Zhong, J.; Kumar, M.; et al. Ribosomal protein s15 phosphorylation mediates LRRK2 neurodegeneration in Parkinson’s disease. Cell 2014, 157, 472–485. [Google Scholar] [CrossRef]
- Härtlova, A.; Herbst, S.; Peltier, J.; Rodgers, A.; Bilkei-Gorzo, O.; Fearns, A.; Dill, B.D.; Lee, H.; Flynn, R.; Cowley, S.A. LRRK2 is a negative regulator of Mycobacterium tuberculosis phagosome maturation in macrophages. EMBO J. 2018, 37, e98694. [Google Scholar] [CrossRef] [PubMed]
- Esposito, G.; Clara, F.A.; Verstreken, P. Synaptic vesicle trafficking and Parkinson’s disease. Dev. Neurobiol. 2012, 72, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Siebert, M.; Westbroek, W.; Chen, Y.-C.; Moaven, N.; Li, Y.; Velayati, A.; Saraiva-Pereira, M.L.; Martin, S.E.; Sidransky, E. Identification of miRNAs that modulate glucocerebrosidase activity in Gaucher disease cells. RNA Biol. 2014, 11, 1291–1300. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Control | PD | |
|---|---|---|
| Control | 24 | 8 |
| PD | 3 | 22 |
| Term | Overlap | p-Value | Adjusted p-Value |
|---|---|---|---|
| Parkinson’s disease DOID-14330 human GSE19587 sample 740 | 65/207 | 5.02 × | 4.18 × |
| Parkinson’s disease DOID-14330 human GSE19587 sample 1080 | 56/167 | 5.88 × | 1.60 × |
| Parkinson’s disease DOID-14330 human GSE19587 sample 496 | 73/361 | 3.90 × | 1.59 × |
| Parkinson’s disease DOID-14330 human GSE7621 sample 940 | 67/365 | 2.96 × | 6.06 × |
| Dystonia C0393593 human GSE3064 sample 329 | 62/317 | 1.06 × | 1.74 × |
| Term | Overlap | p-Value | Adjusted p-value |
|---|---|---|---|
| Parkinson’s disease DOID-14330 human GSE19587 sample 741 | 33/158 | 5.14 × | 1.22 × |
| Parkinson’s disease DOID-14330 human GSE7621 sample 940 | 35/235 | 1.05 × | 1.74 × |
| Parkinson’s disease DOID-14330 human GSE7621 sample 941 | 38/342 | 1.55 × | 2.19 × |
| Parkinson’s disease DOID-14330 human GSE19587 sample 1080 | 37/433 | 1.17 × | 8.78 × |
| Parkinson’s disease DOID-14330 human GSE6613 sample 788 | 26/274 | 1.24 × | 5.50 × |
| Parkinson’s disease DOID-14330 human GSE19587 sample 496 | 15/239 | 1.03 × | 2.15 × |
| Term | Overlap | p-Value | Adjusted p-Value |
|---|---|---|---|
| Ribosome_Homo sapiens_hsa03010 | 28/137 | 1.68 × | 2.92 × |
| Phagosome_Homo sapiens_hsa04145 | 16/154 | 1.72 × | 1.49 × |
| Synaptic vesicle cycle_Homo sapiens_hsa04721 | 10/63 | 4.11 × | 2.38 × |
| Pathogenic Escherichia coli infection_Homo sapiens_hsa05130 | 9/55 | 2.38 × | 1.04 × |
| Gap junction_Homo sapiens_hsa04540 | 10/88 | 1.18 × | 4.11 × |
| Mineral absorption_Homo sapiens_hsa04978 | 8/51 | 2.68 × | 7.76 × |
| Oxidative phosphorylation_Homo sapiens_hsa00190 | 10/133 | 6.09 × | 1.51 × |
| Parkinson’s disease_Homo sapiens_hsa05012 | 10/142 | 1.11 × | 2.42 × |
| Vibrio cholerae infection_Homo sapiens_hsa05110 | 6/51 | 8.84 × | 1.71 × |
| GABAergic synapse_Homo sapiens_hsa04727 | 7/88 | 2.15 × | 3.73 × |
| Term | Overlap | p-Value | Adjusted p-Value | Reference |
|---|---|---|---|---|
| hsa-miR-92a-3p | 37/1404 | 1.41 × 10−8 | 2.71 × | [21] |
| hsa-miR-16-5p | 37/1555 | 1.93 × 10−7 | 1.85 × | [22] |
| hsa-miR-615-3p | 25/891 | 1.38 × 10−6 | 8.85 × | [23] |
| hsa-miR-877-3p | 19/606 | 5.92 × 10−6 | 2.28 × | [24] |
| hsa-miR-100-5p | 12/250 | 5.37× 10−6 | 2.28 × | [25] |
| hsa-miR-320a | 18/584 | 1.33 × 10−5 | 4.25 × | [26] |
| hsa-miR-877-5p | 11/235 | 1.68 × 10−5 | 4.63 × | [24] |
| hsa-miR-23a-3p | 11/249 | 2.88 × 10−5 | 6.91 × | [25] |
| hsa-miR-484 | 22/890 | 4.37 × 10−5 | 9.33 × | [25] |
| hsa-miR-23b-3p | 12/322 | 6.55 × 10−5 | 1.26 × | [27] |
| mmu-miR-15a-5p | 15/499 | 9.42 × 10−5 | 1.65 × | [25] |
| hsa-miR-324-3p | 12/338 | 1.04 × 10−4 | 1.66 × | [28] |
| mmu-miR-19b-3p | 11/310 | 2.03 × 10−4 | 3.00 × | [20] |
| mmu-miR-7b-5p | 13/438 | 3.13 × 10−4 | 4.02 × | [20] |
| hsa-miR-505-3p | 9/222 | 2.93 × 10−4 | 4.02 × | [29] |
| Name | Overlap | p-Value | Adjusted p-Value |
|---|---|---|---|
| LRRK2 Gly2019Ser (G2019S) mutation knockin human GSE36321 sample 1688 | 21/335 | 1.03 × | 4.82 × |
| LRRK2 mutant human GSE33298 sample 2039 | 16/309 | 4.94 × | 1.55 × |
| LRRK2 dominant negative mutation-G2019S homozygous human GSE33298 sample 1743 | 12/280 | 1.68 × | 4.14 × |
| LRRK2 dominant negative mutation-G2019S homozygous human GSE33298 sample 1741 | 12/337 | 1.01 × | 2.33 × |
| Name | Overlap | p-Value | Adjusted p-Value |
|---|---|---|---|
| LRRK2 Gly2019Ser (G2019S) mutation knockin human GSE36321 sample 1688 | 24/265 | 8.38 × | 9.78 × |
| LRRK2 dominant negative mutation-G2019S heterozygous human GSE33298 sample 1739 | 9/282 | 1.61 × | 3.56 × |
| Term | Overlap | p-Value | Adjusted p-Value | |
|---|---|---|---|---|
| Oxcarbazepine-1600-mg/kg-in_CMC-Rat-Brain-3d-dn | 31/369 | 1.66 × | 1.31 × | |
| Carbachol-15-mg/kg_in_Water-Rat-Brain-3d-up | 26/318 | 4.96 × | 7.82 × | |
| Piracetam-2500_mg/kg_in_CMC-Rat-Brain-5d-up | 27/325 | 7.89 × | 2.56 × | |
| Theophylline-225_mg/kg_in_Water-Rat-Brain-3d-dn | 25/314 | 3.87 × | 5.08 × | |
| Tramadol-114_mg/kg_in_Water-Rat-Brain-5d-dn | 26/315 | 3.93 × | 7.75 × | |
| Term | Overlap | p-Value | Adjusted p-Value |
|---|---|---|---|
| GTEX-X585-0011-R2B-SM-46MVF_brain_male_50-59_years | 81/1895 | 1.63 × | 4.72 × |
| GTEX-WHSE-0011-R2A-SM-3P5ZL_brain_male_20-29_years | 71/1660 | 7.67 × | 1.11 × |
| GTEX-X261-0011-R8A-SM-4E3I5_brain_male_50-59_years | 70/1878 | 8.35 × | 8.08 × |
| GTEX-N7MT-0011-R10A-SM-2I3E1_brain_female_60-69_years | 70/1918 | 2.88 × | 2.09 × |
| GTEX-TSE9-0011-R8A-SM-3DB7R_brain_female_60-69_years | 62/1548 | 2.52 × | 1.47 × |
| Term | Overlap | p-Value | Adjusted p-Value |
|---|---|---|---|
| GTEX-S4Q7-1226-SM-4AD5I_testis_male_20-29_years | 22/329 | 8.84 × | 2.36 × |
| GTEX-U4B1-1526-SM-4DXSL_testis_male_40-49_years | 20/282 | 3.48 × | 3.09 × |
| GTEX-UPK5-1426-SM-4JBHH_liver_male_40-49_years | 79/3879 | 2.06 × | 2.74 × |
| GTEX-OHPM-2126-SM-3LK75_testis_male_50-59_years | 26/525 | 6.48 × | 4.07 × |
| GTEX-S7PM-0626-SM-4AD4Q_testis_male_60-69_years | 34/911 | 7.63 × | 4.07 × |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taguchi, Y.-h.; Wang, H. Exploring MicroRNA Biomarkers for Parkinson’s Disease from mRNA Expression Profiles. Cells 2018, 7, 245. https://doi.org/10.3390/cells7120245
Taguchi Y-h, Wang H. Exploring MicroRNA Biomarkers for Parkinson’s Disease from mRNA Expression Profiles. Cells. 2018; 7(12):245. https://doi.org/10.3390/cells7120245
Chicago/Turabian StyleTaguchi, Y-h., and Hsiuying Wang. 2018. "Exploring MicroRNA Biomarkers for Parkinson’s Disease from mRNA Expression Profiles" Cells 7, no. 12: 245. https://doi.org/10.3390/cells7120245
APA StyleTaguchi, Y.-h., & Wang, H. (2018). Exploring MicroRNA Biomarkers for Parkinson’s Disease from mRNA Expression Profiles. Cells, 7(12), 245. https://doi.org/10.3390/cells7120245