Differences in the Emission of Volatile Organic Compounds (VOCs) between Non-Differentiating and Adipogenically Differentiating Mesenchymal Stromal/Stem Cells from Human Adipose Tissue

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
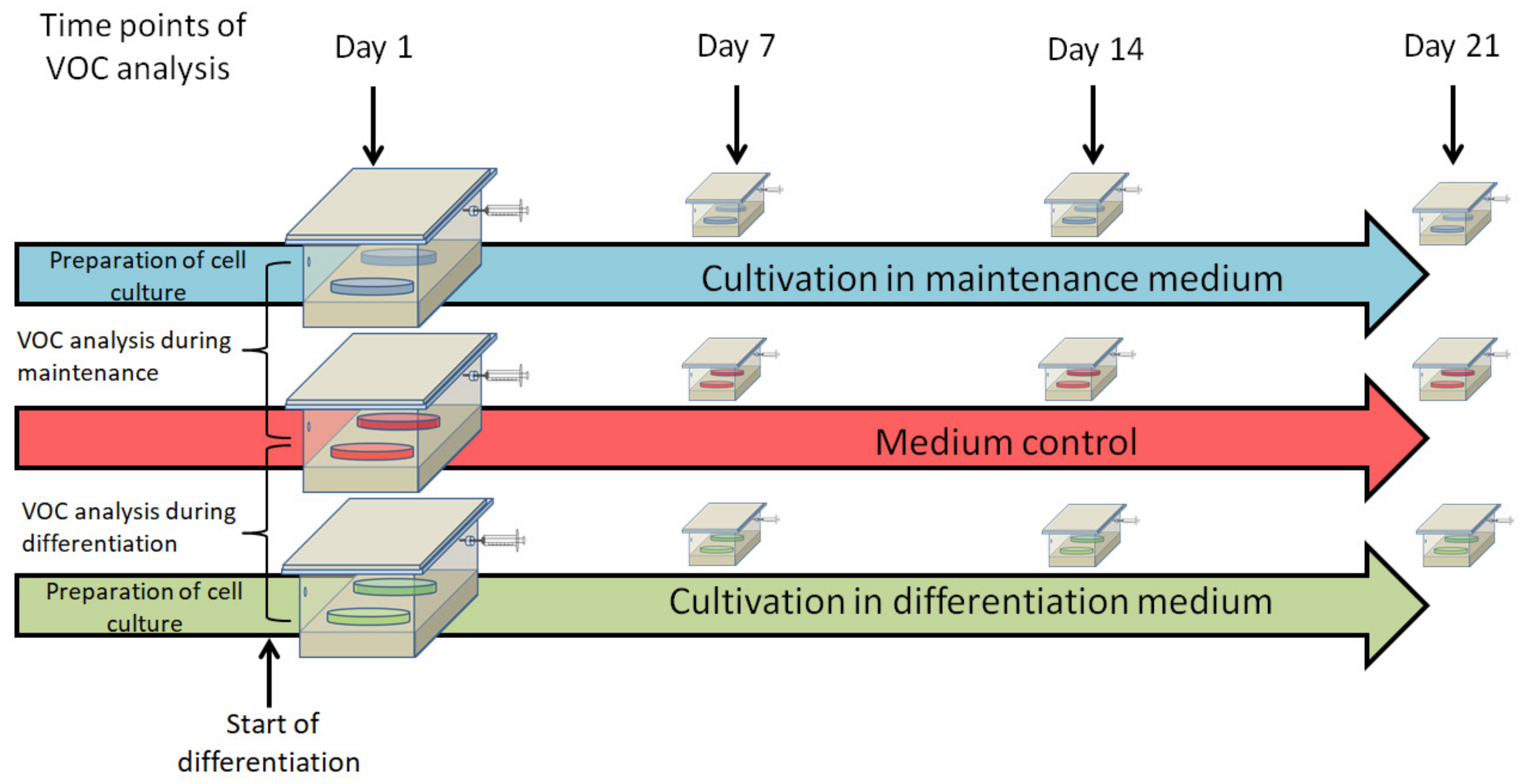
2.1. Isolation, Cultivation, and Differentiation of Adipose Tissue-Derived Mesenchymal Stromal/Stem Cells (ASCs)
2.2. Analysis of Cell Numbers
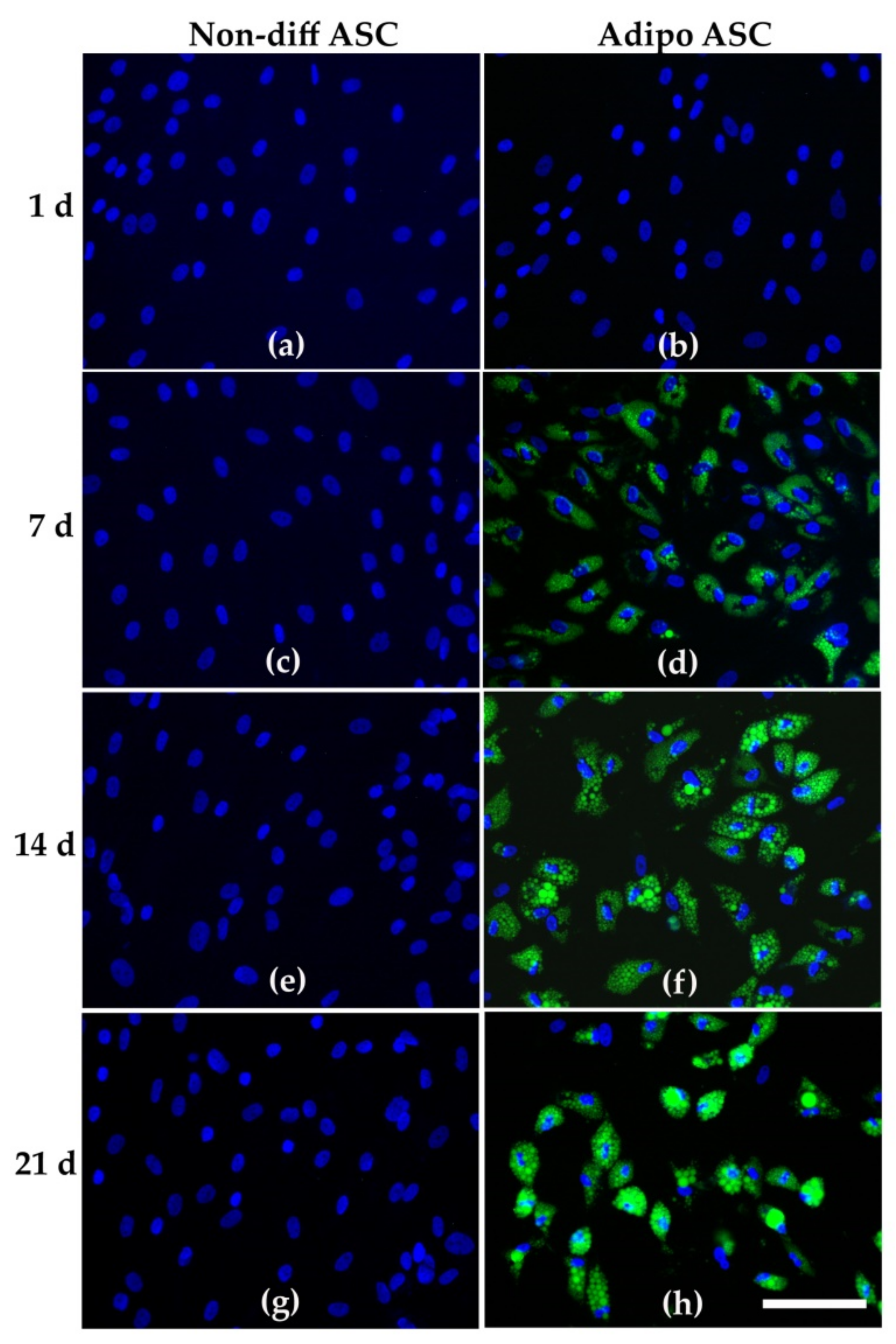
2.3. Analysis of Adipogenic Differentiation
2.4. Volatile Organic Compound (VOC) Sampling by Means of Needle Trap Micro Extraction
2.5. Gas Chromatography and Mass Spectrometry (GC-MS) Analysis
2.6. Reference Substances
2.7. Statistical Analysis
3. Results
3.1. Adipogenic Differentiation of ASCs
3.2. Comparision of VOC Emissions of Non-Differentiating and Adipogenically Differentiating ASCs
3.3. Comparision of VOC Emissions from Medium Control and Corresponding Cell Cultures
3.3.1. VOC Consumption during Adipogenic Differentiation
3.3.2. Culture Medium-Dependent VOCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Valtieri, M.; Sorrentino, A. The mesenchymal stromal cell contribution to homeostasis. J. Cell. Physiol. 2008, 217, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.H.; Kim, B.; Choi, I.; Kim, H.; Choi, H.S.; Suh, K.; Bae, Y.C.; Jung, J.S. Characterization and expression analysis of mesenchymal stem cells from human bone marrow and adipose tissue. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2004, 14, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Peroni, D.; Scambi, I.; Pasini, A.; Lisi, V.; Bifari, F.; Krampera, M.; Rigotti, G.; Sbarbati, A.; Galiè, M. Stem molecular signature of adipose-derived stromal cells. Exp. Cell Res. 2008, 314, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Spitkovsky, D.; Hescheler, J. Adult mesenchymal stromal stem cells for therapeutic applications. Minim. Invasive Ther. Allied Technol. MITAT Off. J. Soc. Minim. Invasive Ther. 2008, 17, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Ashjian, P.; De Ugarte, D.A.; Huang, J.I.; Mizuno, H.; Alfonso, Z.C.; Fraser, J.K.; Benhaim, P.; Hedrick, M.H. Human adipose tissue is a source of multipotent stem cells. Mol. Biol. Cell 2002, 13, 4279–4295. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef]
- De Ugarte, D.A.; Morizono, K.; Elbarbary, A.; Alfonso, Z.; Zuk, P.A.; Zhu, M.; Dragoo, J.L.; Ashjian, P.; Thomas, B.; Benhaim, P.; et al. Comparison of multi-lineage cells from human adipose tissue and bone marrow. Cells Tissues Organs 2003, 174, 101–109. [Google Scholar] [CrossRef]
- Schop, D.; Janssen, F.W.; van Rijn, L.D.S.; Fernandes, H.; Bloem, R.M.; de Bruijn, J.D.; van Dijkhuizen-Radersma, R. Growth, metabolism, and growth inhibitors of mesenchymal stem cells. Tissue Eng. Part A 2009, 15, 1877–1886. [Google Scholar] [CrossRef]
- Pattappa, G.; Heywood, H.K.; de Bruijn, J.D.; Lee, D.A. The metabolism of human mesenchymal stem cells during proliferation and differentiation. J. Cell. Physiol. 2011, 226, 2562–2570. [Google Scholar] [CrossRef]
- Li, Q.; Gao, Z.; Chen, Y.; Guan, M.-X. The role of mitochondria in osteogenic, adipogenic and chondrogenic differentiation of mesenchymal stem cells. Protein Cell 2017, 8, 439–445. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Marsboom, G.; Toth, P.T.; Rehman, J. Mitochondrial respiration regulates adipogenic differentiation of human mesenchymal stem cells. PLoS ONE 2013, 8, e77077. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.; Salamon, A.; Mispagel, S.; Kamp, G.; Peters, K. Energy metabolic capacities of human adipose-derived mesenchymal stromal cells in vitro and their adaptations in osteogenic and adipogenic differentiation. Exp. Cell Res. 2018, 370, 632–642. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.; Engelmann, R.; Kamp, G.; Peters, K. Human adipocytes and CD34+ cells from the stromal vascular fraction of the same adipose tissue differ in their energy metabolic enzyme configuration. Exp. Cell Res. 2019, 380, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, J.; Ruohola-Baker, H. Metabolic remodeling during the loss and acquisition of pluripotency. Development 2017, 144, 541–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capuano, R.; Spitalieri, P.; Talarico, R.V.; Domakoski, A.C.; Catini, A.; Paolesse, R.; Martinelli, E.; Novelli, G.; Sangiuolo, F.; Di Natale, C. A preliminary analysis of volatile metabolites of human induced pluripotent stem cells along the in vitro differentiation. Sci. Rep. 2017, 7, 1621. [Google Scholar] [CrossRef] [PubMed]
- Filipiak, W.; Sponring, A.; Mikoviny, T.; Ager, C.; Schubert, J.; Miekisch, W.; Amann, A.; Troppmair, J. Release of volatile organic compounds (VOCs) from the lung cancer cell line CALU-1 in vitro. Cancer Cell Int. 2008, 8. [Google Scholar] [CrossRef]
- Filipiak, W.; Sponring, A.; Baur, M.M.; Ager, C.; Filipiak, A.; Wiesenhofer, H.; Nagl, M.; Troppmair, J.; Amann, A. Characterization of volatile metabolites taken up by or released from Streptococcus pneumoniae and Haemophilus influenzae by using GC-MS. Microbiology 2012, 158, 3044–3053. [Google Scholar] [CrossRef] [Green Version]
- Kai, M.; Haustein, M.; Molina, F.; Petri, A.; Scholz, B.; Piechulla, B. Bacterial volatiles and their action potential. Appl. Microbiol. Biotechnol. 2009, 81, 1001–1012. [Google Scholar] [CrossRef]
- Trefz, P.; Rösner, L.; Hein, D.; Schubert, J.K.; Miekisch, W. Evaluation of needle trap micro-extraction and automatic alveolar sampling for point-of-care breath analysis. Anal. Bioanal. Chem. 2013, 405, 3105–3115. [Google Scholar] [CrossRef]
- Bergmann, A.; Trefz, P.; Fischer, S.; Klepik, K.; Walter, G.; Steffens, M.; Ziller, M.; Schubert, J.K.; Reinhold, P.; Köhler, H.; et al. In Vivo Volatile Organic Compound Signatures of Mycobacterium avium subsp. paratuberculosis. PLoS ONE 2015, 10, e0123980. [Google Scholar] [CrossRef]
- Filipiak, W.; Mochalski, P.; Filipiak, A.; Ager, C.; Cumeras, R.; Davis, C.E.; Agapiou, A.; Unterkofler, K.; Troppmair, J. A Compendium of Volatile Organic Compounds (VOCs) Released By Human Cell Lines. Curr. Med. Chem. 2016, 23, 2112–2131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trefz, P.; Koehler, H.; Klepik, K.; Moebius, P.; Reinhold, P.; Schubert, J.K.; Miekisch, W. Volatile Emissions from Mycobacterium avium subsp. paratuberculosis Mirror Bacterial Growth and Enable Distinction of Different Strains. PLoS ONE 2013, 8, e76868. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, C.A.; Sanchez, E.Y.; Reyes, J.G.; Young, M.E. Volatile profiles of human skin cell cultures in different degrees of senescence. J. Chromatogr. B Analyt. Technol. Biomed. Life. Sci. 2010, 878, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, C.A.; Sánchez, E.Y.; Reyes, J.G.; Young, M.E. Volatile organic compounds produced by human skin cells. Biol. Res. 2007, 40, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Wang, T.; Spanĕl, P. On-line, simultaneous quantification of ethanol, some metabolites and water vapour in breath following the ingestion of alcohol. Physiol. Meas. 2002, 23, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Miekisch, W.; Schubert, J.K.; Noeldge-Schomburg, G.F.E. Diagnostic potential of breath analysis--focus on volatile organic compounds. Clin. Chim. Acta Int. J. Clin. Chem. 2004, 347, 25–39. [Google Scholar] [CrossRef]
- Bischoff, A.-C.; Oertel, P.; Sukul, P.; Rimmbach, C.; David, R.; Schubert, J.; Miekisch, W. Smell of cells: Volatile profiling of stem- and non-stem cell proliferation. J. Breath Res. 2017, 12. [Google Scholar] [CrossRef]
- Traxler, S.; Bischoff, A.-C.; Trefz, P.; Schubert, J.K.; Miekisch, W. Versatile set-up for non-invasive in vitro analysis of headspace VOCs. J. Breath Res. 2018, 12, 041001. [Google Scholar] [CrossRef]
- Mochalski, P.; Sponring, A.; King, J.L.; Unterkofler, K.; Troppmair, J.; Amann, A. Release and uptake of volatile organic compounds by human hepatocellular carcinoma cells (HepG2) in vitro. Cancer Cell Int. 2013, 13, 72. [Google Scholar] [CrossRef]
- Küntzel, A.; Fischer, S.; Bergmann, A.; Oertel, P.; Steffens, M.; Trefz, P.; Miekisch, W.; Schubert, J.K.; Reinhold, P.; Köhler, H. Effects of biological and methodological factors on volatile organic compound patterns during cultural growth of Mycobacterium avium ssp. paratuberculosis. J. Breath Res. 2016, 10, 037103. [Google Scholar] [CrossRef]
- Mochalski, P.; Theurl, M.; Sponring, A.; Unterkofler, K.; Kirchmair, R.; Amann, A. Analysis of volatile organic compounds liberated and metabolised by human umbilical vein endothelial cells (HUVEC) in vitro. Cell Biochem. Biophys. 2015, 71, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Odinokov, A.; Ostroumov, D. Structural Degradation and Swelling of Lipid Bilayer under the Action of Benzene. J. Phys. Chem. B 2015, 119, 15006–15013. [Google Scholar] [CrossRef] [PubMed]
- Michoudet, C.; Baverel, G. Characteristics of acetaldehyde metabolism in isolated dog, rat and guinea-pig kidney tubules. Biochem. Pharmacol. 1987, 36, 3987–3991. [Google Scholar] [CrossRef]
- Nelson, D.L.; Cox, M.M. Lehninger Biochemie; Springer: Berlin, Heidelberg, 2001; ISBN 978-3-662-08290-4. [Google Scholar]
- Kortelainen, M.L.; Huttunen, P.; Hirvonen, J. Histochemical and biochemical detection of alcohol dehydrogenase in rat brown adipose tissue. Alcohol Fayettev. N 1991, 8, 151–154. [Google Scholar] [CrossRef]
- Tang, H.; Lu, Y.; Zhang, L.; Wu, Z.; Hou, X.; Xia, H. Determination of volatile organic compounds exhaled by cell lines derived from hematological malignancies. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef] [PubMed]
- Amann, A.; de Costello, B.L.; Miekisch, W.; Schubert, J.; Buszewski, B.; Pleil, J.; Ratcliffe, N.; Risby, T. The human volatilome: Volatile organic compounds (VOCs) in exhaled breath, skin emanations, urine, feces and saliva. J. Breath Res. 2014, 8, 034001. [Google Scholar] [CrossRef] [PubMed]
- El Masry, A.M.; Smith, J.N.; Williams, R.T. Studies in detoxication. 69. The metabolism of alkylbenzenes: N-propylbenzene and n-butylbenzene with further observations on ethylbenzene. Biochem. J. 1956, 64, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Marchitti, S.A.; Brocker, C.; Stagos, D.; Vasiliou, V. Non-P450 aldehyde oxidizing enzymes: The aldehyde dehydrogenase superfamily. Expert Opin. Drug Metab. Toxicol. 2008, 4, 697–720. [Google Scholar] [CrossRef]
- Marcato, P.; Dean, C.A.; Giacomantonio, C.A.; Lee, P.W.K. Aldehyde dehydrogenase: Its role as a cancer stem cell marker comes down to the specific isoform. Cell Cycle Georget. Tex 2011, 10, 1378–1384. [Google Scholar] [CrossRef]
- Zimmermann, D.; Hartmann, M.; Moyer, M.P.; Nolte, J.; Baumbach, J.I. Determination of volatile products of human colon cell line metabolism by GC/MS analysis. Metabolomics 2007, 3, 13–17. [Google Scholar] [CrossRef]
- Wermuth, B.; Münch, J.D.; von Wartburg, J.P. Purification and properties of NADPH-dependent aldehyde reductase from human liver. J. Biol. Chem. 1977, 252, 3821–3828. [Google Scholar] [PubMed]
- Klyosov, A.A. Kinetics and Specificity of Human Liver Aldehyde Dehydrogenases toward Aliphatic, Aromatic, and Fused Polycyclic Aldehydes †. Biochemistry 1996, 35, 4457–4467. [Google Scholar] [CrossRef] [PubMed]
- Jareño-Esteban, J.J.; Muñoz-Lucas, M.Á.; Carrillo-Aranda, B.; Maldonado-Sanz, J.Á.; de Granda-Orive, I.; Aguilar-Ros, A.; Civera-Tejuca, C.; Gutiérrez-Ortega, C.; Callol-Sánchez, L.M. Volatile Organic Compounds in Exhaled Breath in a Healthy Population: Effect of Tobacco Smoking. Arch. Bronconeumol. Engl. Ed. 2013, 49, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Kneepkens, C.M.; Lepage, G.; Roy, C.C. The potential of the hydrocarbon breath test as a measure of lipid peroxidation. Free Radic. Biol. Med. 1994, 17, 127–160. [Google Scholar] [CrossRef]
- Corradi, M.; Pignatti, P.; Manini, P.; Andreoli, R.; Goldoni, M.; Poppa, M.; Moscato, G.; Balbi, B.; Mutti, A. Comparison between exhaled and sputum oxidative stress biomarkers in chronic airway inflammation. Eur. Respir. J. Off. J. Eur. Soc. Clin. Respir. Physiol. 2004, 24, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Free radicals, antioxidants, and human disease: Curiosity, cause, or consequence? Lancet Lond. Engl. 1994, 344, 721–724. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance | Normalized VOC Concentration [pmol/L per 1 × 106 cells ± SD] | Statistically Significant | |
|---|---|---|---|
| Nondifferentiating ASCs | Adipogenically Differentiating ASCs | ||
| Acetaldehyde | 229.3 (± 34.5) | 686.4 (± 14.4) | Yes |
| Pentane | 64.2 (± 8.7) | 86.9 (± 1.1) | Yes |
| 1,3-bis-(1,1-dimethylethyl) Benzene | 0.368 (± 0.04) | 1.4 (± 0.2) | Yes |
| Ethylbenzene | 0.94 (± 0.1) | 1.9 (± 0.4) | Yes |
| Benzaldehyde | 6.17 (± 2.1) | 5.93 (± 1.2) | No |
| Heptanal | 9.6 (± 1.9) | 25.4 (± 5.1) | Yes |
| Octanal | 11.5 (± 1.1) | 23.8 (± 0.6) | Yes |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klemenz, A.-C.; Meyer, J.; Ekat, K.; Bartels, J.; Traxler, S.; Schubert, J.K.; Kamp, G.; Miekisch, W.; Peters, K. Differences in the Emission of Volatile Organic Compounds (VOCs) between Non-Differentiating and Adipogenically Differentiating Mesenchymal Stromal/Stem Cells from Human Adipose Tissue. Cells 2019, 8, 697. https://doi.org/10.3390/cells8070697
Klemenz A-C, Meyer J, Ekat K, Bartels J, Traxler S, Schubert JK, Kamp G, Miekisch W, Peters K. Differences in the Emission of Volatile Organic Compounds (VOCs) between Non-Differentiating and Adipogenically Differentiating Mesenchymal Stromal/Stem Cells from Human Adipose Tissue. Cells. 2019; 8(7):697. https://doi.org/10.3390/cells8070697
Chicago/Turabian StyleKlemenz, Ann-Christin, Juliane Meyer, Katharina Ekat, Julia Bartels, Selina Traxler, Jochen K. Schubert, Günter Kamp, Wolfram Miekisch, and Kirsten Peters. 2019. "Differences in the Emission of Volatile Organic Compounds (VOCs) between Non-Differentiating and Adipogenically Differentiating Mesenchymal Stromal/Stem Cells from Human Adipose Tissue" Cells 8, no. 7: 697. https://doi.org/10.3390/cells8070697
APA StyleKlemenz, A.-C., Meyer, J., Ekat, K., Bartels, J., Traxler, S., Schubert, J. K., Kamp, G., Miekisch, W., & Peters, K. (2019). Differences in the Emission of Volatile Organic Compounds (VOCs) between Non-Differentiating and Adipogenically Differentiating Mesenchymal Stromal/Stem Cells from Human Adipose Tissue. Cells, 8(7), 697. https://doi.org/10.3390/cells8070697