Vertically-Aligned Functionalized Silicon Micropillars for 3D Culture of Human Pluripotent Stem Cell-Derived Cortical Progenitors

 , , ,
, , ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
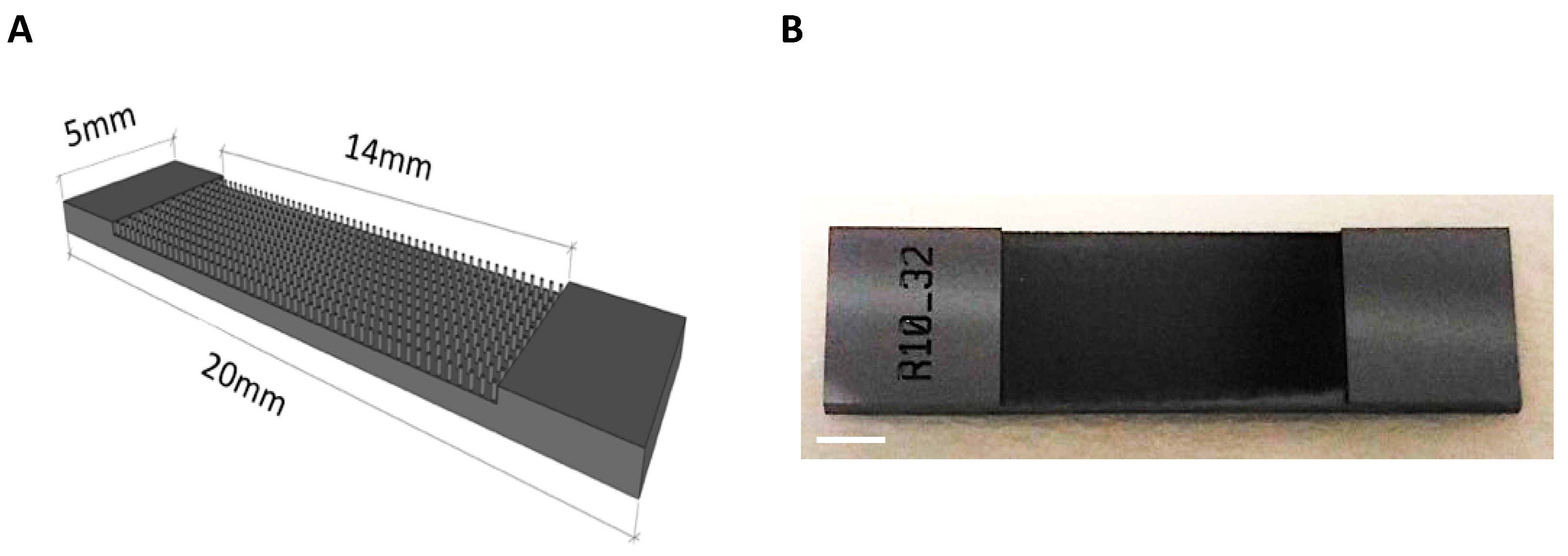
2.1. Vertically-Aligned Silicon Micropillar Array Fabrication
2.2. Cell Cultures
2.3. Scanning Electron Microscopy (SEM)
2.4. Cell Growth/Viability Assay
2.5. Immunocytochemistry
2.6. Time Lapse Analysis
2.7. RNA Isolation and Quantitative RT-PCR (qRT-PCR)
2.8. Statistical Analysis
3. Results and Discussion
3.1. Generation of Vertically-Aligned Silicon Micropillar Array Structures
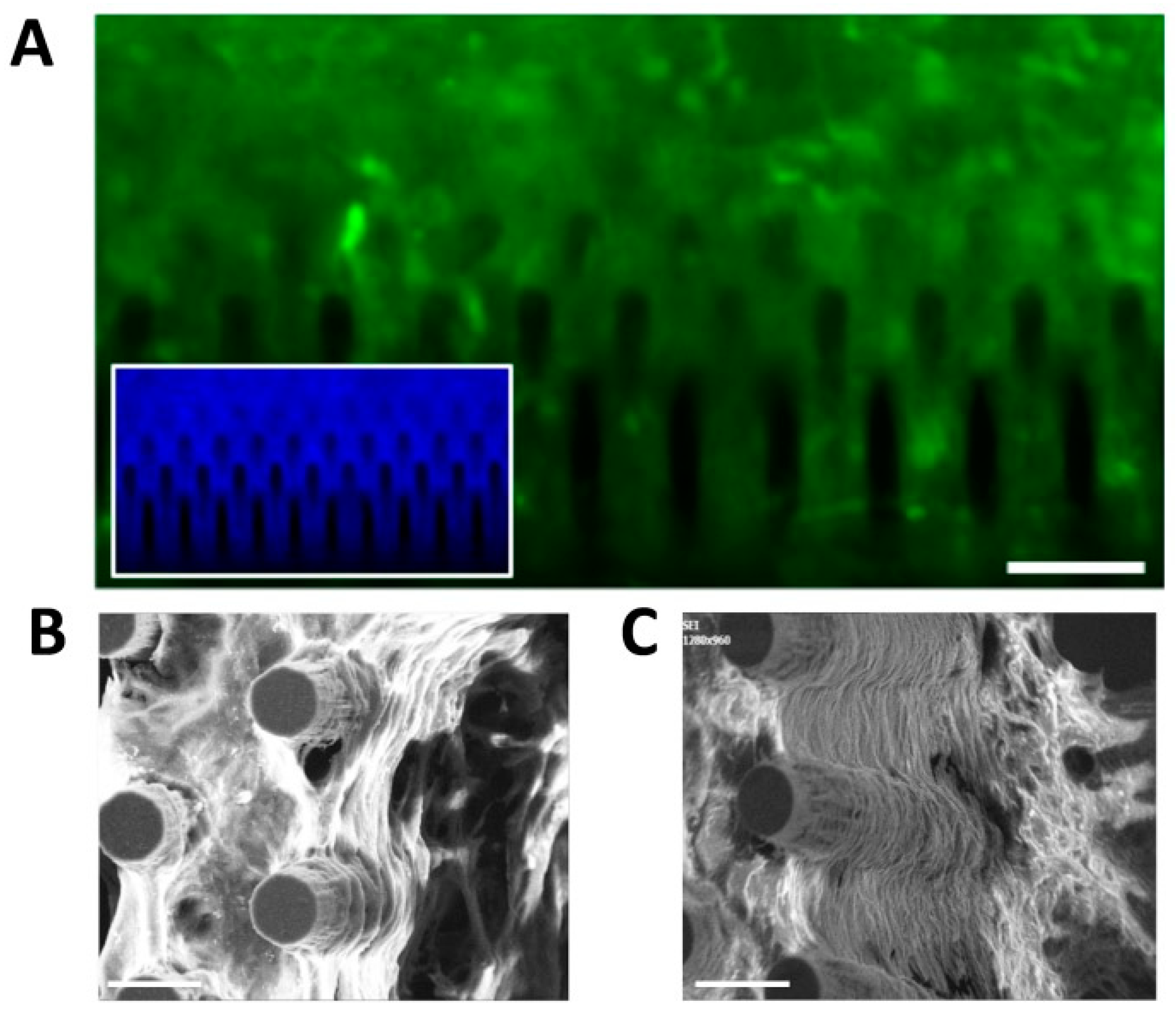
3.2. Culturing Human Cortical Progenitors on Vertically-Aligned Silicon Micropillar Arrays
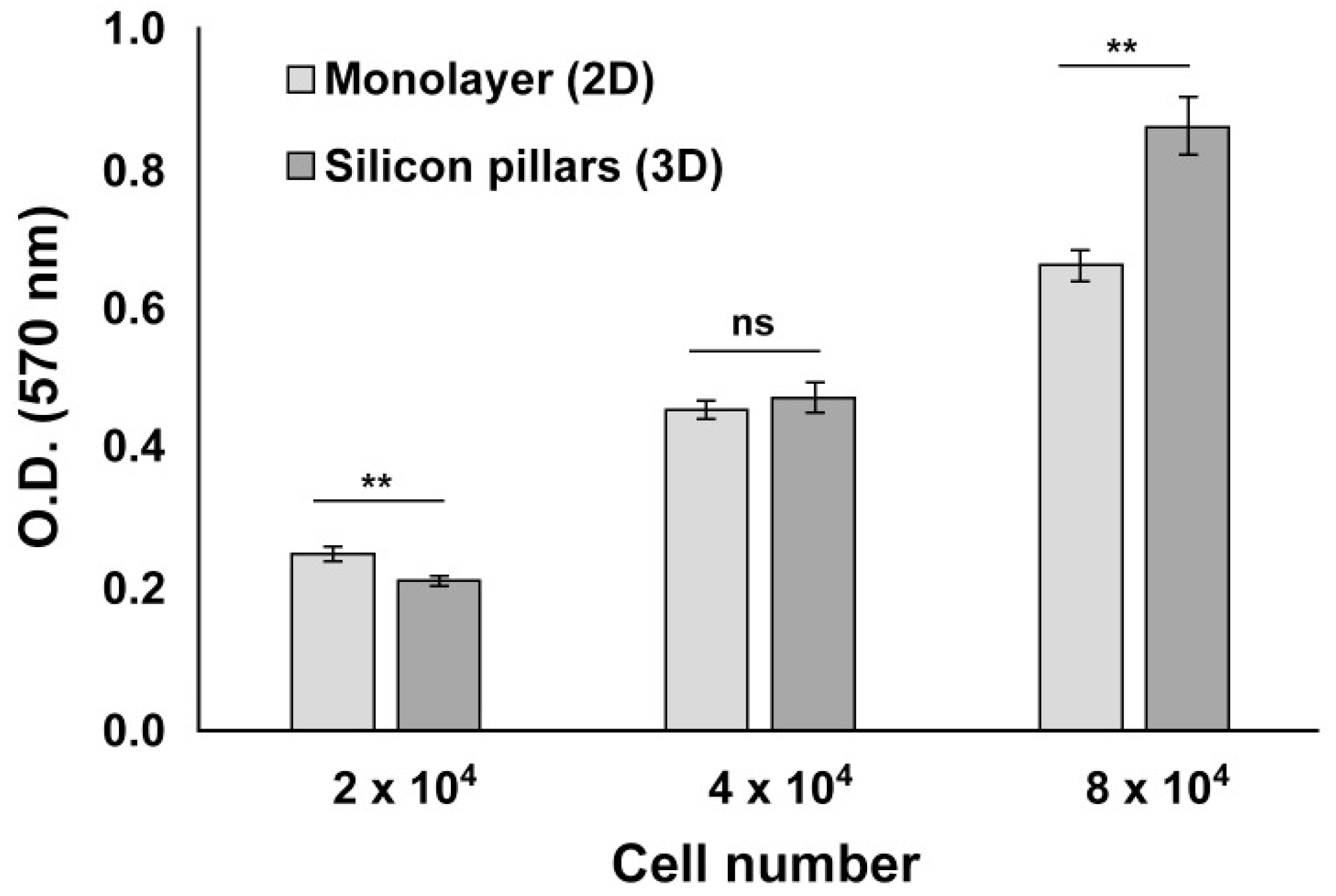
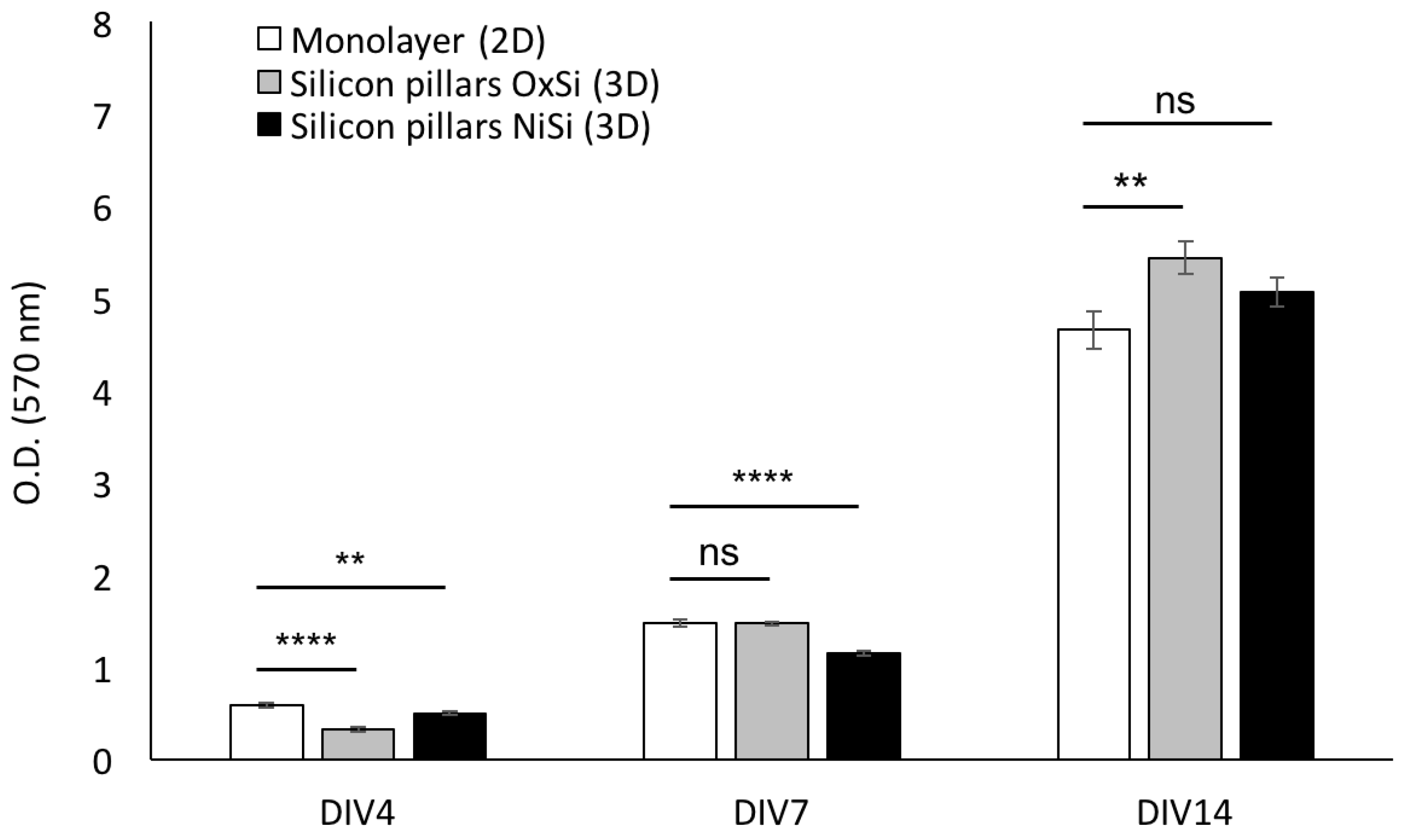
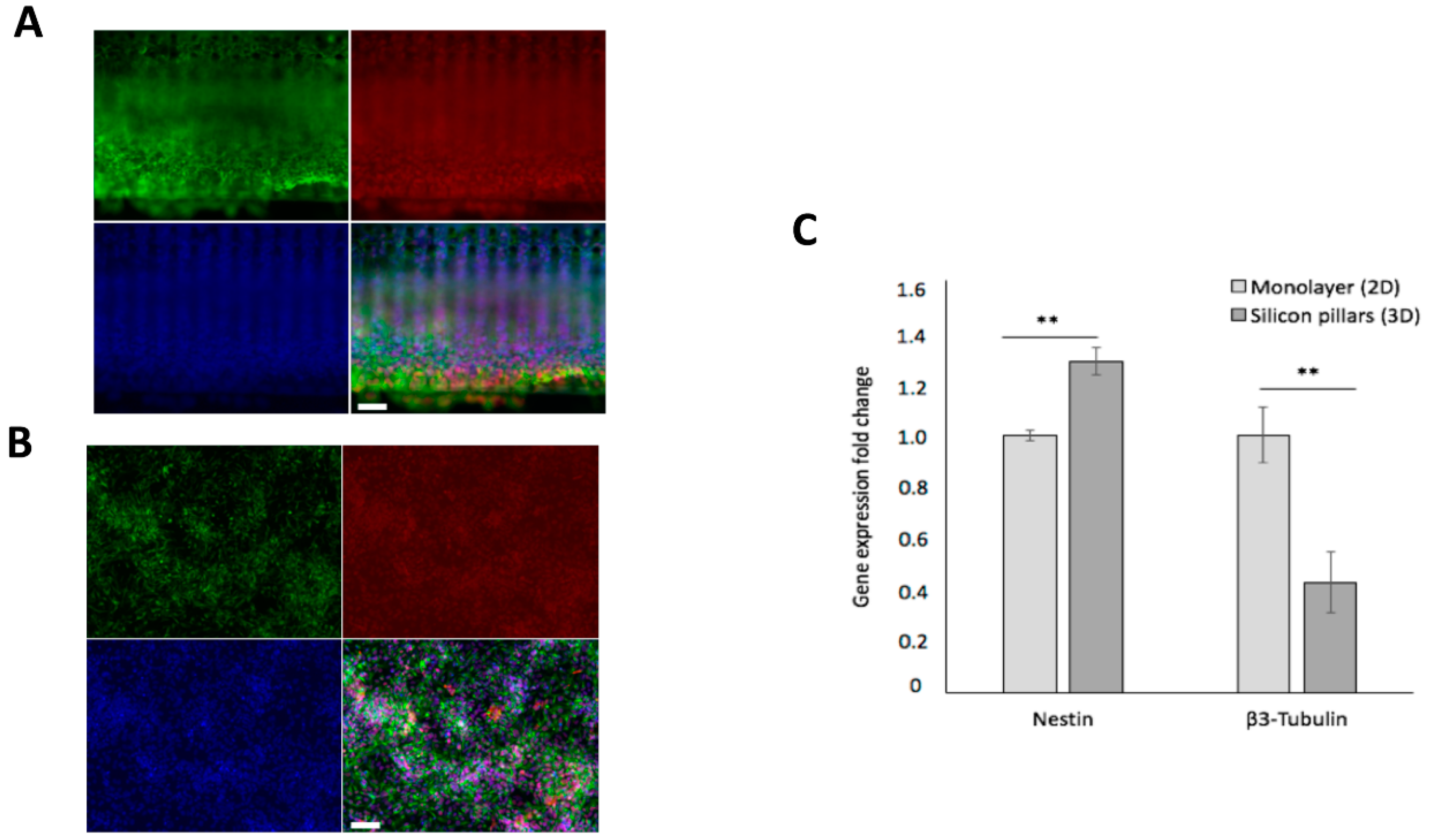
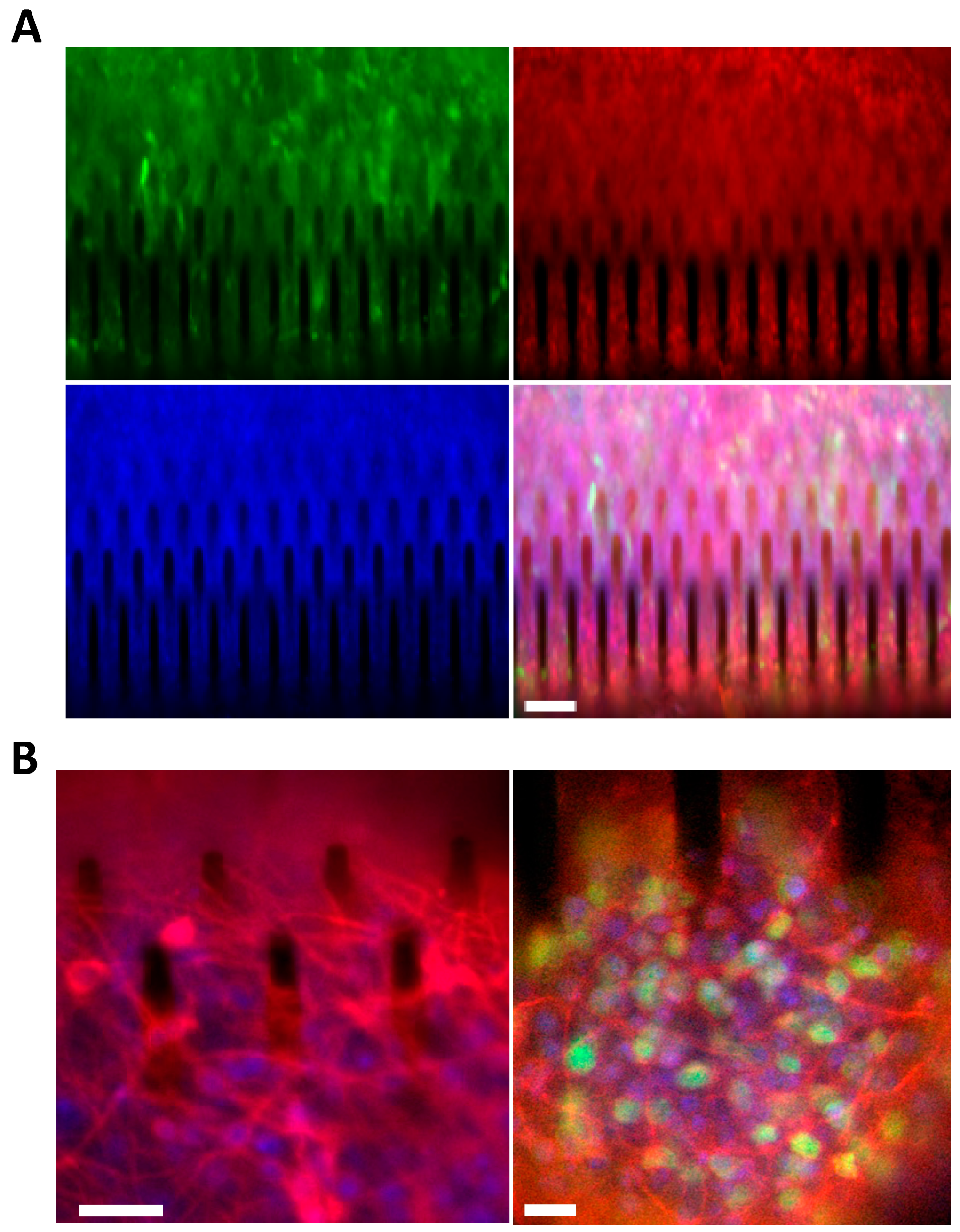
3.3. hCPs Seeded on Silicon Micropillar Arrays Proliferate and form 3D Layered Structures
3.4. hCPs Grown on Silicon Micropillar Arrays Retain Their Multipotency and Regional Identity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
References
- Davenport, R.J. What controls organ regeneration? Science 2005, 309, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffith, L.G.; Swartz, M.A. Capturing complex 3D tissue physiology in vitro. Nat. Rev. Mol. Cell Biol. 2006, 7, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Vergani, L.; Grattarola, M.; Nicolini, C. Modifications of chromatin structure and gene expression following induced alterations of cellular shape. Int. J. Biochem. Cell Biol. 2004, 36, 1447–1461. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.H.; Collier, J.H.; Sfeir, C.S.; Healy, K.E. Engineering gene expression and protein synthesis by modulation of nuclear shape. Proc. Natl. Acad. Sci. USA 2002, 99, 1972–1977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhadriraju, K.; Chen, C.S. Engineering cellular microenvironments to cell-based drug testing improve cell-based drug testing. Drug Discov. Today 2002, 7, 612–620. [Google Scholar] [CrossRef]
- Sun, T.; Jackson, S.; Haycock, J.W.; MacNeil, S. Culture of skin cells in 3D rather than 2D improves their ability to survive exposure to cytotoxic agents. J. Biotechnol. 2006, 122, 372–381. [Google Scholar] [CrossRef]
- Knight, E.; Przyborski, S. Advances in 3D cell culture technologies enabling tissue-like structures to be created in vitro. J. Anat. 2015, 227, 746–756. [Google Scholar] [CrossRef] [Green Version]
- Abbott, R.D.; Kaplan, D.L. Strategies for improving the physiological relevance of human engineered tissues. Trends Biotechnol. 2015, 33, 401–407. [Google Scholar] [CrossRef] [Green Version]
- LaPlaca, M.C.; Vernekar, V.N.; Shoemaker, J.T.; Cullen, D.K. Three-dimensional neuronal cultures. In 3D Tissue Engineering; Berthiaume, F., Morgan, J.R., Eds.; Artech House: Boston, MA, USA, 2010; Chapter 11; pp. 187–204. [Google Scholar]
- Bercu, M.M.; Arien-Zakay, H.; Stoler, D.; Lecht, S.; Lelkes, P.I.; Samuel, S.; Or, R.; Nagler, A.; Lazarovici, P.; Elchalal, U. Enhanced survival and neurite network formation of human umbilical cord blood neuronal progenitors in three-dimensional collagen constructs. J. Mol. Neurosci. 2013, 51, 249–261. [Google Scholar] [CrossRef]
- Cukierman, E.; Pankov, R.; Yamada, K.M. Cell interactions with three-dimensional matrices. Curr. Opin. Cell Biol. 2002, 14, 633–639. [Google Scholar] [CrossRef]
- Mazzoleni, G.; Di Lorenzo, D.; Steimberg, N. Modelling tissues in 3D: The next future of pharmacotoxicology and food research. Genes Nutr. 2009, 4, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Vinci, M.; Gowan, S.; Boxall, B.; Patterson, L.; Zimmermann, M.; Court, W.; Lomas, C.; Mendiola, M.; Hardisson, D.; Eccles, S.A. Advances in establishment and analysis of three-dimensional tumor spheroid-based functional assays for target validation and drug evaluation. BMC Biol. 2012, 10, 29. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.; Cheng, K.; Kisaalita, W. Three-dimensional neuronal cell cultures more accurately model Voltage Gated Calcium Channel functionality in freshly dissected nerve tissue. PLoS ONE 2012, 7, 45074. [Google Scholar] [CrossRef]
- Cavallo, F.; Huang, Y.; Dent, E.W.; Williams, J.C.; Lagally, M.G. Neurite guidance and three-dimensional confinement via compliant semiconductor scaffolds. ACS Nano 2014, 8, 12219–12227. [Google Scholar] [CrossRef]
- McNamara, M.C.; Sharifi, F.; Wrede, A.H.; Kimlinger, D.F.; Thomas, D.G.; Vander Wiel, J.B.; Chen, Y.; Montazami, R.; Hashemi, N.N. Microfibers as physiologically relevant platforms for creation of 3D cell cultures. Macromol. Biosci. 2017, 17, 1700279. [Google Scholar] [CrossRef] [Green Version]
- Limongi, T.; Cesca, F.; Gentile, F.; Marotta, R.; Ruffilli, R.; Barberis, A.; Dal Maschio, M.; Petrini, E.M.; Santoriello, S.; Benfenati, F.; et al. Nanostructured superhydrophobic substrates trigger the development of 3D neuronal networks. Small 2013, 9, 402–412. [Google Scholar] [CrossRef]
- Johansson, F.; Carlberg, P.; Danielsen, N.; Montelius, L.; Kanje, M. Axonal outgrowth on nano-imprinted patterns. Biomaterials 2006, 27, 1251–1258. [Google Scholar] [CrossRef]
- Bucaro, M.A.; Vasquez, Y.; Hatton, B.D.; Aizenberg, J. Fine-tuning the degree of stem cell polarization and alignment on ordered arrays of high-aspect-ratio nanopillars. ACS Nano 2012, 6, 6222–6230. [Google Scholar] [CrossRef]
- Dowell-Mesfin, N.M.; Abdul-Karim, M.A.; Turner, A.M.; Schanz, S.; Craighead, H.G.; Roysam, B.; Turner, J.N.; Shain, W. Topographically modified surfaces affect orientation and growth of hippocampal neurons. J. Neural Eng. 2004, 1, 78–90. [Google Scholar] [CrossRef]
- Kwiat, M.; Elnathan, R.; Pevzner, A.; Peretz, A.; Barak, B.; Peretz, H.; Ducobni, T.; Stein, D.; Mittleman, M.; Ashery, U.; et al. Highly ordered large-scale neuronal networks of individual cells—Toward single cell to 3D nanowire intracellular interfaces. ACS Appl. Mater. Interfaces 2012, 4, 3542–3549. [Google Scholar] [CrossRef]
- Yu, M.; Huang, Y.; Ballweg, J.; Shin, H.; Huang, M.; Savage, D.E.; Lagally, M.G.; Dent, E.W.; Blick, R.H.; Williams, J.C. Semiconductor Nanomembrane Tubes: Three-Dimensional Confinement for Controlled Neurite Outgrowth. ACS Nano 2011, 26, 2447–2457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, F.; Huang, Y.; Cangellaris, O.V.; Huang, W.; Dent, E.W.; Gillette, M.U.; Williams, J.C. Toward intelligent synthetic neural circuits: Directing and accelerating neuron cell growth by self-rolled-up silicon nitride microtube array. ACS Nano 2014, 25, 11108–11117. [Google Scholar]
- Mazzini, G.; Carpignano, F.; Surdo, S.; Aredia, F.; Panini, N.; Torchio, M.; Erba, E.; Danova, M.; Scovassi, A.I.; Barillaro, G. 3D Silicon Microstructures: A new tool for evaluating biological aggressiveness of tumor cells. IEEE Trans. Nanobiosci. 2015, 14, 797–805. [Google Scholar] [CrossRef] [PubMed]
- McConnell, S.K. Constructing the cerebral cortex: Neurogenesis and fate determination. Neuron 1995, 15, 761–768. [Google Scholar] [CrossRef] [Green Version]
- Solozobova, V.; Wyvekens, N.; Pruszak, J. Lessons from the embryonic neural stem cell niche for neural lineage differentiation of pluripotent stem cells. Stem Cell. Rev. Rep. 2012, 8, 813–829. [Google Scholar] [CrossRef] [Green Version]
- Boissart, C.; Poulet, A.; Georges, P.; Darville, H.; Julita, E.; Delorme, R.; Bourgeron, T.; Peschanski, M.; Benchoua, A. Differentiation from human pluripotent stem cells of cortical neurons of the superficial layers amenable to psychiatric disease modeling and high-throughput drug screening. Transl. Psychiatry 2013, 3, 294. [Google Scholar] [CrossRef]
- Corti, S.; Faravelli, I.; Cardano, M.; Conti, L. Human pluripotent stem cells as tools for neurodegenerative and neurodevelopmental disease modeling and drug discovery. Expert Opin. Drug Discov. 2015, 10, 615–629. [Google Scholar] [CrossRef]
- Brilli, E.; Reitano, E.; Conti, L.; Conforti, P.; Gulino, R.; Consalez, G.G.; Cesana, E.; Smith, A.; Rossi, F.; Cattaneo, E. Neural stem cells engrafted in the adult brain fuse with endogenous neurons. Stem Cells Dev. 2013, 22, 538–547. [Google Scholar] [CrossRef] [Green Version]
- Conti, L.; Pollard, S.M.; Gorba, T.; Reitano, E.; Toselli, M.; Biella, G.; Sun, Y.; Sanzone, S.; Ying, Q.L.; Cattaneo, E.; et al. Niche-independent symmetrical self-renewal of a mammalian tissue stem cell. PLoS Biol. 2005, 3, 283. [Google Scholar] [CrossRef] [Green Version]
- Pollard, S.; Conti, L.; Smith, A. Exploitation of adherent neural stem cells in basic and applied neurobiology. Regen. Med. 2006, 1, 111–118. [Google Scholar] [CrossRef]
- Pollard, S.M.; Conti, L. Investigating radial glia in vitro. Prog. Neurobiol. 2007, 83, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gjorevski, N.; Ranga, N.; Lutolf, M.P. Bioengineering approaches to guide stem cell-based organogenesis. Development 2014, 141, 1794–1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kratochvil, M.J.; Seymour, A.J.; Li, T.L.; Sergiu, P.P.; Kuo, C.J.; Heilshorn, S.C. Engineered materials for organoid systems. Nat. Rev. Mater. 2019, 4, 606–622. [Google Scholar] [CrossRef]
- Seliktar, D. Designing cell-compatible hydrogels for biomedical applications. Science 2012, 336, 1124–1128. [Google Scholar] [CrossRef]
- Fang, Y.; Eglen, R.M. Three-dimensional cell cultures in drug discovery and development. Slas Discov. 2017, 22, 456–472. [Google Scholar]
- Simitzi, C.; Efstathopoulos, P.; Kourgiantaki, A.; Ranella, A.; Charalampopoulos, I.; Fotakis, C.; Αthanassakis, I.; Stratakis, E.; Gravanis, A. Laser fabricated discontinuous anisotropic microconical substrates as a new model scaffold to control the directionality of neuronal network outgrowth. Biomaterials 2015, 67, 115–128. [Google Scholar] [CrossRef]
- Yao, L.; Wang, S.; Cui, W.; Sherlock, R.; O’Connell, C.; Damodaran, G.; Gorman, A.; Windebank, A.; Pandit, A. Effect of functionalized micropatterned PLGA on guided neurite growth. Acta Biomater. 2009, 5, 580–588. [Google Scholar] [CrossRef]
- Rajnicek, A.; Britland, S.; McCaig, C. Contact guidance of CNS neurites on grooved quartz: Influence of groove dimensions, neuronal age and cell type. J. Cell Sci. 1997, 110, 2905–2913. [Google Scholar]
- Cecchini, M.; Bumma, G.; Serresi, M.; Beltrami, F. PC12 differentiation on biopolymer nanostructures. Nanotechnology 2007, 18, 505103. [Google Scholar] [CrossRef]
- Hanson, J.N.; Motala, M.J.; Heien, M.L.; Gillette, M.; Sweedler, J.; Nuzzo, R.G. Textural guidance cues for controlling process outgrowth of mammalian neurons. Lab Chip 2009, 9, 122–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattotti, M.; Micholt, L.; Braeken, D.; Kovacic, D. Characterization of spiral ganglion neurons cultured on silicon micro-pillar substrates for new auditory neuro-electronic interfaces. J. Neural Eng. 2015, 12, 026001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, M.S.; Hubner, J.; Boisen, A. Large Area Fabrication of Leaning Silicon Nanopillars for Surface Enhanced Raman Spectroscopy. Adv. Mater. 2012, 24, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Magno, G.; Belier, B.; Barbillon, G. Al/Si Nanopillars as Very Sensitive SERS Substrates. Materials 2018, 11, 1534. [Google Scholar] [CrossRef]
- Vereecke, G.; Xu, X.M.; Tsai, W.K.; Yang, H.; Armini, S.; Delande, T.; Doumen, G.; Kentie, F.; Shi, X.; Simms, I.; et al. Partial Wetting of aqueous solutions on high aspect ratio nanopillars with hydrophilic surface finish. ECS J. Solid State Sci. Technol. 2014, 3, 3095–3100. [Google Scholar] [CrossRef]
- Liu, C.; Sun, J.; Li, J.; Xiang, C.; Che, L.; Wang, Z.; Zhou, X. Long-range spontaneous droplet self-propulsion on wettability gradient surfaces. Sci. Rep. 2017, 7, 7552. [Google Scholar] [CrossRef]
- Park, Y.S.; Yoon, S.Y.; Park, J.S.; Lee, J.S. Deflection induced cellular focal adhesion and anisotropic growth on vertically aligned silicon nanowires with differing elasticity. NPG Asia Mater. 2016, 8, 249. [Google Scholar] [CrossRef] [Green Version]
- Tullii, G.; Giona, F.; Lodola, F.; Bonfadini, S.; Bossio, C.; Varo, S.; Desii, A.; Criante, L.; Sala, C.; Pasini, M.; et al. High-aspect-ratio semiconducting polymer pillars for 3D cell cultures. ACS Appl. Mater. Interfaces 2019, 11, 28125–28137. [Google Scholar] [CrossRef] [Green Version]
- Medina Benavente, J.J.; Mogami, H.; Sakurai, T.; Sawada, K. Evaluation of silicon nitride as a substrate for culture of PC12 cells: An interfacial model for functional studies in neurons. PLoS ONE 2014, 9, 90189. [Google Scholar] [CrossRef]
- Liliom, H.; Lajer, P.; Berces, Z.; Csernyus, B.; Szabò, A.; Pinke, D.; Low, P.; Fekete, Z.; Pongracz, A.; Schlett, K. Comparing the effects of uncoated nanostructured surfaces on primary neurons and astrocytes. J. Biomed. Mater. Res. A 2019, 107, 2350–2359. [Google Scholar] [CrossRef]
- Fan, Y.W.; Cui, F.Z.; Hou, S.P.; Xu, Q.Y.; Chen, L.N.; Lee, I.S. Culture of neural cells on silicon wafers with nano-scale surface topograph. J. Neurosci. Methods 2002, 120, 17–23. [Google Scholar] [CrossRef]
- Chen, W.S.; Guo, L.Y.; Tang, C.C.; Tsai, C.K.; Huang, H.H.; Chin, T.Y.; Yang, M.L.; Chen-Yang, Y.W. The effect of laminin surface modification of electrospun silica nanofiber substrate on neuronal tissue engineering. Nanomaterials 2018, 8, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turunen, S.; Joki, T.; Hiltunen, M.L.; Ihalainen, T.O.; Narkilahti, S.; Kellomaki, M. Direct laser writing of tubular microtowers for 3D culture of human pluripotent stem cell-derived neuronal cells. ACS Appl. Mater. Interfaces 2017, 9, 25717–25730. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, F.; Patel, B.B.; Dzuilko, A.K.; Montazami, R.; Sakaguchi, D.S.; Hashemi, N. Polycaprolactone microfibrous scaffols to navigate neural stem cells. Biomacromolecules 2016, 17, 3287–3297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timashev, P.S.; Vedunova, M.V.; Guseva, D.; Ponimaskin, E.; Deiwick, A.; Mishchenko, T.A.; Mitroshina, E.V.; Koroleva, A.V.; Pimashkin, A.S.; Mukhina, I.V.; et al. 3D in vitro platform produced by two-photon polymerization for the analysis of neural network formation and function. Biomed. Phys. Eng. Exp. 2016, 2, 035001. [Google Scholar] [CrossRef] [Green Version]
- Koroleva, A.; Gill, A.A.; Ortega, I.; Haycock, J.W.; Schilie, S.; Gittard, S.D.; Chichkov, B.N.; Claeyssens, F. Two-photon polymerization-generated and micromolding-replicated 3D scaffolds for peripheral neural tissue engineering applications. Biofabrication 2012, 4, 025005. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Slide Code | Pillar Diameter (μm) | Distance Among Pillars (μm) | Topography |
|---|---|---|---|
| A1 | 10 | 20 | Alligned |
| A2 | 10 | 30 | Alligned |
| A3 | 10 | 40 | Alligned |
| A4 | 15 | 20 | Alligned |
| A5 | 15 | 30 | Alligned |
| A6 | 15 | 40 | Alligned |
| S1 | 10 | 20 | Staggered |
| S2 | 10 | 30 | Staggered |
| S3 | 10 | 40 | Staggered |
| S4 | 15 | 20 | Staggered |
| S5 | 15 | 30 | Staggered |
| S6 | 15 | 40 | Staggered |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cutarelli, A.; Ghio, S.; Zasso, J.; Speccher, A.; Scarduelli, G.; Roccuzzo, M.; Crivellari, M.; Maria Pugno, N.; Casarosa, S.; Boscardin, M.; et al. Vertically-Aligned Functionalized Silicon Micropillars for 3D Culture of Human Pluripotent Stem Cell-Derived Cortical Progenitors. Cells 2020, 9, 88. https://doi.org/10.3390/cells9010088
Cutarelli A, Ghio S, Zasso J, Speccher A, Scarduelli G, Roccuzzo M, Crivellari M, Maria Pugno N, Casarosa S, Boscardin M, et al. Vertically-Aligned Functionalized Silicon Micropillars for 3D Culture of Human Pluripotent Stem Cell-Derived Cortical Progenitors. Cells. 2020; 9(1):88. https://doi.org/10.3390/cells9010088
Chicago/Turabian StyleCutarelli, Alessandro, Simone Ghio, Jacopo Zasso, Alessandra Speccher, Giorgina Scarduelli, Michela Roccuzzo, Michele Crivellari, Nicola Maria Pugno, Simona Casarosa, Maurizio Boscardin, and et al. 2020. "Vertically-Aligned Functionalized Silicon Micropillars for 3D Culture of Human Pluripotent Stem Cell-Derived Cortical Progenitors" Cells 9, no. 1: 88. https://doi.org/10.3390/cells9010088
APA StyleCutarelli, A., Ghio, S., Zasso, J., Speccher, A., Scarduelli, G., Roccuzzo, M., Crivellari, M., Maria Pugno, N., Casarosa, S., Boscardin, M., & Conti, L. (2020). Vertically-Aligned Functionalized Silicon Micropillars for 3D Culture of Human Pluripotent Stem Cell-Derived Cortical Progenitors. Cells, 9(1), 88. https://doi.org/10.3390/cells9010088