Previously Identified Genetic Variants in ADGRL3 Are not Associated with Risk for Equine Degenerative Myeloencephalopathy across Breeds



{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
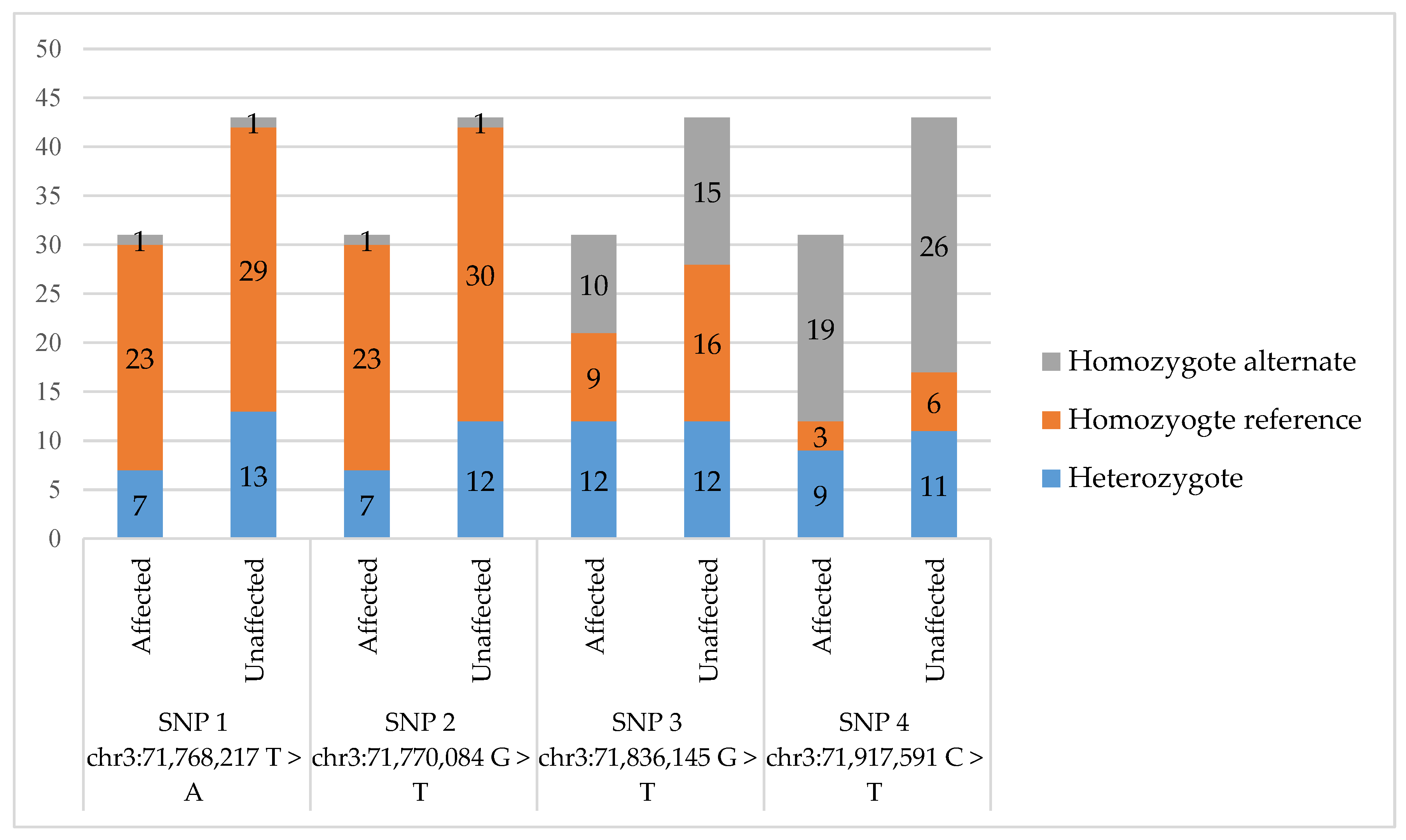
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mayhew, I.M.; Brown, C.M.; Stowe, H.D.; Trapp, A.L.; Derksen, F.J.; Clement, S.F. Equine Degenerative Myeloencephalopathy: A Vitamin E Deficiency that May be Familial. J. Vet. Intern. Med. 1987, 1, 45–50. [Google Scholar] [CrossRef]
- Aleman, M.; Finno, C.J.; Higgins, R.J.; Puschner, B.; Gericota, B.; Gohil, K.; LeCouteur, R.A.; Madigan, J.E. Evaluation of Epidemiological, Clinical, and Pathological Features of Neuroaxonal Dystrophy in Quarter Horses. J. Am. Vet. Med. Assoc. 2011, 239, 823–833. [Google Scholar] [CrossRef]
- Finno, C.J.; Higgins, R.J.; Aleman, M.; Ofri, R.; Hollingsworth, S.R.; Bannasch, D.L.; Reilly, C.M.; Madigan, J.E. Equine Degenerative Myeloencephalopathy in Lusitano Horses. J. Vet. Intern. Med. 2011, 25, 1439–1446. [Google Scholar] [CrossRef]
- Finno, C.J.; Valberg, S.J.; Shivers, J.; D’Almeida, E.; Armién, A.G. Evidence of the Primary Afferent Tracts Undergoing Neurodegeneration in Horses with Equine Degenerative Myeloencephalopathy Based on Calretinin Immunohistochemical Localization. Vet. Pathol. 2016, 53, 77–86. [Google Scholar] [CrossRef]
- Finno, C.J.; Famula, T.; Aleman, M.; Higgins, R.J.; Madigan, J.E.; Bannasch, D.L. Pedigree Analysis and Exclusion of α-Tocopherol Transfer Protein (TTPA) as a Candidate Gene for Neuroaxonal Dystrophy in the American Quarter Horse. J. Vet. Intern. Med. 2013, 27, 177–185. [Google Scholar] [CrossRef]
- Finno, C.J.; Bordbari, M.H.; Valberg, S.J.; Lee, D.; Herron, J.; Hines, K.; Monsour, T.; Scott, E.; Bannasch, D.L.; Mickelson, J.; et al. Transcriptome Profiling of Equine Vitamin E Deficient Neuroaxonal Dystrophy Identifies Upregulation of Liver X Receptor Target Genes. Free Radic. Biol. Med. 2016, 101, 261–271. [Google Scholar] [CrossRef]
- Nappert, G.; Vrins, A.; Breton, L.; Beauregard, M. A Retrospective Study of Nineteen Ataxic Horses. Can. Vet. J. La Rev. Vet. Can. 1989, 30, 802–806. [Google Scholar]
- Posbergh, C.J.; Pollott, G.E.; Southard, T.L.; Divers, T.J.; Brooks, S.A. A Nonsynonymous Change in Adhesion G Protein–Coupled Receptor L3 Associated with Risk for Equine Degenerative Myeloencephalopathy in the Caspian Horse. J. Equine Vet. Sci. 2018, 70, 96–100. [Google Scholar] [CrossRef]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A.M. Primer3Plus, an Enhanced Web Interface to Primer3. Nucleic Acids Res. 2007, 35, W71–W74. [Google Scholar] [CrossRef]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative Genomics Viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef]
- Leeb, T. Concern Regarding the Publication by Posbergh et al. A Nonsynonymous Change in Adhesion G Protein–Coupled Receptor L3 Associated with Risk for Equine Degenerative Myeloencephalopathy in the Caspian Horse. J. Equine Vet. Sci. 2019, 70, 96–100. [Google Scholar] [CrossRef]
- Petersen, J.L.; Mickelson, J.R.; Cothran, E.G.; Andersson, L.S.; Axelsson, J.; Bailey, E.; Bannasch, D.; Binns, M.M.; Borges, A.S.; Brama, P.; et al. Genetic Diversity in the Modern Horse Illustrated from Genome-Wide SNP Data. PLoS ONE 2013, 8, e54997. [Google Scholar] [CrossRef]
- Acosta, M.T.; Swanson, J.; Stehli, A.; Molina, B.S.; MTATeam Martinez, A.F.; Arcos-Burgos, M.; Muenke, M. ADGRL3 (LPHN3) Variants Are Associated with a Refined Phenotype of ADHD in the MTA Study. Mol. Genet. Genom. Med. 2016, 4, 540–547. [Google Scholar] [CrossRef]
- Kappel, D.B.; Schuch, J.B.; Rovaris, D.L.; da Silva, B.S.; Müller, D.; Breda, V.; Teche, S.P.; Riesgo, R.S.; Schüler-Faccini, L.; Rohde, L.A.; et al. ADGRL3 Rs6551665 as a Common Vulnerability Factor Underlying Attention-Deficit/Hyperactivity Disorder and Autism Spectrum Disorder. Neuromol. Med. 2019, 21, 60–67. [Google Scholar] [CrossRef]
- Martinez, A.F.; Abe, Y.; Hong, S.; Molyneux, K.; Yarnell, D.; Löhr, H.; Driever, W.; Acosta, M.T.; Arcos-Burgos, M.; Muenke, M. An Ultraconserved Brain-Specific Enhancer Within ADGRL3 (LPHN3) Underpins Attention-Deficit/Hyperactivity Disorder Susceptibility. Biol. Psychiatry 2016, 80, 943–954. [Google Scholar] [CrossRef] [Green Version]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marquardt, S.A.; Wilcox, C.V.; Burns, E.N.; Peterson, J.A.; Finno, C.J. Previously Identified Genetic Variants in ADGRL3 Are not Associated with Risk for Equine Degenerative Myeloencephalopathy across Breeds. Genes 2019, 10, 681. https://doi.org/10.3390/genes10090681
Marquardt SA, Wilcox CV, Burns EN, Peterson JA, Finno CJ. Previously Identified Genetic Variants in ADGRL3 Are not Associated with Risk for Equine Degenerative Myeloencephalopathy across Breeds. Genes. 2019; 10(9):681. https://doi.org/10.3390/genes10090681
Chicago/Turabian StyleMarquardt, Sabin A., Callie V. Wilcox, Erin N. Burns, Janel A. Peterson, and Carrie J. Finno. 2019. "Previously Identified Genetic Variants in ADGRL3 Are not Associated with Risk for Equine Degenerative Myeloencephalopathy across Breeds" Genes 10, no. 9: 681. https://doi.org/10.3390/genes10090681