Chromosome Mapping of 5S Ribosomal Genes in Indo-Pacific and Atlantic Muraenidae: Comparative Analysis by Dual Colour Fluorescence In Situ Hybridisation

 ,
, 
 and
and
Abstract
1. Introduction
2. Materials and Methods
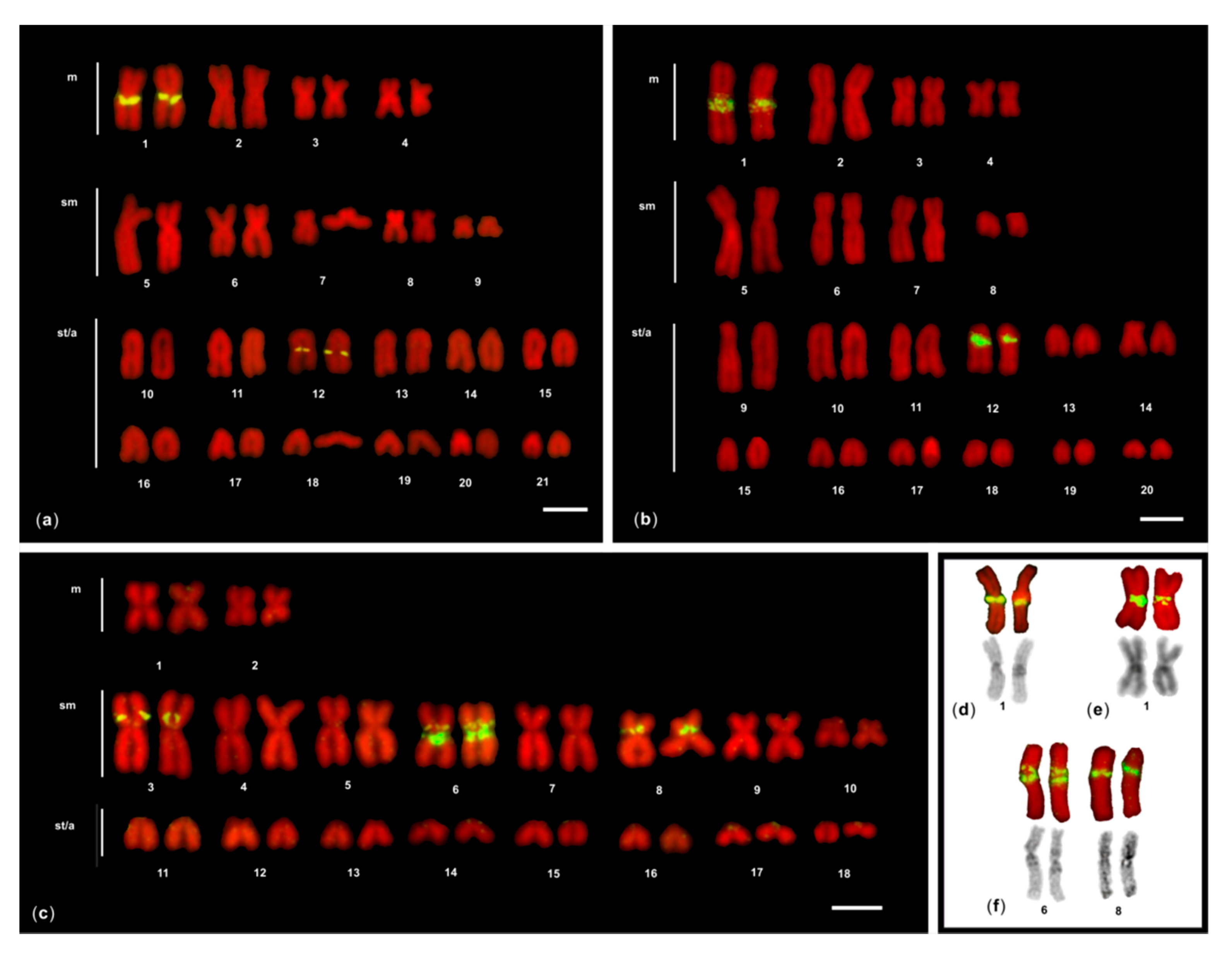
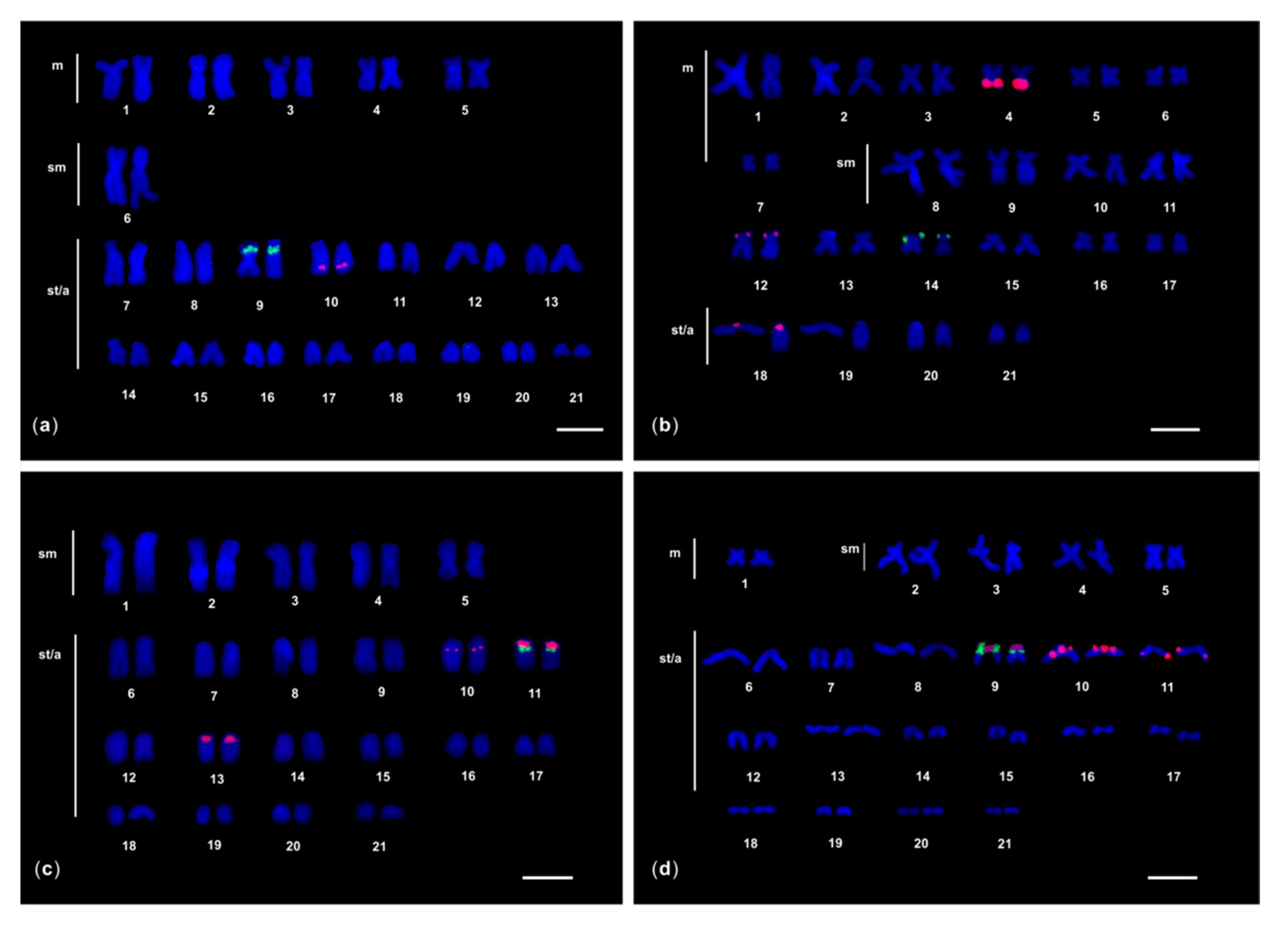
3. Results
3.1. FISH of 5S rDNA
3.1.1. M. helena and G. javanicus
3.1.2. G. flavimarginatus
3.2. Dual Colour FISH of 5S rDNA and 28S rDNA
3.2.1. G. unicolor
3.2.2. G. tile
3.2.3. G. undulatus and G. fimbriatus
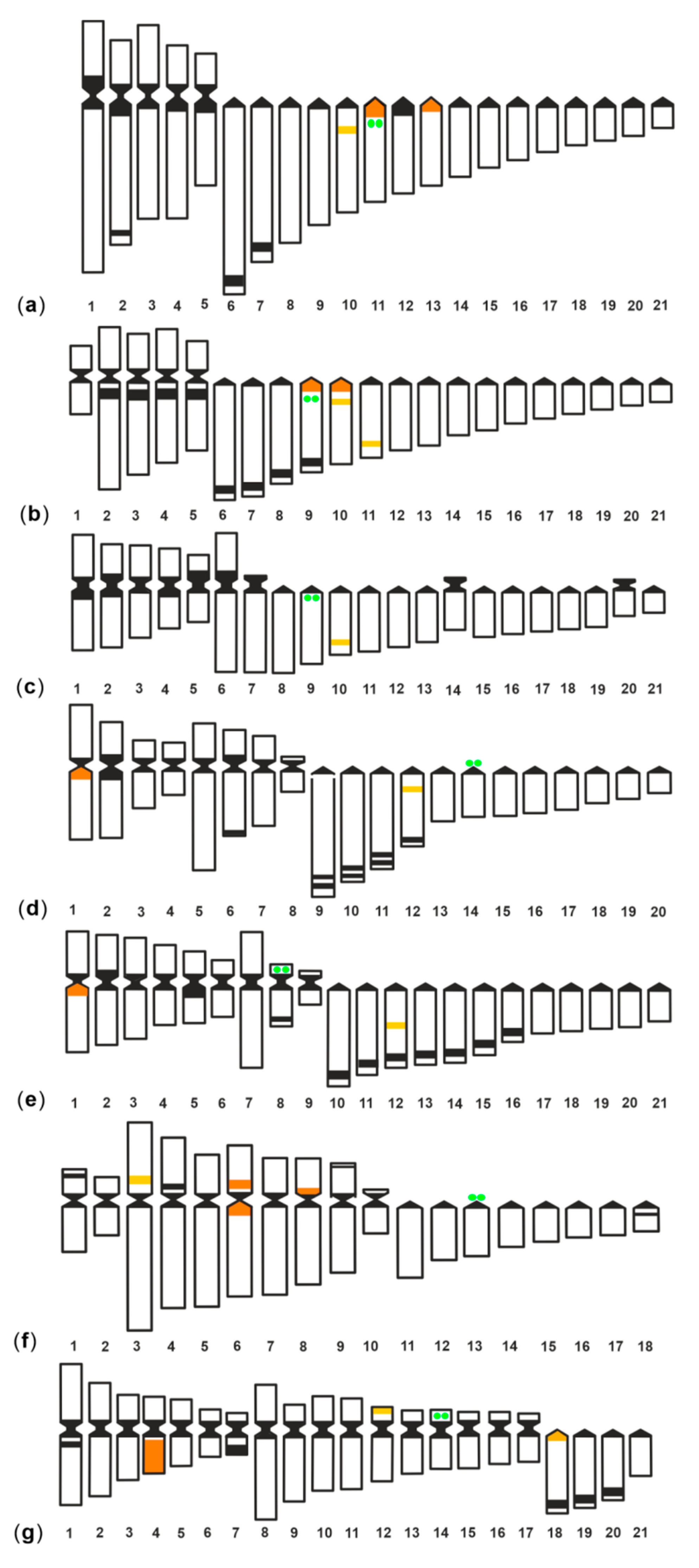
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, J.N.; Samadi, S.; Chen, W.J. Elopomorpha (Teleostei) as a New Model Fish Group for Evolutionary Biology and Comparative Genomics. In Evolutionary Biology: Biodiversification from Genotype to Phenotype; Pontarotti, P., Ed.; Springer: Cham, Switzerland, 2015; pp. 329–344. [Google Scholar] [CrossRef]
- Fricke, R.; Eschmeyer, W.N.; Fong, J.D. Eschmeyer’s Catalog of Fishes: Species by Family/Subfamily. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/SpeciesByFamily.asp Electronic version (accessed on 20 August 2020).
- Wiley, E.O.; Johnson, G.D. A teleost classification based on monophyletic groups. In Origin and Phylogenetic Interrelationships of Teleosts; Nelson, J.S., Schultze, H.-P., Wilson, M.V.H., Eds.; Verlag Dr. Friedrich Pfeil: Munchen, Germany, 2010; pp. 123–182. [Google Scholar]
- Tang, K.L.; Fielitz, C. Phylogeny of moray eels (Anguilliformes: Muraenidae), with a revised classification of true eels (Teleostei: Elopomorpha: Anguilliformes). Mitochondr. DNA 2013, 24, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Santini, F.; Kong, X.; Sorenson, L.; Carnevale, G.; Metha, R.S.; Alfaro, M.E. A multi-locus molecular timescale for the origin and diversification of eels (Order: Anguilliformes). Mol. Phylogenet. Evol. 2013, 69, 884–894. [Google Scholar] [CrossRef] [PubMed]
- Dornburg, A.; Friedman, M.; Near, T.J. Phylogenetic analysis of molecular and morphological data highlights uncertainty in the relationships of fossil and living species of Elopomorpha (Actinopterygii: Teleostei). Mol. Phylogenet. Evol. 2015, 89, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Böhlke, E.B.; Mc Cosker, J.E.; Böhlke, J.E. Family Muraenidae. In Fishes of the Western North Atlantic; Böhlke, E.B., Ed.; Sears Foundation Marine Research: New Haven, CT, USA, 1989; pp. 104–206. [Google Scholar]
- Böhlke, E.B.; Smith, D.G. Type catalogue of Indo-Pacific Muraenidae. P Acad. Nat. Sci. Phila. 2002, 152, 89–172. [Google Scholar] [CrossRef]
- Froese, R.; Pauly, D. (Eds.) FishBase: World Wide Web Electronic Publication. Available online: http://www.fishbase.org (accessed on 20 August 2020).
- Reece, J.S.; Bowen, B.W.; Smith, D.G.; Larson, A. Molecular phylogenetics of moray eels (Muraenidae) demonstrates multiple origins of a shell-crushing jaw (Gymnomuraena, Echidna) and multiple colonizations of the Atlantic Ocean. Mol. Phylogenet. Evol. 2010, 57, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, S.; Schönhuth, S.; Lozano, I.J.; González, J.A.; Sevilla, R.G.; Diez, A.; Bautista, J.M. Morphological, ecological, and molecular analyses separate Muraena augusti from Muraena helena as a valid species. Copeia 2007, 1, 101–113. [Google Scholar] [CrossRef]
- Loh, K.H.; Chen, I.S.; Randall, J.E.; Chen, H.M. A review and molecular phylogeny of the moray eel subfamily Uropterygiinae (Anguilliformes: Muraenidae) from Taiwan, with description of a new species. Raffles Bull. Zool. 2008, 19, 135–150. [Google Scholar]
- Almada, V.C.; Pérez-Portela, R.; Robalo, J.I.; Brito, A. Phylogenetics relationships of Gymnothorax bacalladoi Böhlke and Brito (1987) a poorly known moray eel of the Macronesian islands. Mol. Phylogenet. Evol. 2009, 52, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Yin, S.W.; Niu, B.Z. Phylogenetic relationships of twenty Gymnothorax species based on cytochrome b sequence data. Genet. Mol. Res. 2016, 15, 15028157. [Google Scholar] [CrossRef] [PubMed]
- Ribout, C.; Bech, N.; Briand, M.J.; Guyonnet, D.; Letourneur, Y.; Brischoux, F.; Bonnet, X. A lack of spatial genetic structure of Gymnothorax chilospilus (moray eel) suggests peculiar population functioning. Biol. J. Linn. Soc. 2018, 125, 142–151. [Google Scholar] [CrossRef]
- Nakayama, C.; Jégu, M.; Porto, J.I.R.; Feldberg, E.; McEachran, J.D. Karyological Evidence for a Cryptic Species of Piranha within Serrasalmus rhombeus (Characidae, Serrasalminae) in the Amazon. Copeia 2001, 3, 866–869. [Google Scholar] [CrossRef]
- Souza-Shibatta, L.; Pezenti, L.F.; Ferreira, D.G.; Almeida, F.S.; Sofia, S.H.; Shibatta, O.A. Cryptic species of the genus Pimelodella (Siluriformes: Heptapteridae)from the Miranda River, Paraguay River basin, Pantanal of Mato Grosso do Sul, Central Brazil. Neotrop. Ichthyol. 2013, 11, 101–109. [Google Scholar] [CrossRef]
- Molina, W.F.; Martinez, P.A.; Bertollo, L.C.; Bidau, C.J. Evidence for meiotic drive as an explanation for karyotype changes in fishes. Mar. Genom. 2014, 15, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Sola, L.; Camerini, B.; Cataudella, S. Cytogenetics of Atlantic eels: C- and G-banding, nucleolus organizer regions, and DNA content. Cytogenet. Cell Genet. 1984, 38, 206–210. [Google Scholar] [CrossRef]
- Salvadori, S.; Deiana, A.M.; Deidda, F.; Lobina, C.; Mulas, A.; Coluccia, E. XX/XY sex chromosome system and chromosome markers in the snake eel Ophisurus serpens (Anguilliformes: Ophichtidae). Mar. Biol. Res. 2018, 14, 158–164. [Google Scholar] [CrossRef]
- Takai, A.; Ojima, Y. Karyotypic studies of five species of Anguilliformes (Pisces). Proc. Japan Acad. Ser B. 1985, 61, 253–256. [Google Scholar] [CrossRef][Green Version]
- Nirchio, M.; Rossi, A.R.; Foresti, F.; Oliveira, C. Chromosome evolution in fishes: A new challenging proposal from Neotropical species. Neotrop. Ichthyol. 2014, 12, 761–770. [Google Scholar] [CrossRef]
- Coluccia, E.; Deidda, F.; Cannas, R.; Lobina, C.; Cuccu, D.; Deiana, A.M.; Salvadori, S. Comparative cytogenetics of six Indo-Pacific moray eels (Anguilliformes: Muraenidae) by chromosomal banding and fluorescence in situ hybridization. J. Fish Biol. 2015, 87, 634–645. [Google Scholar] [CrossRef]
- Salvadori, S.; Coluccia, E.; Cannas, R.; Cau, A.; Deiana, A.M. Replication banding in two Mediterranean moray eels: Chromosomal characterization and comparison. Genetica 2003, 119, 253–258. [Google Scholar] [CrossRef]
- Coluccia, E.; Deiana, A.M.; Libertini, A.; Salvadori, S. Cytogenetic characterization of the moray eel Gymnothorax tile and chromosomal banding comparison in Muraenidae (Anguilliformes). Mar. Biol. Res. 2010, 6, 106–111. [Google Scholar] [CrossRef]
- Pichiri, G.; Nieddu, M.; Mezzanotte, R.; Coni, P.P.; Salvadori, S.; Deiana, M.; Deiana, A.M.; Mezzanotte, R. The molecular characterization of the genome of Muraena helena L. I. Isolation and hybridization of two MboI-restricted DNA fractions. Genome 1995, 38, 809–813. [Google Scholar] [CrossRef]
- Salvadori, S.; Deiana, A.M.; Coluccia, E.; Cannas, R.; Cau, A.; Milia, A. Heterochromatin distribution and structure in Gymnothorax unicolor (Anguilliformes, Muraenidae). Ital. J. Zool. 1997, 64, 125–129. [Google Scholar] [CrossRef][Green Version]
- Coluccia, E.; Pichiri, G.; Nieddu, M.; Coni, P.; Manconi, S.; Deiana, A.M.; Salvadori, S.; Mezzanotte, R. Identification of two new repetitive elements and chromosomal mapping of repetitive DNA sequences in the fish Gymnothorax unicolor (Anguilliformes: Muraenidae). Eur. J. Histochem. 2011, 55, 68–72. [Google Scholar] [CrossRef]
- Pichiri, G.; Coni, P.P.; Deiana, A.M.; Nieddu, M.; Mezzanotte, R. On the variability of MboI repeated sequences and 5S rDNA in Muraena helena and Gymnothorax unicolor (Anguilliformes, Muraenidae). Chromosome Res. 2000, 8, 443–445. [Google Scholar] [CrossRef]
- Pendás, A.M.; Morán, P.; Freije, J.P.; García-Vázquez, E. Chromosomal mapping and nucleotide sequence of two tandem repeats of Atlantic salmon 5S rDNA. Cytogenet. Genome Res. 1994, 67, 31–36. [Google Scholar] [CrossRef]
- Zardoya, R.; Meyer, A. Evolutionary relationships of the coelacanth, lungfishes, and tetrapods based on the 28S ribosomal RNA gene. PNAS 1996, 93, 5449–5454. [Google Scholar] [CrossRef] [PubMed]
- Cau, A.; Salvadori, S.; Deiana, A.M.; Bella, J.L.; Mezzanotte, R. The characterization of Muraena helena L. mitotic chromosomes: Karyotype, C-banding, nucleolar organizer regions, and in situ digestion with restriction endonucleases. Cytogenet. Cell Genet. 1988, 47, 223–226. [Google Scholar] [CrossRef]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Deiana, A.M.; Salvadori, S.; Cau, A. The characterization of somatic chromosomes of Gymnothorax unicolor (Delaroche, 1809) by C-banding and NOR staining (Osteichthyes, Anguilliformes). Genetica 1990, 81, 17–20. [Google Scholar] [CrossRef]
- Martínez, J.L.; Morán, P.; García-Vázquez, E.; Pendás, A.M. Chromosomal localization of the major and 5S rRNA genes in the European eel (Anguilla anguilla). Cytogenet. Cell Genet. 1996, 73, 149–152. [Google Scholar] [CrossRef]
- Nieddu, M.; Pichiri, G.; Coni, P.; Salvadori, S.; Deiana, A.M.; Mezzanotte, R. A comparative analysis of European and American eel (Anguilla anguilla and Anguilla rostrata) genomic DNA: 5S rDNA polymorphism permits the distinction between the two populations. Genome 1998, 41, 728–732. [Google Scholar] [CrossRef]
- Deiana, A.M.; Coluccia, E.; Cannas, R.; Pesci, P.; Fonnesu, A.; Salvadori, S. Colocalization of the ribosomal gene families in Conger conger (Anguilliformes, Congridae). Ital. J. Zool. 2006, 73, 1–5. [Google Scholar] [CrossRef]
- Long, E.O.; Dawid, I.D. Repeated genes in eukaryotes. Annu. Rev. Biochem. 1980, 49, 727–764. [Google Scholar] [CrossRef]
- Cabral-de-Mello, D.C.; Cabrero, J.; López-León, M.D.; Camacho, J.P.M. Evolutionary dynamics of 5S rDNA location in acridid grasshoppers and its relationship with H3 histone gene and 45S rDNA location. Genetica 2011, 139, 921–931. [Google Scholar] [CrossRef]
- Pinhal, D.; Araki, C.S.; Gadig, O.B.F.; Martins, C. Molecular organization of 5S rDNA in sharks of the genus Rhizoprionodon: Insights into the evolutionary dynamics of 5S rDNA in vertebrate genomes. Genet. Res. Camb. 2009, 91, 61–72. [Google Scholar] [CrossRef]
- Ubeda-Manzanaro, M.; Merlo, M.A.; Palazon, J.L.; Sarasquete, C.; Rebordinos, L. Sequence characterization and phylogenetic analysis of the 5S ribosomal DNA in species of the family Batrachoididae. Genome 2010, 53, 723–730. [Google Scholar] [CrossRef]
- Ferreira, I.A.; Oliveira, C.; Venere, P.C.; Galetti, P.M., Jr.; Martins, C. 5S rDNA variation and its phylogenetic inference in the genus Leporinus (Characiformes: Anostomidae). Genetica 2007, 129, 253–257. [Google Scholar] [CrossRef]
- Campo, D.; Machado-Schiaffino, G.; Horreo, J.L.; Garcia-Vazquez, E. Molecular organization and evolution of 5S rDNA in the genus Merluccius and their phylogenetic implications. J. Mol. Evol. 2009, 68, 208–216. [Google Scholar] [CrossRef]
- Merlo, M.A.; Cross, I.; Manchado, M.; Cardenas, S.; Rebordinos, L. The 5S rDNA high dynamism in Diplodus sargus is a transposon-mediated mechanism. Comparison with other multi gene families and Sparidae species. J. Mol. Evol. 2013, 76, 83–97. [Google Scholar] [CrossRef]
- Martins, C.; Wasko, A.P. Organization and evolution of 5S Ribosomal DNA in the fish genome. In Focus on Genome Research; Williams, C.R., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2004; pp. 335–363. [Google Scholar]
- Cioffi, M.B.; Bertollo, L.A.C. Chromosomal distribution and evolution of repetitive DNAs in fish. In Repetitive DNA; Garrido-Ramos, M.A., Ed.; Karger: Basel, Switzerland, 2012; pp. 197–221. [Google Scholar] [CrossRef]
- Vasconcelos, A.J.M.; Molina, W.F. Cytogenetical studies in five Atlantic Anguilliformes fishes. Genet. Mol. Biol. 2009, 32, 83–90. [Google Scholar] [CrossRef]
- Rebordinos, L.; Cross, I.; Merlo, A. High Evolutionary Dynamism in 5S rDNA of Fish: State of the Art. Cytogenet. Genome Res. 2013, 141, 103–113. [Google Scholar] [CrossRef]
- Smith, D.G.; Bogorodsky, S.V.; Mal, A.O.; Alpermann, T.J. Review of the moray eels (Anguilliformes: Muraenidae) of the Red Sea, with description of a new species. Zootaxa 2019, 4704, 1–87. [Google Scholar] [CrossRef]
- Gromicho, M.; Coutanceau, J.-P.; Ozouf-Costaz, C.; Collares-Pereira, M.J. Contrast between extensive variation of 28S rDNA and stability of 5S rDNA and telomeric repeats in the diploid-polyploid Squalius alburnoides complex and in its maternal ancestor Squalius pyrenaicus (Teleostei, Cyprinidae). Chrom. Res. 2006, 14, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, M.; Abel, L.D.; Moreira-filho, O. Conserved 5S and variable 45S rDNA chromosomal localization revealed by FISH in Astyanax scabripinnis (Pisces, Characidae). Genetica 2005, 123, 211–216. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | 2n | NF | 45S rDNA Location/Chrom | 5S rDNA | References | ||
|---|---|---|---|---|---|---|---|
| Sites (N) | Pairs (N) | Location/Chrom | |||||
| Family Muraenidae | |||||||
| Gymnothorax undulatus | 42 | 52 | i/a11 | 3 | 3 | i/a10; p/a11; p/a13 | [23], present study |
| G. fimbriatus | 42 | 52 | i/a9 | 4 | 3 | p/a9; p/a10; i/a10; i/a11 | [23], present study |
| G. unicolor | 42 | 54 | i/a9 | 1 | 1 | i/a10 | [28], present study |
| G. javanicus | 40 | 56 | d/a14 | 2 | 2 | p/qarm1; i/a12 | [23], present study |
| Muraena helena | 42 | 60 | d/parm8 | 2 | 2 | p/qarm1; i/a12 | [29], present study |
| G. flavimarginatus | 36 | 56 | d/a13 | 4 | 3 | i/parm3; p/parm6; i/parm6; p/parm8 | [23], present study |
| G. tile | 42 | 76 | d/parm14 | 3 | 3 | qarm4; d/parm12; p/A18 | [25], present study |
| Family Anguillidae | |||||||
| Anguilla anguilla | 38 | 60 | d/parm8 | 1 | 1 | p/a19 | [19,35] |
| A. rostrata | 38 | 60 | d/parm8 | 1 | 1 | p/a19 | [19,36] |
| Family Congridae | |||||||
| Conger conger | 38 | 50 | p/a19 | 1 | 1 | p/a19 | [37] |
| Family Ophichthidae | |||||||
| Ophisurus serpens | 38 | ♀ 76 ♂ 75 | d/parm9 | ♀ 4 ♂ 3 | ♀ 2 ♂ 1 + X | ♀d/parm15; d/parmX ♂ d/parm15; d/parmX | [20] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coluccia, E.; Deidda, F.; Lobina, C.; Melis, R.; Porcu, C.; Agus, B.; Salvadori, S. Chromosome Mapping of 5S Ribosomal Genes in Indo-Pacific and Atlantic Muraenidae: Comparative Analysis by Dual Colour Fluorescence In Situ Hybridisation. Genes 2020, 11, 1319. https://doi.org/10.3390/genes11111319
Coluccia E, Deidda F, Lobina C, Melis R, Porcu C, Agus B, Salvadori S. Chromosome Mapping of 5S Ribosomal Genes in Indo-Pacific and Atlantic Muraenidae: Comparative Analysis by Dual Colour Fluorescence In Situ Hybridisation. Genes. 2020; 11(11):1319. https://doi.org/10.3390/genes11111319
Chicago/Turabian StyleColuccia, Elisabetta, Federica Deidda, Cinzia Lobina, Riccardo Melis, Cristina Porcu, Blondine Agus, and Susanna Salvadori. 2020. "Chromosome Mapping of 5S Ribosomal Genes in Indo-Pacific and Atlantic Muraenidae: Comparative Analysis by Dual Colour Fluorescence In Situ Hybridisation" Genes 11, no. 11: 1319. https://doi.org/10.3390/genes11111319
APA StyleColuccia, E., Deidda, F., Lobina, C., Melis, R., Porcu, C., Agus, B., & Salvadori, S. (2020). Chromosome Mapping of 5S Ribosomal Genes in Indo-Pacific and Atlantic Muraenidae: Comparative Analysis by Dual Colour Fluorescence In Situ Hybridisation. Genes, 11(11), 1319. https://doi.org/10.3390/genes11111319