Development of Molecular Marker Linked with Bacterial Fruit Blotch Resistance in Melon (Cucumis melo L.)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Population Development
2.2. Bacterial Culture and Inoculum Preparation
2.3. Inoculation, Sample Preparation and Assessment of BFB Resistance
2.4. Selection of Putative R-Genes and Primer Design
2.5. Extraction of Genomic DNA and Detection of DNA Polymorphism
2.6. Cloning and Sequencing of the Polymorphic Gene
2.7. Total RNA Extraction and cDNA Synthesis
2.8. Quantitative RT-PCR (qRT-PCR) Expression of Polymorphic Gene MELO3C022157
2.9. Statistical Analysis
3. Results
3.1. Dominant Inheritance of BFB Resistance in Melon
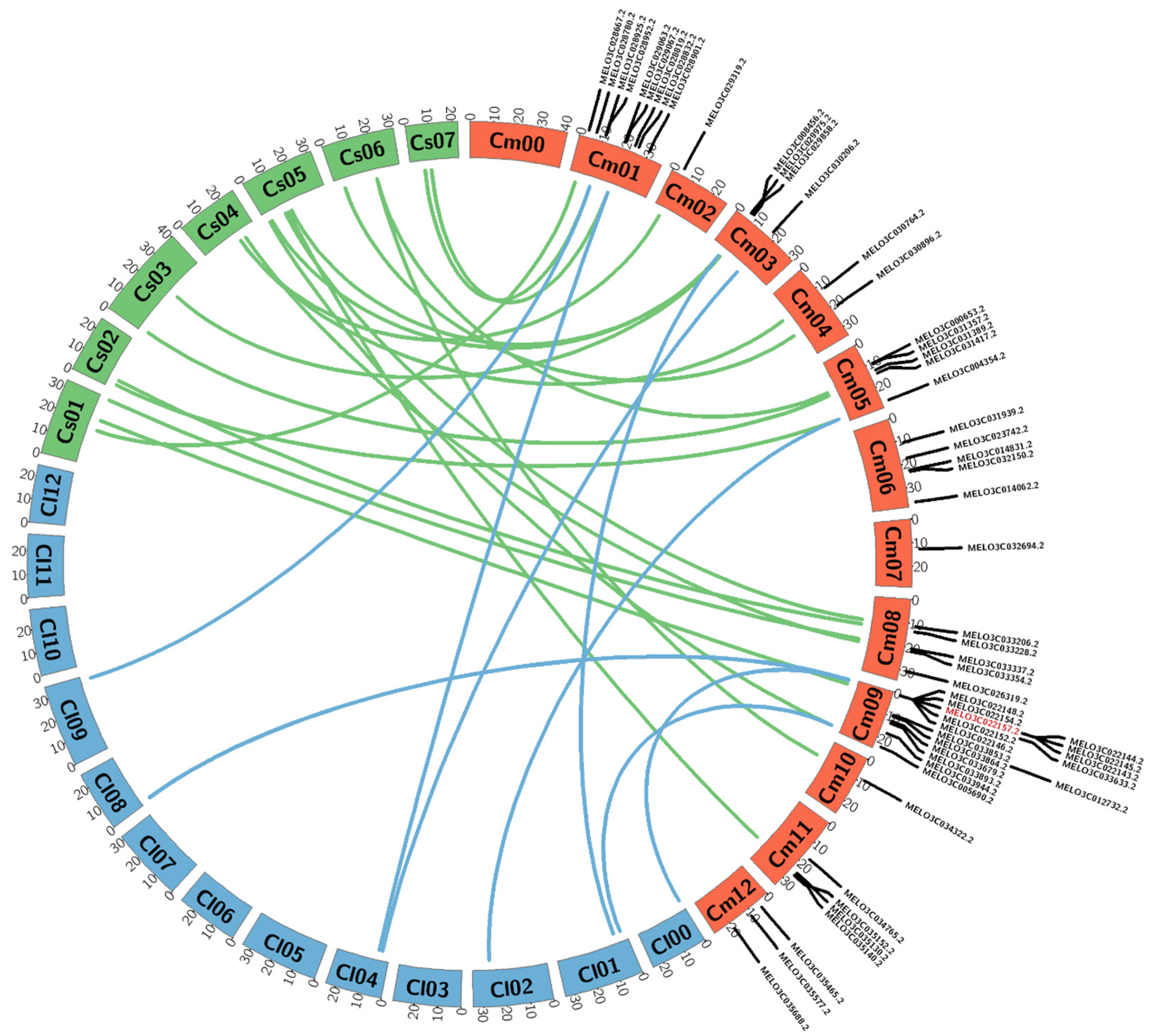
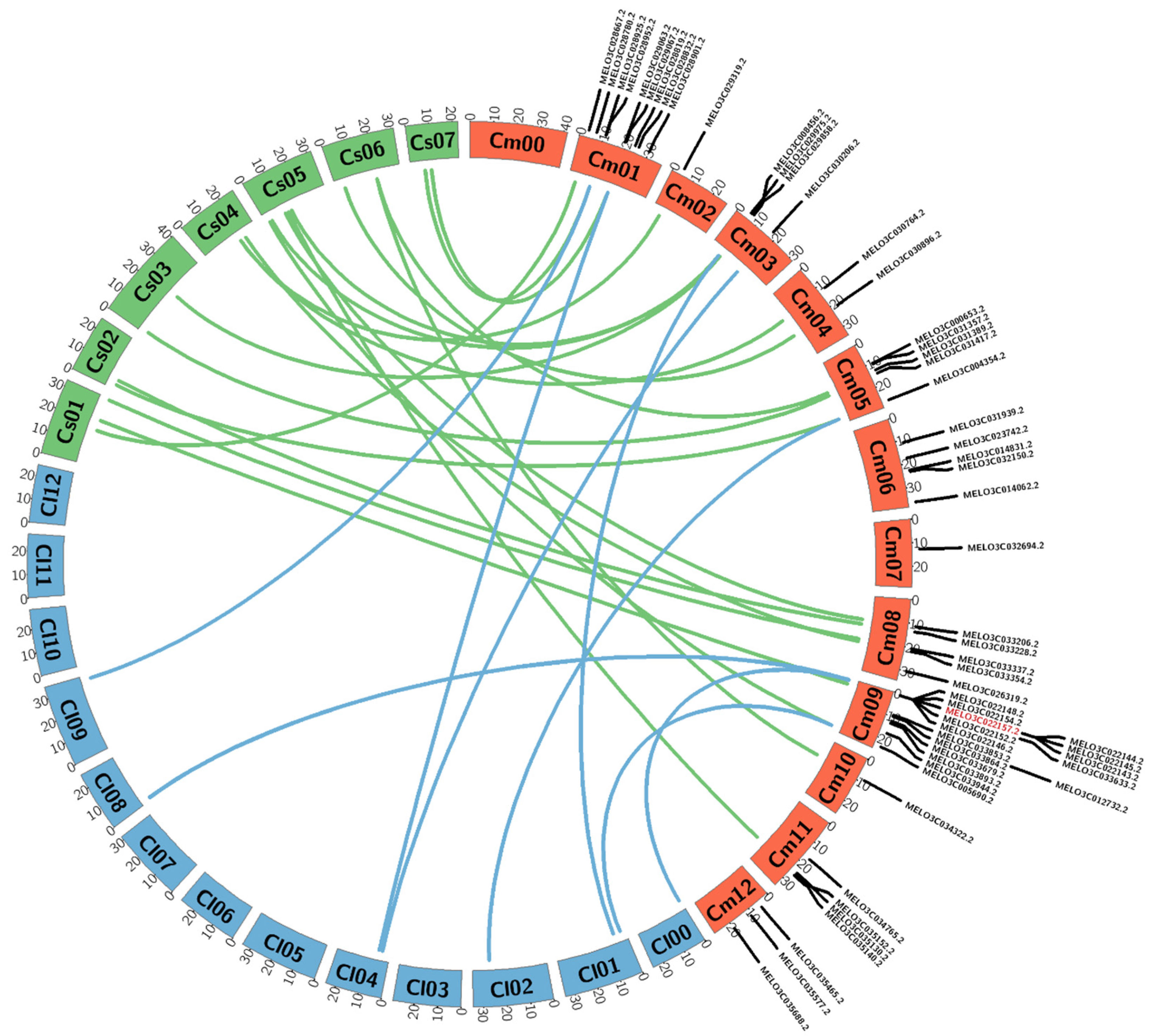
3.2. Microsynteny Analysis of 57 R-Genes of Melon Compared with Watermelon and Cucumber
3.3. Identification of Length Polymorphisms in Putative R-genes
3.4. Cloning and Sequencing of the Polymorphic Gene
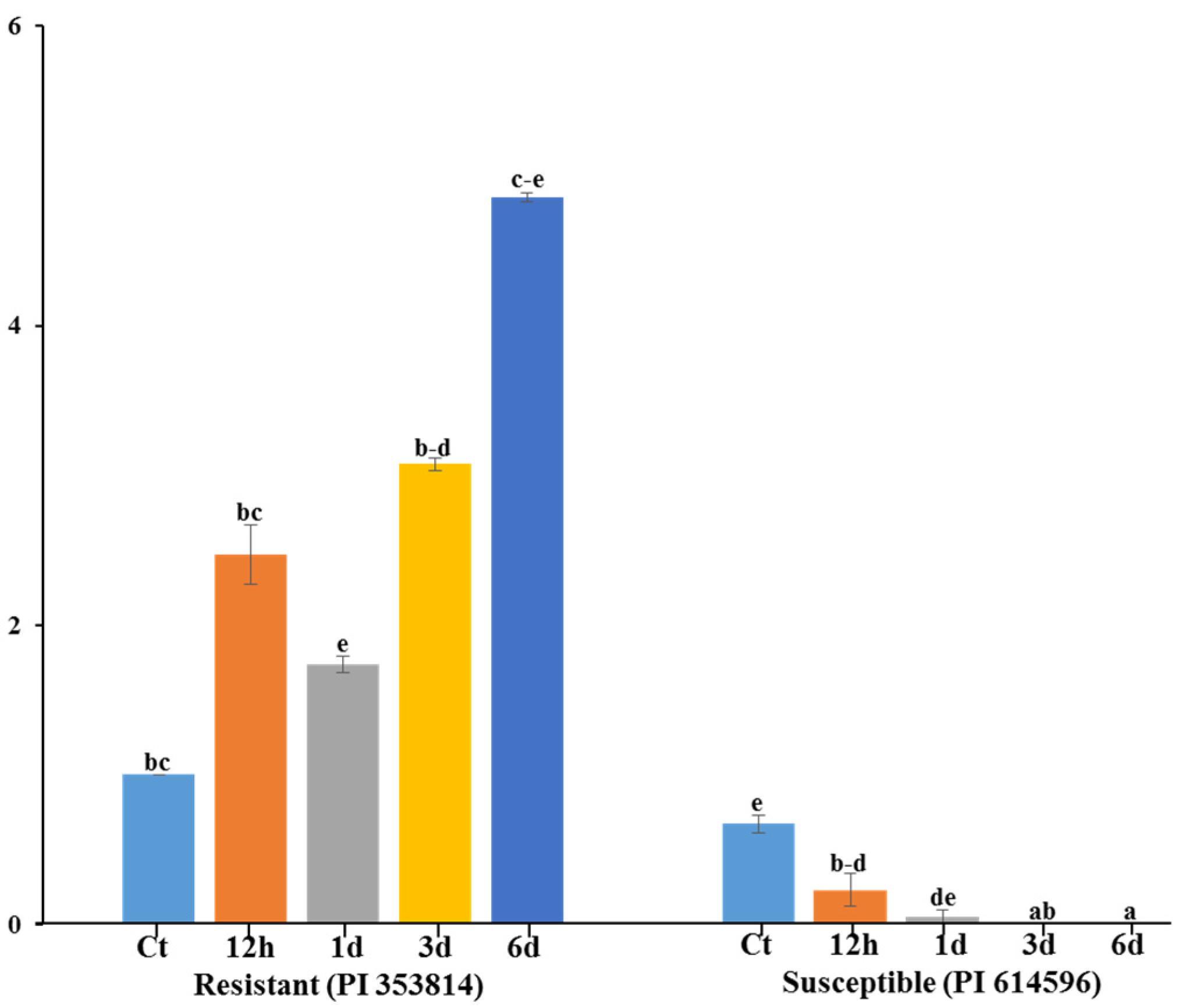
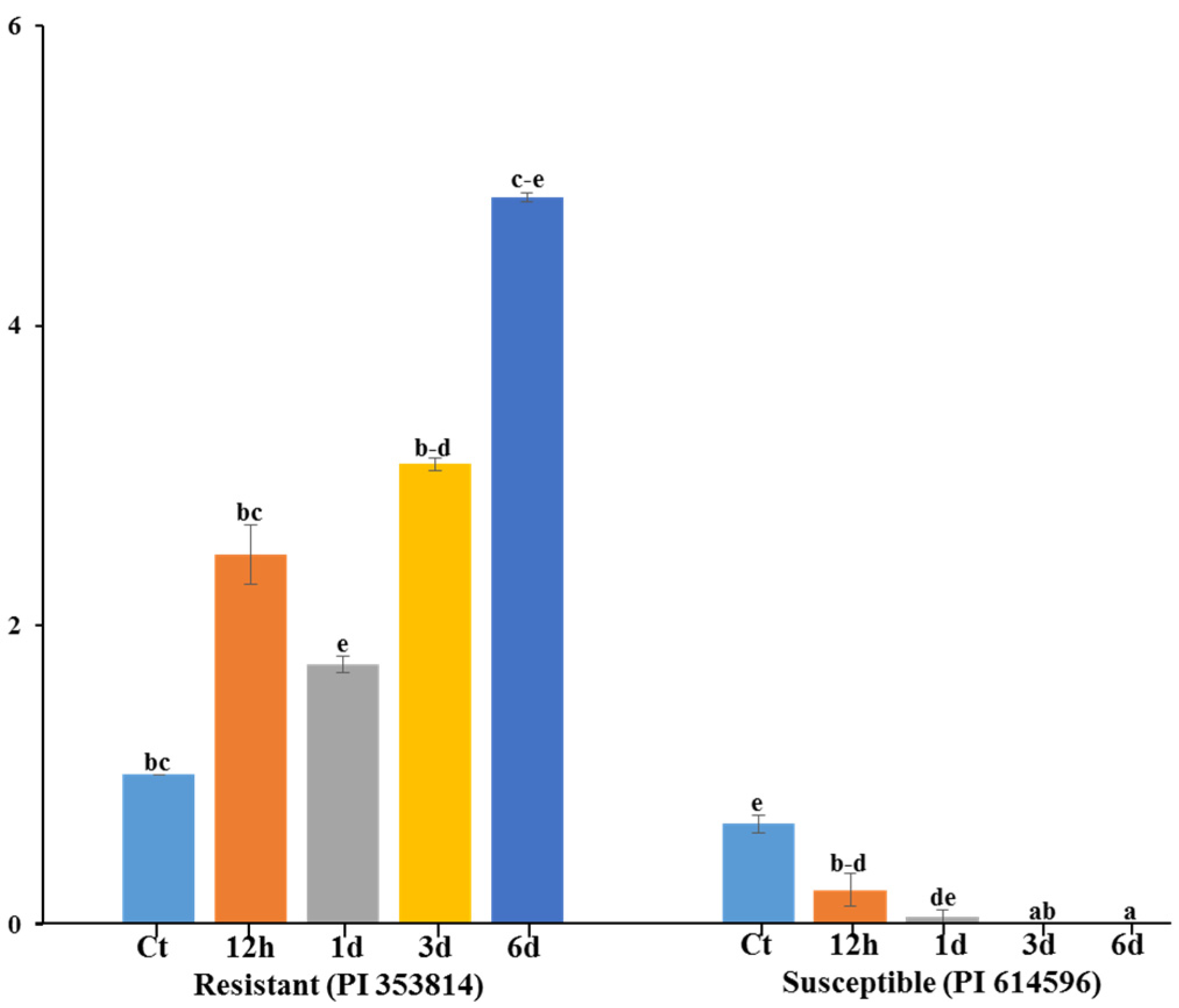
3.5. qRT-PCR Expression Analysis of Polymorphic Gene MELO3C022157
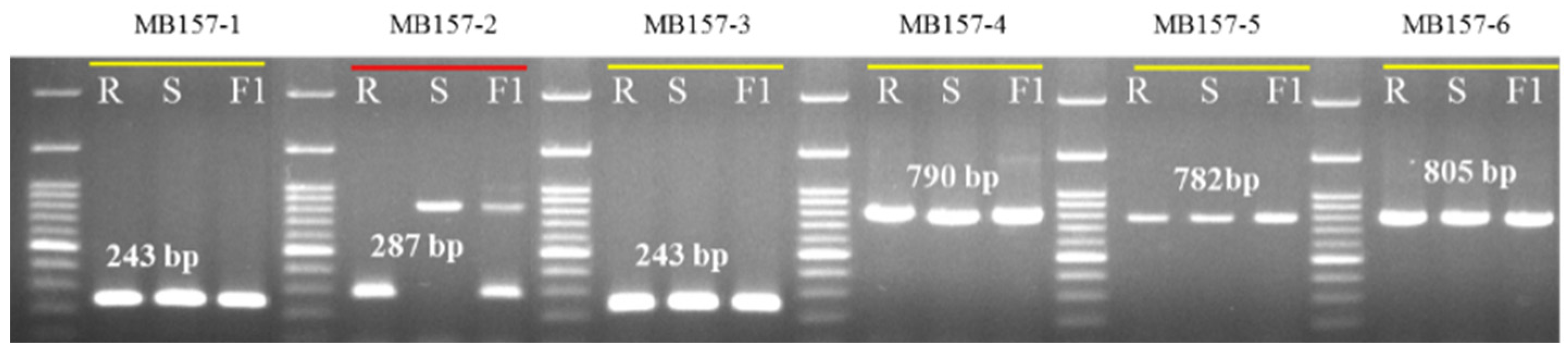
3.6. Development and Validation of InDel Marker Linked to BFB Resistance in Melon
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lester, G.E. Antioxidant, Sugar, Mineral, and Phytonutrient Concentrations across Edible Fruit Tissues of Orange-Fleshed Honeydew Melon (Cucumis melo L.). J. Agric. Food Chem. 2008, 56, 3694–3698. [Google Scholar] [CrossRef] [PubMed]
- FAOstat. Agriculture Organization of the United Nations Statistics Division. Economic and Social Development Department, Rome, Italy. Available online: http://faostat3.fao.org/home/E (accessed on 24 December 2019).
- Schaad, N.W.; Postnikova, E.; Sechler, A.; Claflin, L.E.; Vidaver, A.K.; Jones, J.B.; Agarkova, I.; Ignatov, A.; Dickstein, E.; Ramundo, B.A. Reclassification of subspecies of Acidovorax avenae as A. Avenae (Manns 1905) emend., A. cattleyae (Pavarino, 1911) comb. nov., A. citrulli Schaad et al., 1978) comb. nov., and proposal of A. oryzae sp. nov. Syst. Appl. Microbiol. 2008, 31, 434–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sales Júnior, R.; Menezes, J. Mapeamento das doenças fúngicas, bacterianas e viróticas do cultivo do melão no Estado do RN. Mossoró. Escola Superior de Agricultura de Mossoró 2001. [Google Scholar]
- Conceição, C.S.; Felix, K.C.S.; Mariano, R.L.; Medeiros, E.V.; Souza, E.B. Combined effect of yeast and silicon on the control of bacterial fruit blotch in melon. Sci. Hortic. 2014, 174, 164–170. [Google Scholar] [CrossRef]
- De Melo, E.A.; Rosa de Lima, R.M.; Laranjeira, D.; dos Santos, L.A.; de Omena Gusmão, L.; de Souza, E.B. Efficacy of yeast in the biocontrol of bacterial fruit blotch in melon plants. Trop. Plant Pathol. 2015, 40, 56–64. [Google Scholar] [CrossRef]
- Isakeit, T.; Black, M.; Barnes, L.; Jones, J. First report of infection of honeydew with Acidovorax avenae subsp. citrulli. Plant Dis. 1997, 81, 694. [Google Scholar] [CrossRef]
- Isakeit, T.; Black, M.; Jones, J. Natural infection of citronmelon with Acidovorax avenae subsp. citrulli. Plant Dis. 1998, 82, 351. [Google Scholar] [CrossRef]
- Langston, D., Jr.; Walcott, R.; Gitaitis, R.; Sanders, F., Jr. First report of a fruit rot of pumpkin caused by Acidivorax avenae subsp. citrulli in Georgia. Plant Dis. 1999, 83, 199. [Google Scholar] [CrossRef]
- O’brien, R.; Martin, H. Bacterial blotch of melons caused by strains of Acidovorax avenae subsp. citrulli. Aust. J. Exp. Agric. 1999, 39, 479–485. [Google Scholar]
- Schaad, N.; Postnikova, E.; Randhawa, P. Emergence of Acidovorax avenae Subsp. citrulli as a Crop Threatening Disease of Watermelon and Melon. In Pseudomonas Syringae and Related Pathogens; Springer: Berlin/Heidelberg, Germany, 2003; pp. 573–581. [Google Scholar]
- Walcott, R.; Fessehaie, A.; Castro, A. Differences in pathogenicity between two genetically distinct groups of Acidovorax avenae subsp. citrulli on cucurbit hosts. J. Phytopathol. 2004, 152, 277–285. [Google Scholar] [CrossRef]
- Martin, G.B. Functional analysis of plant disease resistance genes and their downstream effectors. Curr. Opin. Plant. Biol. 1999, 2, 273–279. [Google Scholar] [CrossRef]
- Noh, J.; Kim, J.-H.; Lim, J.H.; Kim, T.B.; Seong, M.H.; Jung, G.T.; Kim, J.M.; Cheong, S.-S.; Oh, N.K.; Lee, W.-H. Occurrence of diseases and case of clinical diagnosis on watermelon in South Korea, 2008–2012. Res. Plant Dis. 2014, 20, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Schaad, N.W.; Sowell, G., Jr.; Goth, R.; Colwell, R.; Webb, R. Pseudomonas pseudoalcaligenes subsp. citrulli subsp.nov. Int. J. Syst. Evol. Microbiol. 1978, 28, 117–125. [Google Scholar]
- Somodi, G.C.; Jones, J.; Hopkins, D.; Stall, R.; Kucharek, T.; Hodge, N.; Watterson, J. Occurrence of a bacterial watermelon fruit blotch in Florida. Plant Dis. 1991, 75, 1053–1056. [Google Scholar] [CrossRef]
- Walcott, R.; Gitaitis, R. Detection of Acidovorax avenae subsp. citrulli in watermelon seed using immunomagnetic separation and the polymerase chain reaction. Plant Dis. 2000, 84, 470–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.R.; Hossain, M.R.; Kim, H.-T.; Nath, U.K.; Abuyusuf, M.; Jung, H.-J.; Park, J.-I.; Nou, I.-S. Molecular characterization of Acidovorax citrulli strain NIHHS15-280 causing bacterial fruit blotch disease in Korea and screening of resistance sources in melon. Hortic. Environ. Biotechnol. 2019, 62, 115–126. [Google Scholar] [CrossRef]
- Carvalho, F.C.; Santos, L.A.; Dias, R.C.; Mariano, R.L.; Souza, E.B. Selection of watermelon genotypes for resistance to bacterial fruit blotch. Euphytica 2013, 190, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Burdman, S.; Walcott, R. Acidovorax citrulli: Generating basic and applied knowledge to tackle a global threat to the cucurbit industry. Mol. Plant Pathol. 2012, 13, 805–815. [Google Scholar] [CrossRef]
- Hopkins, D. The Hypothetical Exam Question Becomes Reality. Plant Dis. 1995, 79, 761–765. [Google Scholar]
- Hopkins, D.L.; Thompson, C.M.; Hilgren, J.; Lovic, B. Wet seed treatment with peroxyacetic acid for the control of bacterial fruit blotch and other seedborne diseases of watermelon. Plant Dis. 2003, 87, 1495–1499. [Google Scholar] [CrossRef] [Green Version]
- Stephens, D.; Schneider, R.; Walcott, R.; Johnson, C. A Procedure, Based on Exposure to Chlorine Gas, for Disinfesting Watermelon Seeds. Phytopathology 2018, S150–S151. [Google Scholar]
- Feng, J.; Li, J.; Randhawa, P.; Bonde, M.; Schaad, N.W. Evaluation of seed treatments for the eradication of Acidovorax avenae subsp. citrulli from melon and watermelon seeds. Can J. Plant Pathol 2009, 31, 180–185. [Google Scholar] [CrossRef]
- Wechter, W.P.; Levi, A.; Ling, K.-S.; Kousik, C.; Block, C.C. Identification of resistance to Acidovorax avenae subsp. citrulli among melon (Cucumis spp.) plant introductions. HortScience 2011, 46, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Bahar, O.; Kritzman, G.; Burdman, S. Bacterial fruit blotch of melon: Screens for disease tolerance and role of seed transmission in pathogenicity. Eur. J. Plant Pathol. 2009, 123, 71–83. [Google Scholar] [CrossRef]
- Ellis, J.; Dodds, P.; Pryor, T. Structure, function and evolution of plant disease resistance genes. Curr. Opin. Plant Biol. 2000, 3, 278–284. [Google Scholar] [CrossRef]
- Harris, C.J.; Slootweg, E.J.; Goverse, A.; Baulcombe, D.C. Stepwise artificial evolution of a plant disease resistance gene. Proc. Natl. Acad. Sci. USA 2013, 110, 21189–21194. [Google Scholar] [CrossRef] [Green Version]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef]
- Meyers, B.C.; Kozik, A.; Griego, A.; Kuang, H.; Michelmore, R.W. Genome-wide analysis of NBS-LRR–encoding genes in Arabidopsis. Plant Cell 2003, 15, 809–834. [Google Scholar] [CrossRef] [Green Version]
- Monosi, B.; Wisser, R.; Pennill, L.; Hulbert, S. Full-genome analysis of resistance gene homologues in rice. Theor. Appl. Genet 2004, 109, 1434–1447. [Google Scholar] [CrossRef]
- Wan, H.; Yuan, W.; Bo, K.; Shen, J.; Pang, X.; Chen, J. Genome-wide analysis of NBS-encoding disease resistance genes in Cucumis sativus and phylogenetic study of NBS-encoding genes in Cucurbitaceae crops. BMC Genom. 2013, 14, 109. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Mas, J.; Benjak, A.; Sanseverino, W.; Bourgeois, M.; Mir, G.; González, V.M.; Hénaff, E.; Câmara, F.; Cozzuto, L.; Lowy, E. The genome of melon (Cucumis melo L.). Proc. Natl. Acad. Sci. USA 2012, 109, 11872–11877. [Google Scholar] [CrossRef] [Green Version]
- Marone, D.; Russo, M.; Laidò, G.; De Leonardis, A.; Mastrangelo, A. Plant nucleotide binding site–leucine-rich repeat (NBS-LRR) genes: Active guardians in host defense responses. Int. J. Mol. Sci. 2013, 14, 7302–7326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morata, J.; Puigdomènech, P. Variability among Cucurbitaceae species (melon, cucumber and watermelon) in a genomic region containing a cluster of NBS-LRR genes. BMC Genom. 2017, 18, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robin, A.H.K.; Larkan, N.J.; Laila, R.; Park, J.-I.; Ahmed, N.U.; Borhan, H.; Parkin, I.A.; Nou, I.-S. Korean Brassica oleracea germplasm offers a novel source of qualitative resistance to blackleg disease. Eur. J. Plant Pathol. 2017, 149, 611–623. [Google Scholar] [CrossRef]
- Hassan, M.; Rahim, M.; Natarajan, S.; Robin, A.; Kim, H.-T.; Park, J.-I.; Nou, I.-S. Gummy Stem Blight Resistance in Melon: Inheritance Pattern and Development of Molecular Markers. Int. J. Mol. Sci. 2018, 19, 2914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Gao, L.; Cao, L.; Liu, Y.; Saba, H.; Huang, Y.; Bie, Z. Assessment of suitable reference genes for quantitative gene expression studies in melon fruits. Front. Plant Sci. 2016, 7, 1178. [Google Scholar] [CrossRef] [Green Version]
- Wolukau, J.N.; Zhou, X.; Chen, J. Identification of amplified fragment length polymorphism markers linked to gummy stem blight (Didymella bryoniae) resistance in melon (Cucumis melo L.) PI 420145. HortScience 2009, 44, 32–34. [Google Scholar] [CrossRef] [Green Version]
- Kourelis, J.; van der Hoorn, R.A. Defended to the nines: 25 years of resistance gene cloning identifies nine mechanisms for R protein function. Plant Cell 2018, 30, 285–299. [Google Scholar] [CrossRef] [Green Version]
- Gururani, M.A.; Venkatesh, J.; Upadhyaya, C.P.; Nookaraju, A.; Pandey, S.K.; Park, S.W. Plant disease resistance genes: Current status and future directions. Physiol. Mol. Plant Pathol. 2012, 78, 51–65. [Google Scholar] [CrossRef]
- Brotman, Y.; Normantovich, M.; Goldenberg, Z.; Zvirin, Z.; Kovalski, I.; Stovbun, N.; Doniger, T.; Bolger, A.M.; Troadec, C.; Bendahmane, A. Dual resistance of melon to Fusarium oxysporum races 0 and 2 and to Papaya ring-spot virus is controlled by a pair of head-to-head-oriented NB-LRR genes of unusual architecture. Mol. Plant 2013, 6, 235–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuste-Lisbona, F.J.; Capel, C.; Gómez-Guillamón, M.L.; Capel, J.; López-Sesé, A.I.; Lozano, R. Codominant PCR-based markers and candidate genes for powdery mildew resistance in melon (Cucumis melo L.). Theor. Appl. Genet 2011, 122, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Dogimont, C.; Chovelon, V.; Pauquet, J.; Boualem, A.; Bendahmane, A. The V at locus encodes for a CC--NBS--LRR protein that confers resistance to Aphis gossypii infestation and A. gossypii—mediated virus resistance. Plant J. 2014, 80, 993–1004. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323. [Google Scholar] [CrossRef] [Green Version]
- Van Ooijen, G.; Mayr, G.; Kasiem, M.M.; Albrecht, M.; Cornelissen, B.J.; Takken, F.L. Structure–function analysis of the NB-ARC domain of plant disease resistance proteins. J. Exp. Bot. 2008, 59, 1383–1397. [Google Scholar] [CrossRef] [Green Version]
- Eitas, T.K.; Dangl, J.L. NB-LRR proteins: Pairs, pieces, perception, partners, and pathways. Curr. Opin. Plant Biol. 2010, 13, 472–477. [Google Scholar] [CrossRef] [Green Version]
- Martin, G.B.; Bogdanove, A.J.; Sessa, G. Understanding the functions of plant disease resistance proteins. Annu. Rev. Plant Biol. 2003, 54, 23–61. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Wang, Y.; Chen, J.-Q.; Araki, H.; Jing, Z.; Jiang, K.; Shen, J.; Tian, D. Genome-wide identification of NBS genes in japonica rice reveals significant expansion of divergent non-TIR NBS-LRR genes. Mol. Genet. Genom. 2004, 271, 402–415. [Google Scholar] [CrossRef]
- Cheng, Y.; Li, X.; Jiang, H.; Ma, W.; Miao, W.; Yamada, T.; Zhang, M. Systematic analysis and comparison of nucleotide--binding site disease resistance genes in maize. FEBS J. 2012, 279, 2431–2443. [Google Scholar] [CrossRef]
- Kang, Y.J.; Kim, K.H.; Shim, S.; Yoon, M.Y.; Sun, S.; Kim, M.Y.; Van, K.; Lee, S.-H. Genome-wide mapping of NBS-LRR genes and their association with disease resistance in soybean. BMC Plant Biol. 2012, 12, 139. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Zhang, Y.; Mason, A.S.; Lin, B.; Zhang, D.; Yu, H.; Fu, D. NBS-Encoding Genes in Brassica napus Evolved Rapidly After Allopolyploidization and Co-localize With Known Disease Resistance Loci. Front. Plant Sci. 2019, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Brotman, Y.; Silberstein, L.; Kovalski, I.; Perin, C.; Dogimont, C.; Pitrat, M.; Klingler, J.; Thompson, G.; Perl-Treves, R. Resistance gene homologues in melon are linked to genetic loci conferring disease and pest resistance. Theor. Appl. Genet. 2002, 104, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, H.; Garcia-Mas, J.; Coca, M.; Puigdoménech, P.; Monfort, A. Analysis of the melon genome in regions encompassing TIR-NBS-LRR resistance genes. Mol. Genet. Genom. 2005, 273, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Ruggieri, V.; Alexiou, K.G.; Morata, J.; Argyris, J.; Pujol, M.; Yano, R.; Nonaka, S.; Ezura, H.; Latrasse, D.; Boualem, A. An improved assembly and annotation of the melon (Cucumis melo L.) reference genome. Sci. Rep. 2018, 8, 8088. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.-H.; Choi, W.; Thomas, C.E.; Dean, R.A. Cloning of disease-resistance homologues in end sequences of BAC clones linked to Fom-2, a gene conferring resistance to Fusarium wilt in melon (Cucumis melo L.). Genome 2002, 45, 473–480. [Google Scholar] [CrossRef]
- Takahashi, H.; Shoji, H.; Ando, S.; Kanayama, Y.; Kusano, T.; Takeshita, M.; Suzuki, M.; Masuta, C. RCY1-mediated resistance to Cucumber mosaic virus is regulated by LRR domain-mediated interaction with CMV (Y) following degradation of RCY1. Mol. Plant Microbe Interact. 2012, 25, 1171–1185. [Google Scholar] [CrossRef] [Green Version]
- Larkan, N.; Lydiate, D.; Parkin, I.; Nelson, M.; Epp, D.; Cowling, W.; Rimmer, S.; Borhan, M. The Brassica napus blackleg resistance gene LepR3 encodes a receptor--like protein triggered by the Leptosphaeria maculans effector AVRLM1. New Phytol. 2013, 197, 595–605. [Google Scholar] [CrossRef]
- DeYoung, B.J.; Innes, R.W. Plant NBS-LRR proteins in pathogen sensing and host defense. Nat. Immunol. 2006, 7, 1243. [Google Scholar] [CrossRef]
- Padmanabhan, M.; Cournoyer, P.; Dinesh--Kumar, S. The leucine--rich repeat domain in plant innate immunity: A wealth of possibilities. Cell. Microbiol. 2009, 11, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Maniatis, T.; Reed, R. An extensive network of coupling among gene expression machines. Nature 2002, 416, 499. [Google Scholar] [CrossRef]
- Rose, A.B. The effect of intron location on intron--mediated enhancement of gene expression in Arabidopsis. Plant J. 2004, 40, 744–751. [Google Scholar] [CrossRef]
- Chorev, M.; Carmel, L. The function of introns. Front. Genet. 2012, 3, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.-F.; Zhu, T.; Niu, D.-K. Association of intron loss with high mutation rate in Arabidopsis: Implications for genome size evolution. Genome Biol. Evolut. 2013, 5, 723–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Lin, Y.; Chen, G.; Shen, Y.; Liu, J.; Zhang, S. Genome-scale identification of resistance gene analogs and the development of their intron length polymorphism markers in maize. Mol. Breed. 2012, 29, 437–447. [Google Scholar] [CrossRef]
- Poczai, P.; Varga, I.; Laos, M.; Cseh, A.; Bell, N.; Valkonen, J.P.; Hyvönen, J. Advances in plant gene-targeted and functional markers: A review. Plant Methods 2013, 9, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wu, W.; Jin, G.; Zhu, J. Genome-wide identification of R genes and exploitation of candidate RGA markers in rice. Chin. Sci. Bull. 2005, 50, 1120. [Google Scholar] [CrossRef]
- Slabaugh, M.B.; Yu, J.K.; Tang, S.; Heesacker, A.; Hu, X.; Lu, G.; Bidney, D.; Han, F.; Knapp, S.J. Haplotyping and mapping a large cluster of downy mildew resistance gene candidates in sunflower using multilocus intron fragment length polymorphisms. Plant Biotechnol. J. 2003, 1, 167–185. [Google Scholar] [CrossRef]
- Abuyusuf, M.; Nath, U.K.; Kim, H.-T.; Biswas, M.K.; Park, J.-I.; Nou, I.-S. Intronic Sequence Variations in a Gene with Peroxidase Domain Alter Bolting Time in Cabbage (Brassica oleracea var. capitata). Plant Mol. Biol. Rep. 2018, 36, 725–737. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl. | Gene ID | Chr. | Domain | Primer | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Amplicon Size (bp) |
|---|---|---|---|---|---|---|---|
| 1 | MELO3C028667 | chr01 | NL | MB8667 | ATGGGGTGCAATTATGGAGA | TTACTCCATTCTACACCGCG | 192 |
| 2 | MELO3C028780 | chr01 | NL | MB780 | ATGCATTCTAGGAGCAATCA | CTAGGGTAGGCTAGGATTGG | 240 |
| 3 | MELO3C028925 | chr01 | NL | MB925 | GGATTCCAGCGAGGAGATTG | CTACCGTGGATTGCAAACAC | 938 |
| 4 | MELO3C028952 | chr01 | NL | MB952 | GGATTGACATTTCCTCTGCA | CTAGGGATCATGTGGTGGTC | 488 |
| 5 | MELO3C029063 | chr01 | NL | MB063 | ATGGGCCGTGCATTCCAGGA | TCACACTCGTTGTCCTGCGG | 243 |
| 6 | MELO3C029067 | chr01 | NL | MB067 | ATGGCCTTCACGCATCCTTC | TTATTGAGTCGTCTTCGAAG | 897 |
| 7 | MELO3C028819 | chr01 | NL | MB819 | GCGGCCGCGACGAATGAACG | CCCCTTTCGCTTCGTCTTCT | 169 |
| 8 | MELO3C028832 | chr01 | NL | MB832 | GACAACCACTCTCTGAGGAT | AAGAAATCACCAAGCGTTAG | 237 |
| 9 | MELO3C028901 | chr01 | NL | MB901 | ATGAGATATGCGATCAAGTG | GCCATACATCTCCTAGCGAG | 259 |
| 10 | MELO3C029319 | chr02 | NL | MB319 | CATACCCAGTAAGGGACCCA | CCCACACTTGGGACATGACT | 696 |
| 11 | MELO3C029975 | chr03 | NL | MB975 | ATGGCGTCAGAGATAGCGTC | TCTGCTTTTCAATCTGTTCC | 666 |
| 12 | MELO3C029858 | chr03 | N | MB858 | ATGGGTTTCCGAAAGGGATT | CAGAATTTCATCAGGGATCC | 186 |
| 13 | MELO3C008456 | chr03 | NL | MB456 | CCACAAGAATACATCAAGGT | TGGTATCGCATATCTCTTCC | 289 |
| 14 | MELO3C030206 | chr03 | NL | MB206 | GGGTAAGTTGATCTTTCACA | CGCGACTCCAAGCCGACCCG | 1560 |
| 15 | MELO3C030764 | chr04 | NL | MB764 | GGTTCTTGGCTTGCAACTAG | TTAGGTGGATCCGTTGCGCG | 535 |
| 16 | MELO3C030896 | chr04 | NL | MB896 | ATGGCATTGGGAATAGGGGG | TCACACGGTCAACCAGCTTC | 279 |
| 17 | MELO3C000653 | chr05 | NL | MB653 | CTTACGGCTCGGCTCGGCTC | GTCGTCGAACGCCAAAGACC | 258 |
| 18 | MELO3C031357 | chr05 | NL | MB357 | CGGCATAAAAGACGTCGGGA | TCCCATCCTCAGAGAGTGGC | 334 |
| 19 | MELO3C031389 | chr05 | NL | MB389 | ATGCAAGAGCAATTACGGAC | ACTTCAATCCGTTCCAAAGC | 442 |
| 20 | MELO3C031417 | chr05 | NL | MB417 | ATGAATCCCACCTGGCGGCA | TTAGCTTGAACGATATTGCG | 348 |
| 21 | MELO3C004354 | chr05 | CNL | MB354-1 | ATGGCGGGAGCTTTAATTGG | ATCTAAGGTGTCTTTGAGGC | 1209 |
| MB354-2 | GCCTCAAAGACACCTTAGAT | TCATGCATATAGCCAATCCA | 1256 | ||||
| 22 | MELO3C031939 | chr06 | NL | MB939 | GCAATCACGGACGAACAAGG | TCATCTACCCAACACCTGAT | 197 |
| 23 | MELO3C023742 | chr06 | NL | MB742 | ATGAGATATGCGAGATCGTG | CTATGCCTGACTCATCTCCT | 270 |
| 24 | MELO3C014831 | chr06 | NL | MB831 | ATGCGGCGGAGGGCAGCTGG | TCATCGGTTCTCTTTCTCCT | 270 |
| 25 | MELO3C032150 | chr06 | NL | MB150 | GGATTGACACTTCCTCTACG | TTACACCTTCGGTCTTTCCA | 540 |
| 26 | MELO3C014062 | chr06 | TN | MB062-1 | ATGGATTCTGATGGGGTCGA | AACCATCTTGTATCTTTGGG | 1437 |
| MB062-2 | CCCAAAGATACAAGATGGTT | TCAGATCTTGCTCCGAAGCC | 1538 | ||||
| 27 | MELO3C032694 | chr07 | NL | MB694 | CCAAGCCTACCCCAAAGGGT | CCTCGTATGCCATTCTACAC | 194 |
| 28 | MELO3C033206 | chr08 | NL | MB206 | ATGGCCTTCACGCGTCCTTC | TAGCACTTTCTTCTGATGGG | 388 |
| 29 | MELO3C033228 | chr08 | NL | MB228 | ATGCATGAGCAATCACGAAC | CAAAAAGTTCACACCGCAAG | 519 |
| 30 | MELO3C033337 | chr08 | NL | MB337 | GCAATCATGGACGAACAAGG | TGCTCAATTAGGACTTTAGC | 349 |
| 31 | MELO3C033354 | chr08 | NL | MB354 | ATGAAGAACTTAAAGCTTGA | GGAATGACGTATTTCCAATG | 417 |
| 32 | MELO3C026319 | chr08 | NL | MB319 | TTGGGGCTCGCGACTCAGCT | TCAGCCGACAGCACTACCGT | 477 |
| 33 | MELO3C033633 | chr09 | NL | MB633 | ATGCGCATAATCAAATGCTG | ACGTCATTTTACATTGCGGG | 181 |
| 34 | MELO3C033853 | chr09 | NL | MB853 | GGATTGACACTTCCTTTTCG | TTACATGCGCATCCTCGACG | 242 |
| 35 | MELO3C033864 | chr09 | NL | MB864 | ATGCAGGAGCAATCACGAAC | CTAACGCTGCATGATTTCTT | 378 |
| 36 | MELO3C033679 | chr09 | NL | MB679 | GATAGGCACTTCCTCTGCGA | CTAGCTCACCTCGTTGCTTA | 237 |
| 37 | MELO3C012732 | chr09 | NL | MB732 | CGGCCGCGACGAAAGGACGT | CCTCTTCCGCTTCGTCTTCT | 180 |
| 38 | MELO3C033893 | chr09 | NL | MB893 | GAGATAAACCGTTCGTGTCC | TCATACGTCTTCTAACGAAC | 198 |
| 39 | MELO3C033944 | chr09 | NL | MB944 | ATGCATTCCAGCAATCACGG | CTAGGGATCATGTGGTGGTC | 585 |
| 40 | MELO3C005690 | chr09 | TN | MB690-1 | ATGGACGTTGGAGAAGAAAG | TATATCGGCTTTCGCCTCCA | 1522 |
| MB690-2 | TGGAGGCGAAAGCCGATATA | AGTTTCACGAGCAGATATCG | 1444 | ||||
| 41 | MELO3C022157 | chr09 | TNL | MB157 | ATGGAAGCAATTGAGGAATC | TACAATGACCTAGTACTCCC | 733 |
| 42 | MELO3C022154 | chr09 | TNL | MB154-1 | CGAAGATACACGTGGCGGTT | TTCTCCCATTCATCCAACCC | 1272 |
| TNL | MB154-2 | GGGTTGGATGAATGGGAGAA | TGGCCTCCTTCTCTTCTTCC | 2044 | |||
| 43 | MELO3C022152 | chr09 | TNL | MB152 | ATGGCTTCTCCAGCAACAAT | TATATAGTTACCTGATCCCG | 1664 |
| 44 | MELO3C022149 | chr09 | TNL | MB149 | CTCCTTCTCCTCCTTATTCT | TTATATCCTCACGGAGCCAC | 1241 |
| 45 | MELO3C022148 | chr09 | TNL | MB148-1 | GAAGGGGCCATCAAAGAAAT | TTGGGATGGAAATTTGGAGG | 1298 |
| TNL | MB148-2 | CCTCCAAATTTCCATCCCAA | ATGTAAAGAGAGAGAGAGAG | 998 | |||
| 46 | MELO3C022146 | chr09 | TNL | MB146-1 | GAGGCGAAGATACTCGTAAT | AAGTTCTTACTGGGAAACCC | 1090 |
| TNL | MB146-2 | GGGTTTCCCAGTAAGAACTT | GCGTGATGGAGTTGAGGGGG | 1164 | |||
| 47 | MELO3C022145 | chr09 | TNL | MB145-1 | ATGGCTGCAGGTTCCTCATC | CAAAGCTAATGGGAGTCTTC | 1329 |
| TNL | MB145-2 | GAAGACTCCCATTAGCTTTG | AACTTGGTGGAAGTTTGTGC | 1194 | |||
| TNL | MB145-3 | GCACAAACTTCCACCAAGTT | CTAGATTTGGCCTAATGTTG | 1324 | |||
| 48 | MELO3C022144 | chr09 | TNL | MB144-1 | ATGCAGAGTTCATCATCGTC | GACTATAACCAAAACTCTCC | 1426 |
| TNL | MB144-2 | GGAGAGTTTTGGTTATAGTC | GGAGAGTTTTGGTTATAGTC | 1596 | |||
| TNL | MB144-3 | GACTATAACCAAAACTCTCC | TCATTGAATTTGAGGCTCCT | 1920 | |||
| 49 | MELO3C022143 | chr09 | TNL | MB143-1 | ATGGCTTCCTCCACCACCAC | ATTCACCACAGTTATGAGGG | 1521 |
| TNL | MB143-2 | CCCTCATAACTGTGGTGAAT | TCAACCCCCATTCTCCCAAG | 1523 | |||
| 50 | MELO3C034322 | chr10 | NL | MB322 | ATGAGCTTCAGGAATACCAT | CTAACGCTTGGTGATTTCTA | 588 |
| 51 | MELO3C034765 | chr11 | NL | MB765 | CGACAAGTACAAGCAGTTCC | TCAGCTTTCGCATTTGTTCC | 165 |
| 52 | MELO3C035130 | chr11 | NL | MB130 | ATGCATTCCAAGAGCAATCA | GCATCGAAATAACTACTCCC | 298 |
| 53 | MELO3C035140 | chr11 | NL | MB140 | ATGCCTCTTCTCCGCAACCT | ATCCTTCGGGATCACTAGAC | 460 |
| 54 | MELO3C035152 | chr11 | NL | MB152 | ATGCAGGAGAAAGCACGGAC | CTACCCTCTGAGGTAGGTTG | 237 |
| 55 | MELO3C035465 | chr12 | NL | MB465 | ATGATTTCAACTTTTAATAT | CTAAATCCACCGGTGCCTGA | 309 |
| 56 | MELO3C035577 | chr12 | NL | MB577 | ATGGCTCGGATGGTTGATGG | CTTGTACTTCAATCCGTTCC | 333 |
| 57 | MELO3C035688 | chr12 | NL | MB688 | CGTCGGAAGTCGTTATTTCC | CTAGGGATCATGTGGTGGTC | 567 |
| Parent/Cross | Resistant (PIA ≤ 20) | Susceptible (PIA > 21) | Phenotypic Ratio (R:S) | χ2 | P |
|---|---|---|---|---|---|
| P1 (PI 353814; Resistant) | 20 | 0 | |||
| P2 (PI 614596; Susceptible) | 0 | 20 | |||
| F1 (PI 614596; ♀ × PI 353814; ♂) | 20 | 0 | |||
| F2 | 360 | 131 | 3:1 | 2.8 | 0.244 |
| Primer | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Amplicon Size (bp) | Primer Position |
|---|---|---|---|---|
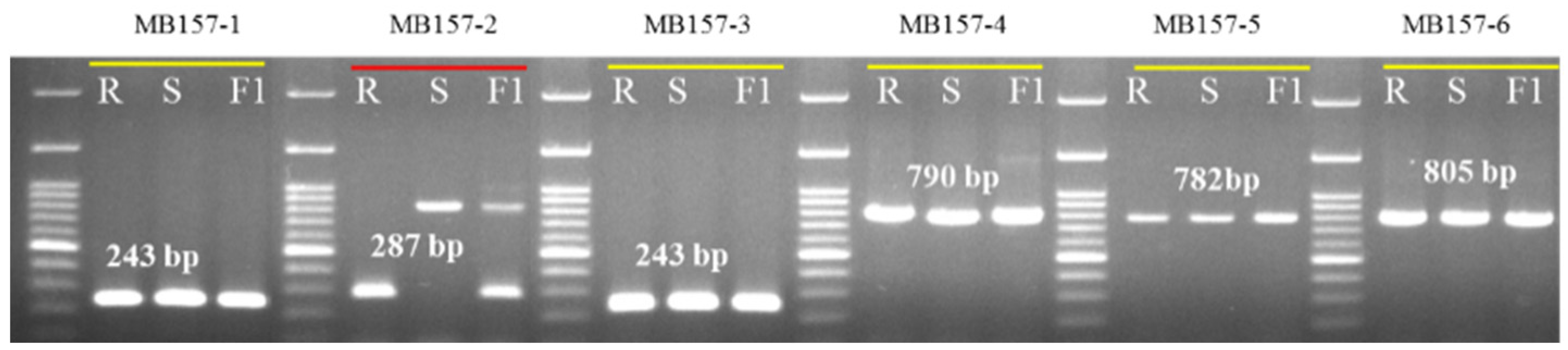
| MB157-1 | F1: ATGGAAGCAATTGAGGAATC | R1: CCGATGAACCTGCTCTTGTG | 243 | 1st. Exon |
| MB157-2 | F2: CACAAGAGCAGGTTCATCGG | R2: GGATTTTTGGTTCAAACGGT | 287 | 1st. Exon and Intron |
| MB157-3 | F3: ACCGTTTGAACCAAAAATCC | R3: TACAATGACCTAGTACTCCC | 243 | 1st. Intron |
| MB157-4 | F4: GGGAGTACTAGGTCATTGTA | R4: TCCAAGAAGTCGAGAGAGTA | 790 | 1st. Intron and 2nd. Exon |
| MB157-5 | F5: TACTCTCTCGACTTCTTGGA | R5: TATGTCGAAAGCATCTCTTC | 782 | 2nd. Exon |
| MB157-6 | F6: GAAGAGATGCTTTCGACATA | R6: TTCAATGATTGGCGACACTG | 805 | 2nd. Exon to 4th. Exon |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.R.; Hossain, M.R.; Jesse, D.M.I.; Jung, H.-J.; Kim, H.-T.; Park, J.-I.; Nou, I.-S. Development of Molecular Marker Linked with Bacterial Fruit Blotch Resistance in Melon (Cucumis melo L.). Genes 2020, 11, 220. https://doi.org/10.3390/genes11020220
Islam MR, Hossain MR, Jesse DMI, Jung H-J, Kim H-T, Park J-I, Nou I-S. Development of Molecular Marker Linked with Bacterial Fruit Blotch Resistance in Melon (Cucumis melo L.). Genes. 2020; 11(2):220. https://doi.org/10.3390/genes11020220
Chicago/Turabian StyleIslam, Md. Rafiqul, Mohammad Rashed Hossain, Denison Michael Immanuel Jesse, Hee-Jeong Jung, Hoy-Taek Kim, Jong-In Park, and Ill-Sup Nou. 2020. "Development of Molecular Marker Linked with Bacterial Fruit Blotch Resistance in Melon (Cucumis melo L.)" Genes 11, no. 2: 220. https://doi.org/10.3390/genes11020220
APA StyleIslam, M. R., Hossain, M. R., Jesse, D. M. I., Jung, H. -J., Kim, H. -T., Park, J. -I., & Nou, I. -S. (2020). Development of Molecular Marker Linked with Bacterial Fruit Blotch Resistance in Melon (Cucumis melo L.). Genes, 11(2), 220. https://doi.org/10.3390/genes11020220