1. Introduction
The bottle gourd (
Lagenaria siceraria, 2n = 2× = 22), a member of the Cucurbitaceae family [
1], is an important horticultural and medicinal crop [
2]. It originated in Africa and was independently domesticated in Africa and Asia [
3]. The fruit of bottle gourd can be used as vegetable, utensil, ornamental, and even as a musical instrument. The bottle gourd is a great source of essential nutrients, such as vitamin B complex, pectin, fibers, ascorbic acid, beta-carotene, protein, amino acids, and minerals [
4].
Bottle gourd fruit varies in shape and size [
5,
6]. Various genetic components are involved in determining fruit size and shape during fruit development [
7]. Fruit development in fleshy fruits can generally be divided into two stages, i.e., early fruit development and fruit ripening. Early fruit development involves (i) fruit setting, (ii) fruit growth by cell division, and (iii) fruit growth by cell expansion until it reaches its final size and shape [
8]. Rapid cell division and cell elongation occur during the growth stage and determine the final size and shape of the fruit [
9]. Fruit size and shape are among the most important characters of any fruit which determine its market value. Optimization of desired shape and size of the fruit demands extensive investigation of molecular mechanisms regulating fruit development.
The plant cell wall is mainly composed of polysaccharides. Hundreds of genes are involved in cell wall metabolism [
10]. Regulation of cell wall biogenesis and modification plays an important role in cell expansion and unidirectional elongation during fruit development [
11,
12]. Expansins are some of the most well characterized cell wall proteins that regulate cell size and growth during all plant growth stages and fruit development [
13]. Previous studies showed that expansins are abundantly expressed during fruit development and ripening [
14]. The cell cycle regulation is of key importance for plant growth and development. Cell division and expansion are controlled by various factors. Cyclins and cyclin-dependent protein kinases are the main regulators of the cell cycle [
15]. Many studies have shown an increase in cyclins’ expression during early fruit development in tomato and cucumber [
14,
16,
17].
Hormones also contribute to regulating fruit development and size [
18]. The seeds are rich in auxin and cytokinin, which are the main regulators of fruit development and also determine the fruit size. An increase in auxin and cytokinin levels in seeds is correlated with fruit growth stages [
19]. The role of auxin in cell expansion during fruit development stages is well established [
20,
21]. Numerous studies have demonstrated the role of auxin response factors (ARFs), the main regulators of auxin, in cell division and growth during fruit development [
20,
21,
22,
23,
24]. Transcription factors also regulate genes related to fruit development [
8,
25,
26].
Recently, transcriptomic sequencing has been used to study the molecular basis of fruit development in many fleshy fruits, including cucumber, melon, and tomato [
14,
27,
28,
29,
30,
31]. However, most of the studies focused on fruit ripening and quality, and very few specifically investigated genes related to fruit size, at least in Cucurbitaceae species. Qi et al. [
32] identified a candidate gene that encodes cyclins controlling cell proliferation and fruit size in cucumber and
C. sativus. Jiang et al. [
15] also highlighted the important role of microtubules and cyclins-related genes in determining cucumber fruit size. Later on, Wang et al. [
33] pinpointed the key role of auxin and cytokinin signaling in fruit length of cucumber. All these previous studies suggest that phytohormones, cell wall metabolism, and cell cycle and cell division are the key pathways for fruit size determination in Cucurbitaceae.
In the present study, we investigated the transcriptome of two cultivars of bottle gourd (USA and Hang) with contrasting fruit size. Our goal was to identify key pathways, structural genes, and regulators involved in the differential fruit size. It is anticipated that the information generated in this work will be helpful for bottle gourd breeding programs aimed at developing cultivars with improved fruit size.
2. Materials and Methods
2.1. Plant Materials
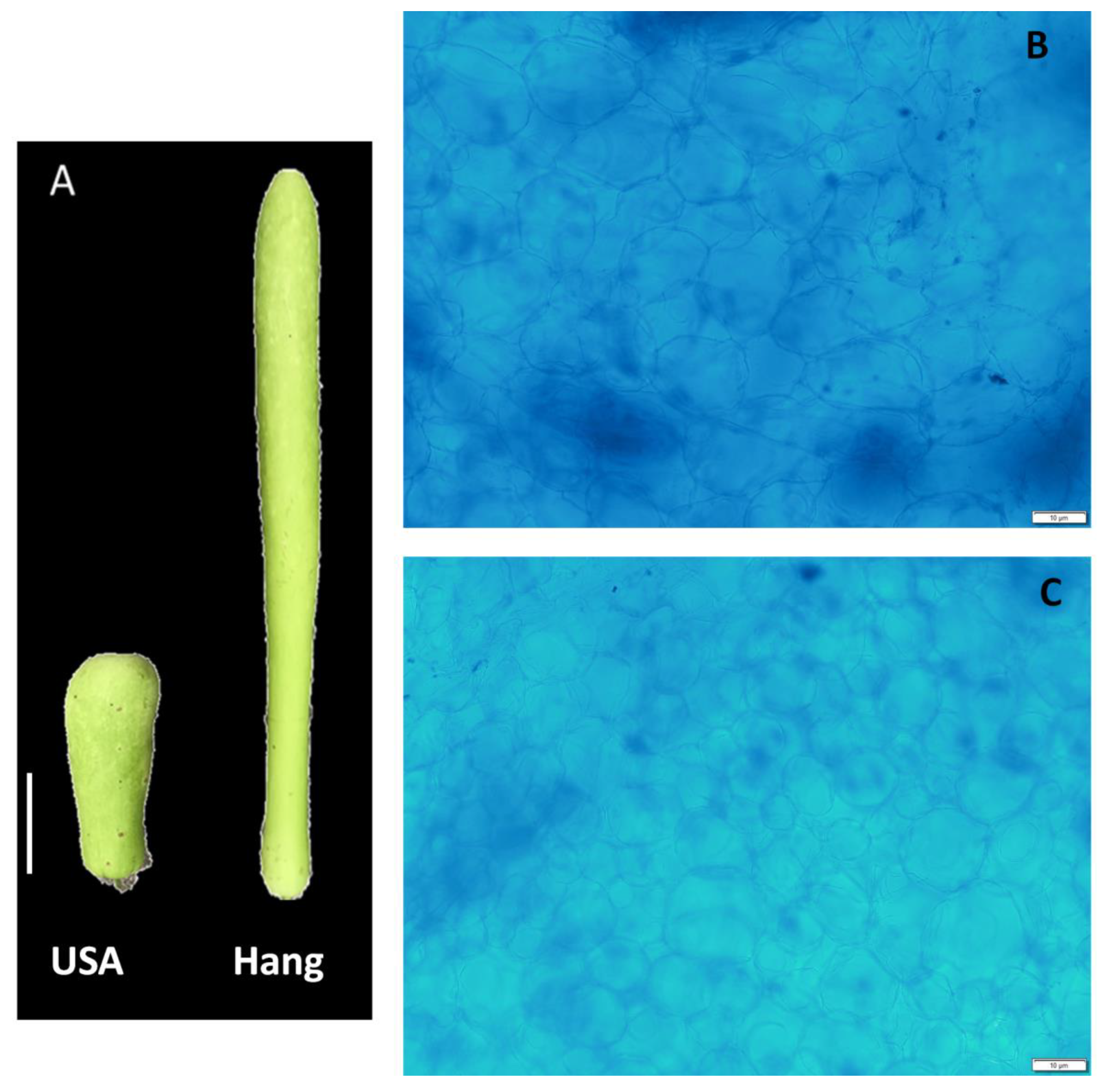
Two cultivars of bottle gourd (Lagenaria siceraria, 2n = 2× = 22 cv. Hang and USA) were used in this study. These commercial cultivars are highly grown and consumed in Wuhan, China. Fruit size of USA is considerably shorter than that of Hang. The plants were grown in the greenhouse of the Vegetable Institute of Wuhan Academy of Agricultural Sciences, Wuhan, China. Plants were grown in a substrate composed of vermiculite:perlite: organic fertilizer = 4:4:1:1. The pH of the culture substrate was about 7.0, and the water content was maintained at about 70% relative humidity. The greenhouse conditions were maintained at 25–28/18–20 °C day/night temperature, photoperiod 14 h/d with a light intensity of 87.5 µmol m−2 s−1. Pest control was performed according to standard practices. Mature fruits were harvested from three individual plants of each cultivar, and 2 cm3 of the pericarp was cut, immediately frozen in liquid nitrogen, and stored at −80 °C until further use.
2.2. Histological Analysis
A scalpel was used to cut the pericarp tissue into thin slices. The slices were fixed for 12 h in Carnoy’s fixative solution (alcohol, acetic acid; V:V 3:1), stained 2 min with 1% Janus green, and then rinsed with 70% ethanol for 10 sec. Sections were examined using an Olympus BX-50 microscope.
2.3. RNA Extraction and Quality Test
Six frozen fruit samples were ground in a mortar with liquid nitrogen, and total RNA was isolated using the RNA extraction kit (Huayueyang, China). RNA degradation and contamination were monitored on 1% agarose gels. RNA purity was checked using the NanoPhotometer® spectrophotometer (IMPLEN, CA, USA). RNA integrity was assessed using the RNA Nano 6000 Assay Kit of the Agilent Bioanalyzer 2100 system (Agilent Technologies, CA, USA).
2.4. RNA-Seq Library Preparation and Sequencing
A total amount of 3 μg RNA per sample was used as input material for the RNA sample preparations. Sequencing libraries were generated using NEBNext®Ultra™ RNA Library Prep Kit for Illumina® (NEB, Carlsbad, CA, USA) following the manufacturer’s recommendations, and index codes were added to attribute sequences to each sample. Briefly, mRNA was purified from total RNA using poly-T oligo-attached magnetic beads. Fragmentation was carried out using divalent cations under elevated temperature in NEBNext First Strand Synthesis Reaction Buffer (5×). First strand cDNA was synthesized using random hexamer primers and M-MuLV Reverse Transcripta (RNase H−). Second strand cDNA synthesis was subsequently synthesized using DNA Polymerase I and RNase H. Remaining overhangs were converted into blunt ends via exonuclease/polymerase activities. After adenylation of the 3′ ends of DNA fragments, NEBNext Adaptors with a hairpin loop structure were ligated to prepare for hybridization. To select cDNA fragments of 150~200 bp in length preferentially, the library fragments were purified with AMPure XP system (Beckman Coulter, Beverly, MA, USA). Then 3 μl USER Enzyme (NEB, Carlsbad, CA, USA) was used with size-selected, adaptor-ligated cDNA at 37 °C for 15 min followed by 5 min at 95 °C before PCR. Then PCR was performed with Phusion High-Fidelity DNA polymerase, Universal PCR primers, and Index (X) Primer. At last, PCR products were purified (AMPure XP system), and library quality was assessed on the Agilent Bioanalyzer 2100 system. The clustering of the index-coded samples was performed on a cBot Cluster Generation System using a TruSeq PE Cluster Kit v3-cBot-HS (Illumina, San Diego, CA, USA) according to the manufacturer’s instructions. After cluster generation, the library preparations were sequenced on an Illumina Hiseq 2000 platform, and paired-end reads were generated.
2.5. Quality Control and Transcriptome Assembly
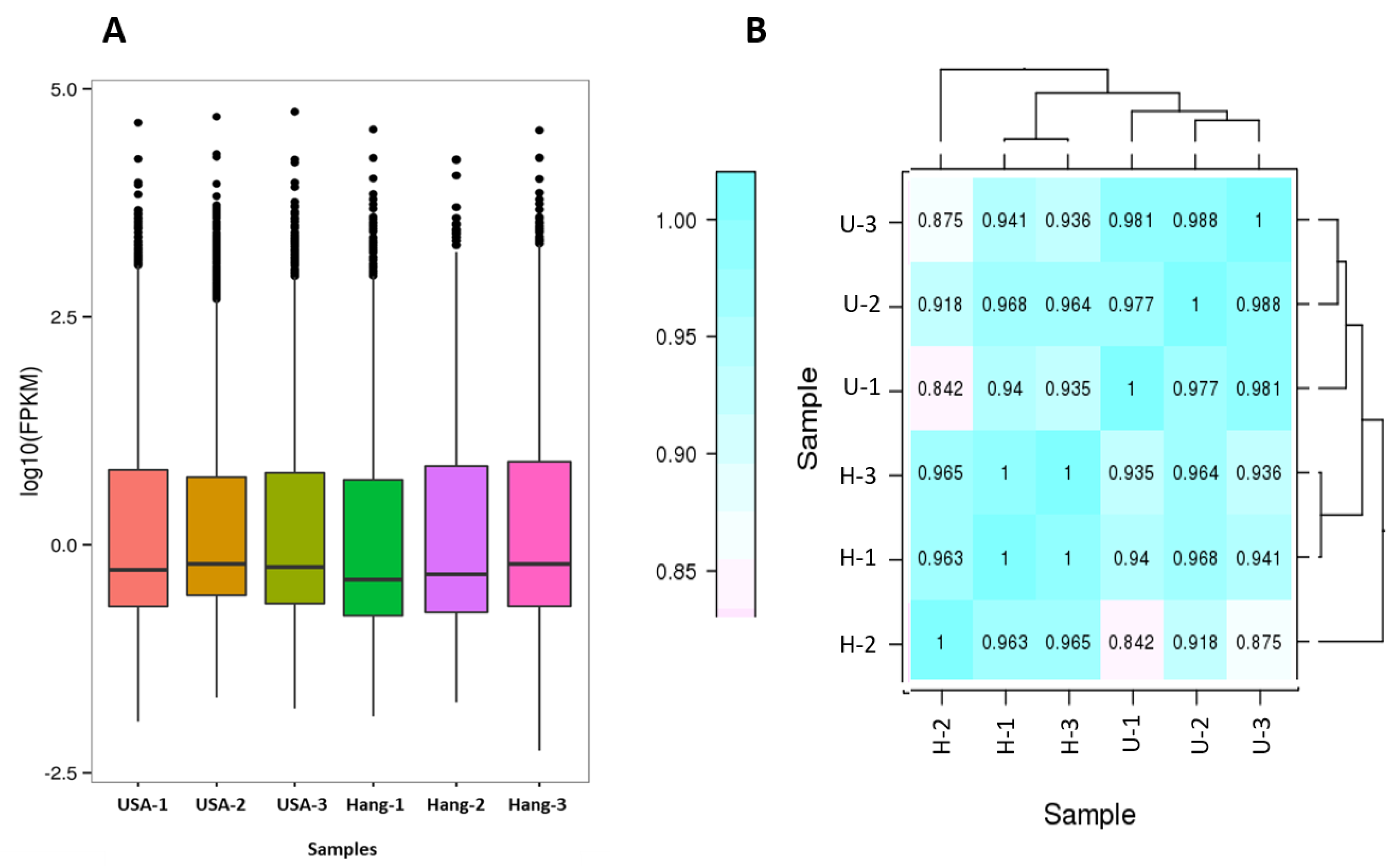
Raw data of fastq format were first processed through in-house perl scripts. In this step, clean data were obtained by removing reads containing adapters, reads containing poly-N, and low-quality reads from raw data. At the same time, Q20, Q30, GC-content, and sequence duplication level of the clean data were calculated. All the downstream analyses were based on clean data with high quality. Transcriptome assembly was accomplished using Trinity [
34] with min_kmer_cov set to 2 by default, and all other parameters set to default.
2.6. Quantification of Gene Expression and Differential Expression Analysis
Gene expression levels were estimated by RSEM [
35] for each sample. Clean data were mapped back onto the assembled transcriptome. The read count for each gene was obtained from the mapping results. Differential expression analysis was performed using the DESeq R package (1.10.1) [
36].
p-value was adjusted using
q-value [
37].
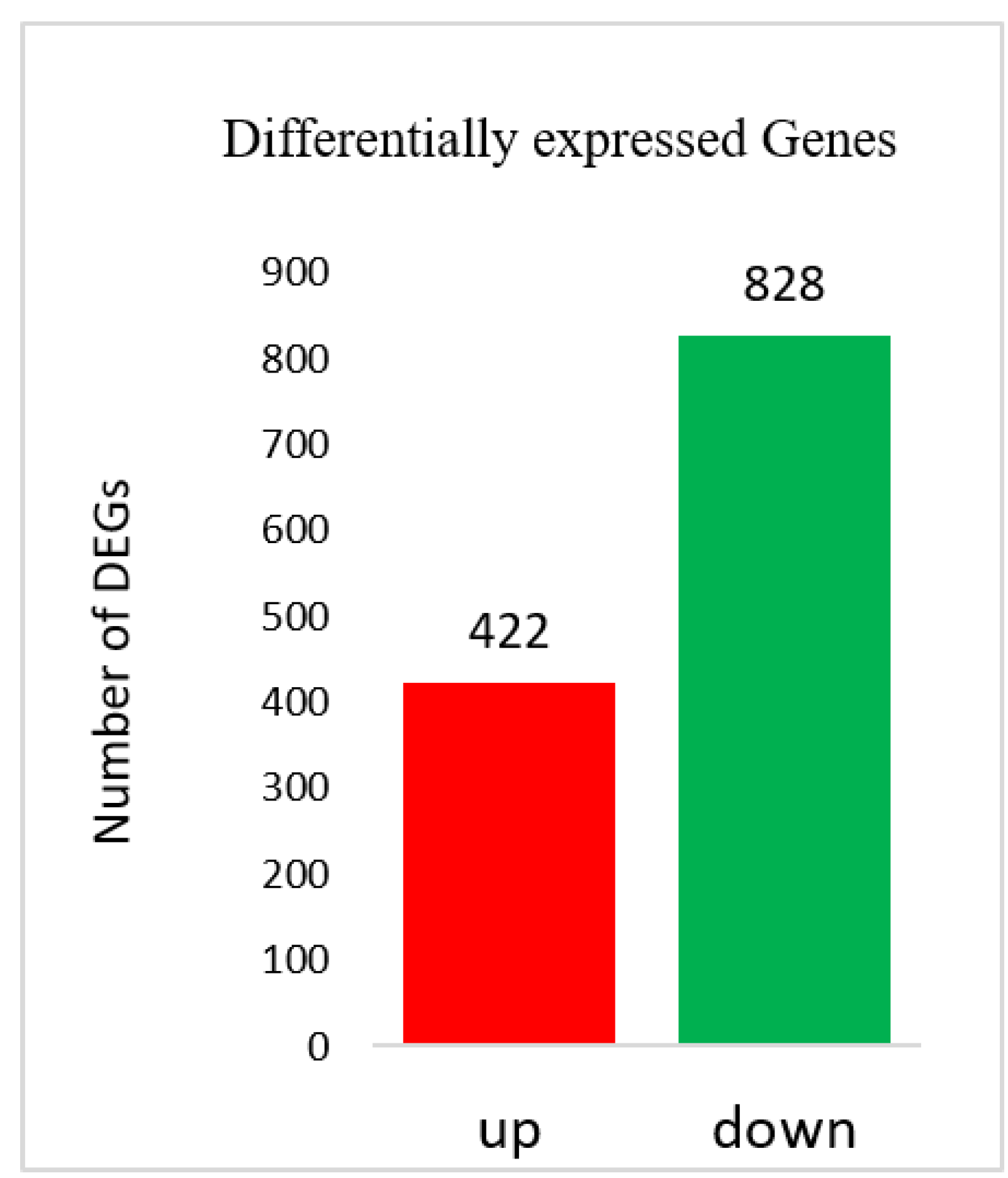
q-value < 0.005 and |log2(fold change)| >1 was set as the threshold for significantly differential expression.
2.7. GO and KEGG Pathway Enrichment Analysis
Gene ontology (GO) enrichment analysis of the differentially expressed genes (DEGs) was implemented by the topGO R packages based Kolmogorov–Smirnov test. We used KOBAS [
38] software to test the statistical enrichment of differential expression genes in Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways.
2.8. Validation of RNA-Seq Results by qRT-PCR
Total RNA was extracted from the flesh of bottle gourd USA and Hang at the mature stage. RNA-free DNaseI (Fermentas, Waltham, MA, USA) was used to remove DNA contamination for 20 min at 37 °C. Approximately 1 μg of total RNA was reverse transcribed using Oligo(dT)
18 and a Fermentas Revert Aid First Strand cDNA Synthesis Kit (Fermentas, Glen Burnie, MD, USA). The reactions were incubated for 30 min at 16 °C, followed by 60 cycles of pulsed reverse transcription at 30 °C for 30 s, 42 °C for 30 s and 50 °C for 1 s; and finally terminated by incubating at 70 °C for 5 min. Relative expression analysis of the mRNA was performed using the ABI Step One Plus™ Real Time PCR System (Applied Biosystems, Carlsbad, CA, USA) and SYBR Green Master Mix (Roche, Berlin, Germany). Quantitative real-time PCR was performed using the following parameters: 5 min at 95 °C, followed by 40 cycles of 15 s at 95 °C and 60 s at 60 °C.
LsH3 was chosen as the endogenous control [
39]. The reactions were performed with three biological replicates, and a melting curve analysis was carried out to verify that only one specific amplification occurred. Comparative expression levels were calculated in the four different samples using the 2
−ΔΔCT method. Primer sequences are provided in
Table S1.
4. Discussion
Transcriptomic sequencing of two bottle gourd accessions yielded 89,347 Unigenes. A small fraction of these Unigenes showed differential expression for bottle gourd fruit size, which indicates that few genes related to bottle gourd fruit size were expressed at the maturity stage. The mining of differentially expressed genes (DEGs) related to fruit size could be improved by sampling at early growth stages of bottle gourd fruit, and this is the objective of an ongoing study.
Fruit growth is a result of rapid cell division and cell elongation until the fruit gains its maximum size and starts ripening [
40]. Cyclins are well known for regulating cell cycle [
51]. The cell cycle is regulated at two points, G1/S and G2/M transition phases [
52]. Cyclins, along with cyclin-dependent kinases, regulate cell cycle at these two checkpoints by phosphorylation of excessive substrate, thus starting DNA replication and mitosis [
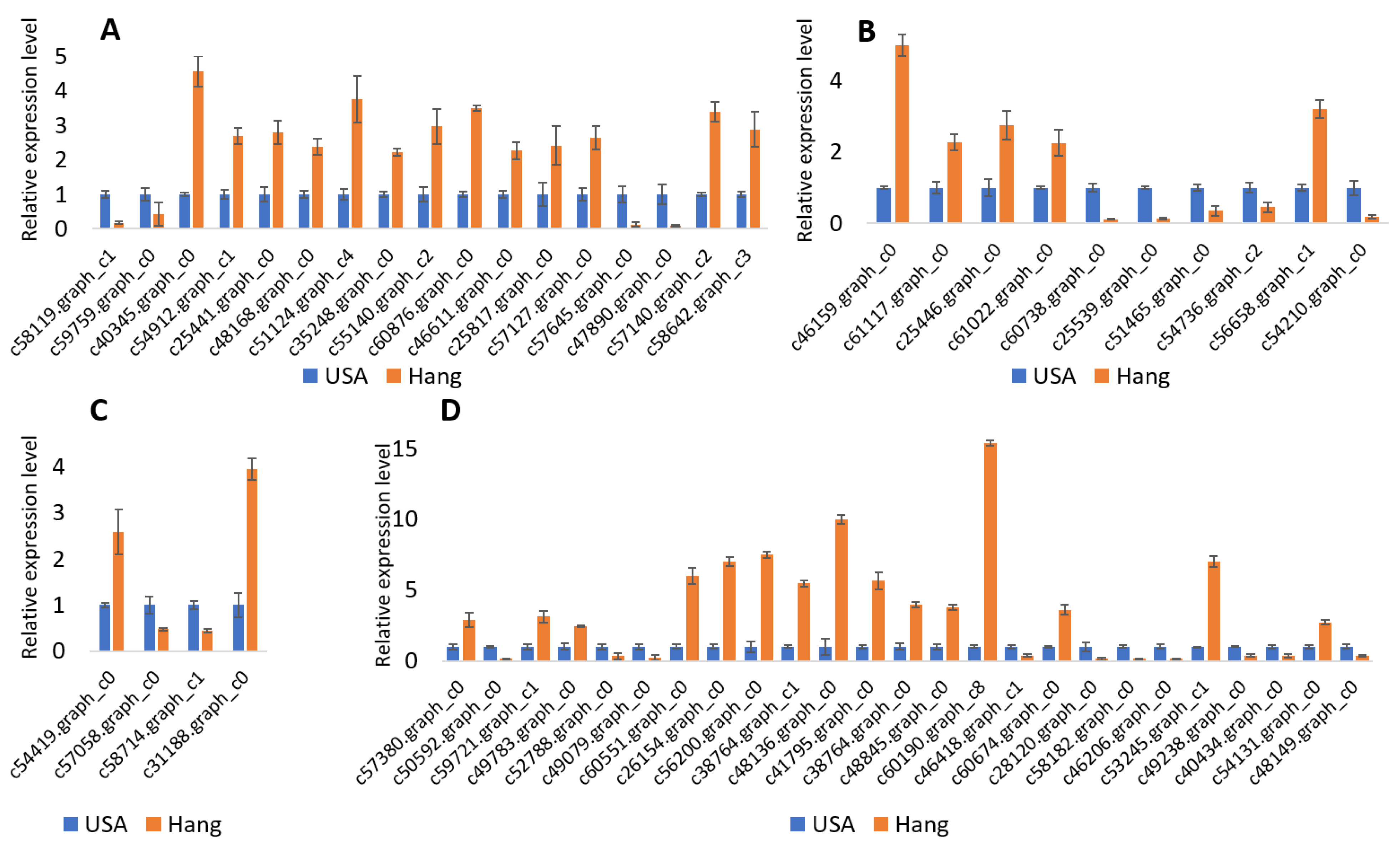
53]. We identified three cyclin genes showing significantly higher expression in the big size fruits of Hang cultivar.
c58714.graph_c1 encodes G2/mitotic-specific cyclin-1, which is essential for the regulation of the G2/M phase transition in mitosis [
54].
c54419.graph_c0 encodes Cyclin-D4-1, which is involved in the G1/S phase transition during cell division [
55]. Higher expression levels of these cyclin genes in Hang fruits suggest their positive role in fruit size determination.
The modification of the plant cell wall is highly required for the release of wall stress and a decrease in rigidity to allow cell elongation. Expansins are known for the modification of the cell wall during the growth stage. Expansins cause extension in the cell wall by disrupting the non-covalent bond between cell wall microfibrils and glucan matrix [
56]. Plant tissues under rapid growth are supposed to have a higher level of expansin gene expression [
57]. Our findings supported this idea. Four expansin genes showing significant differential expression were identified. All of these genes were highly upregulated in Hang fruits, which suggests that these genes could be important candidates for increasing bottle gourd fruit size. Further validation and characterization of these genes can be done by transgenic approaches. Endoglucanases are involved in endohydrolysis of (1->4)-β-D-glucosidic linkages in cellulose and contribute to cellulose microfibril formation and cell wall organization during elongation. Six DEGs related to endoglucanase were identified. Five of these were upregulated in Hang fruits. Xyloglucans are an essential component of the cell wall. Xyloglucan endotransglucosylase/hydrolases (XTH) cleaves and relegates xyloglucan polymers and participates in cell wall construction in growing tissues. Four XTH genes were differentially regulated in the two cultivars, including up- and downregulated genes. Recently, Han et al. [
58] found that two members
DkXTH6 and
DkXTH7 of the XTH family, have opposite expression patterns and play special and divergent roles in persimmon fruit. While one takes part in cell wall restructuring, the other is involved in cell wall assembly. In this study, differential expression of cell wall modification-related genes between the two bottle gourd cultivars suggests their active role in regulating the fruit size in bottle gourd. Further investigations will be required to elucidate the specific function of each DEG and their impact on fruit size.
Hormones also contribute to the regulation of fruit development. Cytokinin accelerates cell division during the growth stage [
59]. Cytokinin dehydrogenase (CKX) breaks down the cytokinin [
60]. Overexpression of cytokinin dehydrogenase in Arabidopsis led to alterations in plant growth [
61]. Two CKX genes were downregulated in Hang fruits suggesting that these genes negatively regulate fruit size. The role of auxin in plant cell expansion is well established [
20]. Many studies have demonstrated the role of auxin response factors (ARFs), the main regulators of auxin, in cell division and plant growth [
21,
23,
24,
62]. We identified eight auxin-related genes showing significant differential expression among the two cultivars. These genes represent important resources to be further characterized to uncover their specific roles in the regulation of fruit size in bottle gourd.
Although several studies were conducted to identify quantitative trait loci related to fruit size and shape in bottle gourd [
63], no putative gene has been specifically reported so far. Well characterized genes related to fruit size were identified in other plants, such as tomato and Arabidopsis. Based on our transcriptome assembly, we successfully identified several homologs in bottle gourd. Fruit weight 2.2 is a negative regulator of cell division and determines fruit size in tomato [
48]. In our study, the expression of the homologous gene was significantly repressed in Hang. In accordance with the notion that Cell Size Regulator (
CSR) positively controls cell size and fruit weight, we observed an upregulation of
CSR in Hang [
49]. Furthermore,
AtHXK-1 activity has been shown to perturb carbon and energy metabolism and reduce fruit and seed size in tomato [
50]. The homologous gene found within our DEGs showed an upregulation in the small-size fruits of USA. Therefore, we deduce that the molecular mechanisms related to fruit size in other well studied plants may be conserved in bottle gourd, and the homology-based candidate gene mining could be an efficient way to pinpoint important gene resources [
64].
Regulation of the expression levels of structural genes is under the tight control of transcription factor (TF) [
8]. In this study, the upregulation of several ERFs in Hang fruit denotes that they positively contribute to the big fruit size. Numerous studies identified ERF genes as master regulators of the expression levels of phytohormones, cell wall modification, cell cycle, and other important structural genes involved in fruit development and ripening [
65,
66,
67]. Decoding the networks connecting ERFs and other TFs identified in the present work to their target structural genes will highlight the most important genes to target for efficient fruit size manipulation in bottle gourd [
68].






{kind=link}
{kind=link}
{kind=link}
{kind=link}