Major QTLs for Trunk Height and Correlated Agronomic Traits Provide Insights into Multiple Trait Integration in Oil Palm Breeding



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and DNA Preparation
2.2. Trait Recording and Statistical Analyses
2.3. DNA Marker Genotyping
2.4. Construction of Linkage Maps
2.5. Quantitative Trait Locus Mapping Analyses
2.6. Cumulative Effects of Quantitative Trait Allele Analysis
3. Results
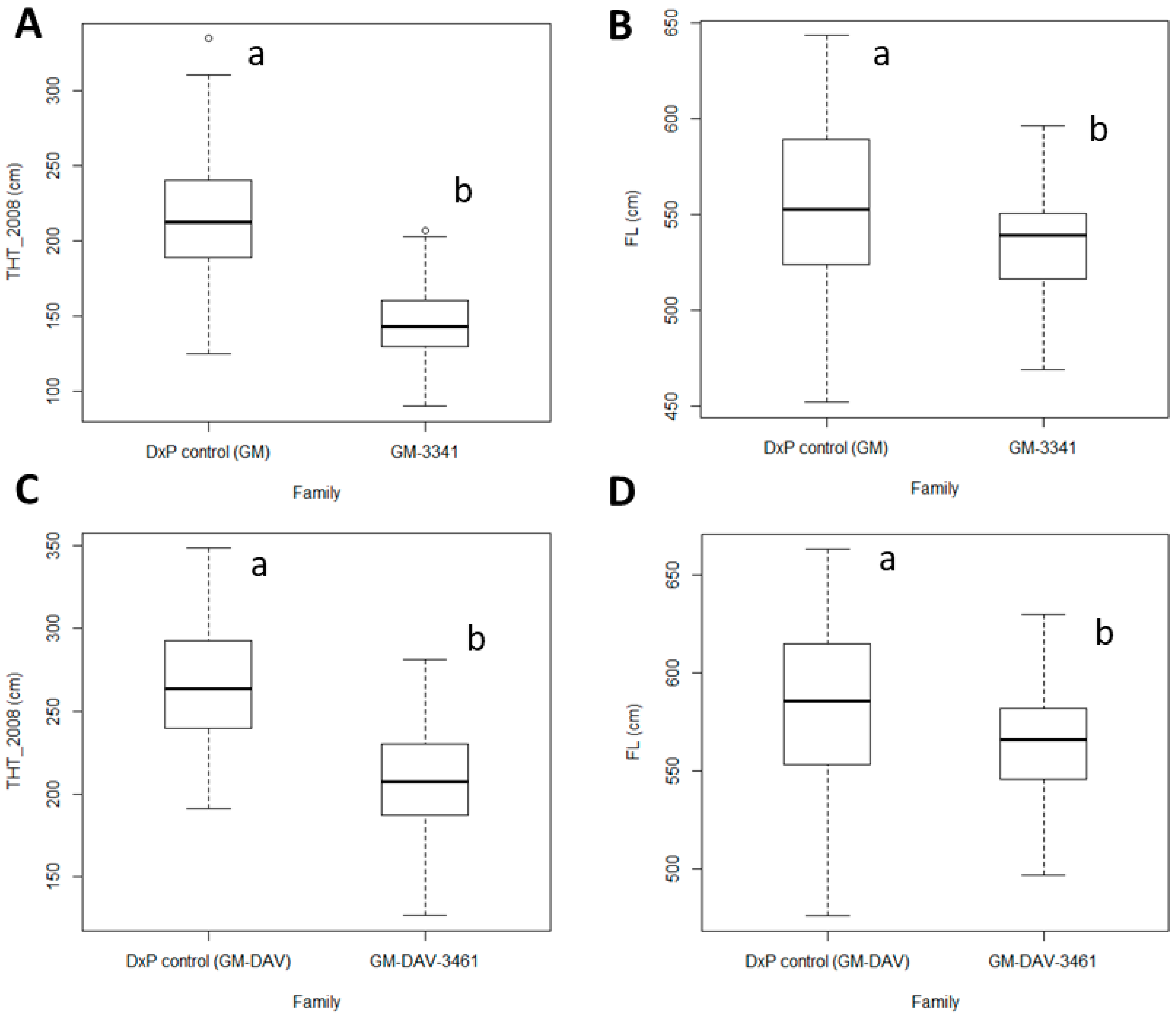
3.1. Descriptive Statistics and Comparison of Traits between Families
3.2. Pearson Correlation among Traits
3.3. Construction of Linkage Maps
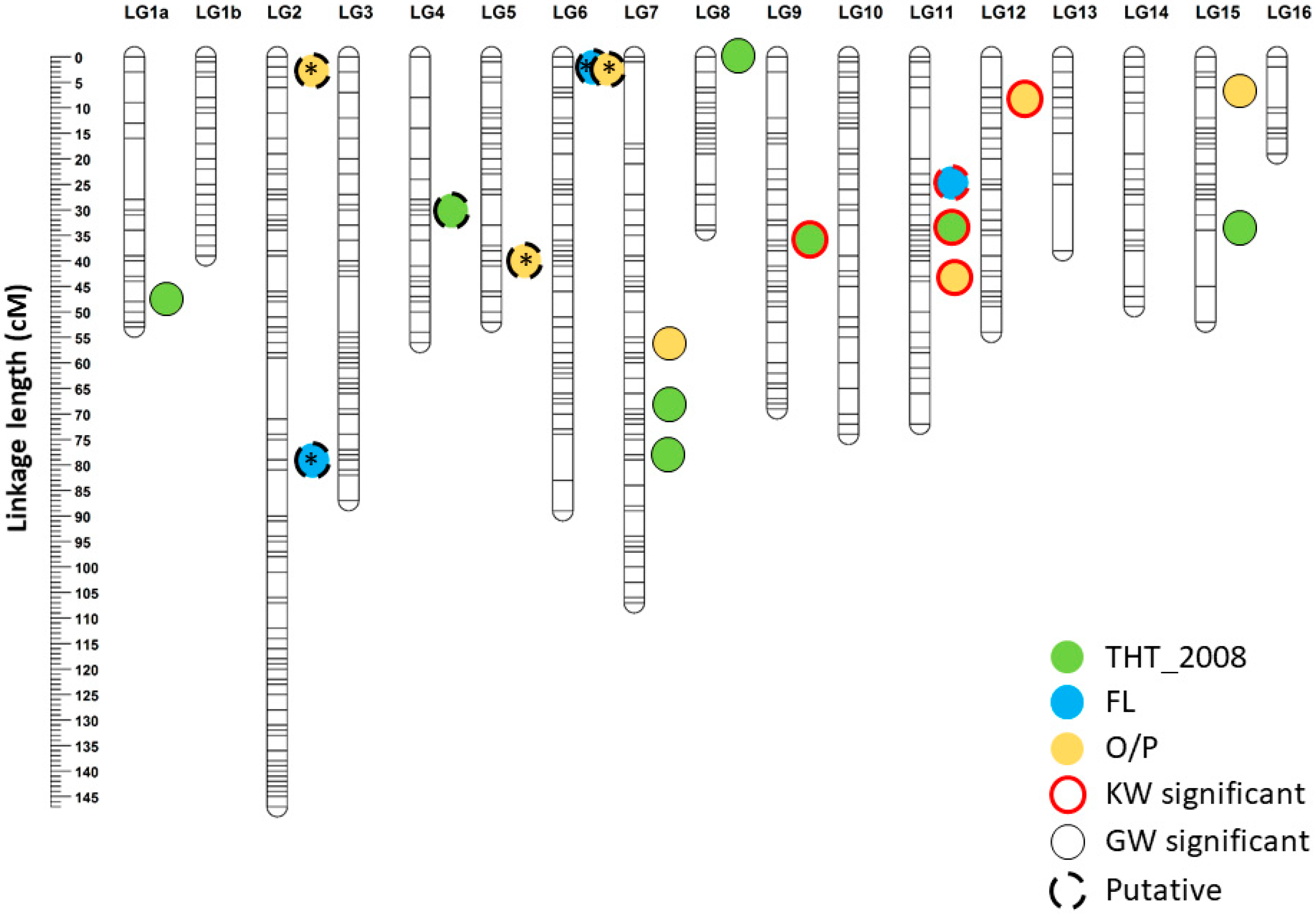
3.4. Mapping of QTLs for THT, FL and O/P Traits
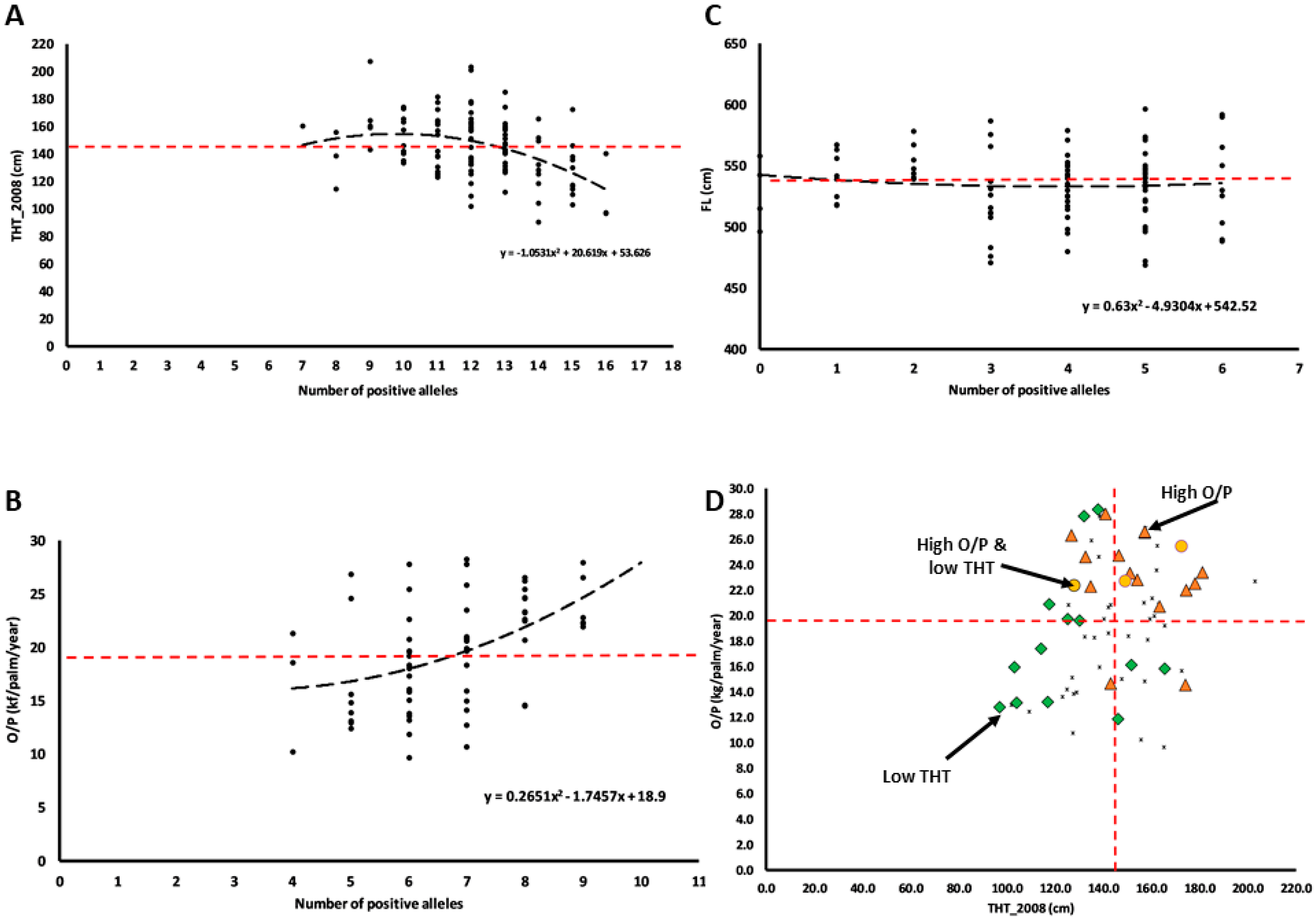
3.5. Cumulative Effects of Quantitative Trait Allele Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- OECD/FAO. OECD-FAO Agricultural Outlook 2019–2028; OECD Publishing: Paris, France; Food and Agriculture Organization of the United Nations: Rome, Italy, 2019. [Google Scholar]
- Sayer, J.; Ghazoul, J.; Nelson, P.; Klintuni Boedhihartono, A. Oil palm expansion transforms tropical landscapes and livehoods. Glob. Food Secur. 2012, 1, 114–119. [Google Scholar] [CrossRef]
- ISTA Mielke. Oil World; ISTA Mielke: Hamburg, Germany, 2018. [Google Scholar]
- Hanim, A. Help for palm oil industry. Star 2012. Available online: https://www.thestar.com.my/business/business-news/2012/05/15/help-for-palm-oil-industry (accessed on 5 May 2020).
- Diana, R. Manpower still a problem in oil palm industry. Star 2013. Available online: https://www.thestar.com.my/news/community/2013/07/24/manpower-still-a-problem-more-locals-needed-to-mitigate-labour-shortage-in-oil-palm-industry/ (accessed on 5 May 2020).
- Le Guan, V.; Ouattara, S.; Jacquemard, J.C. Oil palm selection with a view to easier harvesting. Initial results. Oléagineux 1990, 45, 523–531. [Google Scholar]
- Barcelos, E.; Rios, S.d.A.; Cunha, R.N.V.; Lopes, R.; Motoike, S.Y.; Babiychuk, E.; Skirycz, A.; Kushnir, S. Oil palm natural diversity and the potential for yield improvement. Front. Plant Sci. 2015, 6, 190. [Google Scholar] [CrossRef] [PubMed]
- Jagoe, R.B. The dumpy oil palm Malay. Agric. J. 1952, 35, 12. [Google Scholar]
- Soh, A.C.; Vanialingam, T.; Taniputra, B.; Pamin, K. Derivatives of the dumpy palm—Some experimental results. Plant 1981, 57, 227–239. [Google Scholar]
- Nair, K.P.P. Oil Palm (Elaeis guineensis Jacquin). In The Agronomy and Economy of Important Tree Crops of the Developing World; Nair, K.P.P., Ed.; Elsevier: London, UK, 2010; pp. 209–236. [Google Scholar] [CrossRef]
- Lee, C.H.; Pang, T.Y. Breeding for short height increment in oil palm. In Proceedings of the International Symposium of Oil Palm Genetic Resources and Utilization, Kuala Lumpur, Malaysia, 8–10 June 2000; p. 12. [Google Scholar]
- Kwong, Q.B.; Teh, C.K.; Ong, A.L.; Heng, H.Y.; Lee, H.L.; Mohaimi, M.; Low, J.Z.; Sukganah, A.; Chew, F.T.; Mayes, S.; et al. Development and validation of a high-density SNP genotyping array for african oil palm. Mol. Plant 2016, 9, 1132–1141. [Google Scholar] [CrossRef]
- Corley, R.H.V.; Tinker, P.B. Selection and breeding. In The Oil Palm, 3rd ed.; Blackwell: Oxford, UK, 2003; pp. 133–187. [Google Scholar]
- Blaak, G.; Sparnaaij, L.D.; Menendez, T. Methods of bunch analysis. In Breeding and Inheritance in the Oil Palm (Elaeis Guineensis Jacq.) Part II; Springer: Berlin, Germany, 1963; Volume 4, pp. 146–155. [Google Scholar]
- Rao, V.; Soh, A.C.; Corley, R.H.V.; Lee, C.H.; Rajanaidu, N.; Tan, Y.P.; Chin, C.W.; Lim, K.C.; Tan, S.T.; Lee, T.P.; et al. A critical reexamination of the method of bunch analysis in oil palm breeding. Palm Oil Res. Inst. Malays. Occ. Pap. 1983, 9, 1–28. [Google Scholar]
- Corley, R.H.V.; Hardon, J.J.; Tan, G.Y. Analysis of growth of the oil palm (Elaeis guineensis Jacq.) I. Estimation of growth parameters and application in breeding. Euphytica 1971, 20, 307–315. [Google Scholar] [CrossRef]
- Minitab, Inc. Minitab17; Computer Software; Minitab, Inc.: State College, PA, USA, 2016. [Google Scholar]
- Blair, M.W.; Hedetale, V.; McCouch, S.R. Fluorescent-labeled microsatellite panels useful for detecting allelic diversity in cultivated rice (Oryza sativa L.). Theor. Appl. Genet. 2002, 105, 449–457. [Google Scholar] [CrossRef]
- Rastas, P.; Paulin, L.; Hanski, I.; Lehtonen, R.; Auvinen, P. Lep-MAP: Fast and accurate linkage map construction for large SNP datasets. Bioinformatics 2013, 29, 3128–3134. [Google Scholar] [CrossRef]
- Rastas, P. Lep-MAP3: Robust linkage mapping even for low-coverage whole genome sequencing data. Bioinformatics 2017, 33, 3726–3732. [Google Scholar] [CrossRef]
- Van Ooijen, J.W. Software for the Calculation of Genetic Linkage Maps in Experimental Populations of Diploid Species; Kyazma, B.V.: Wageningen, The Netherlands, 2018. [Google Scholar]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef]
- Singh, R.; Ong-Abdullah, M.; Low, E.T.L.; Manaf, M.A.A.; Rosli, R.; Nookiah, R.; Ooi, L.C.L.; Ooi, S.E.; Chan, K.L.; Halim, M.A.; et al. Oil palm genome sequence reveals divergence of interfertile species in Old and New worlds. Nature 2013, 500, 335–339. [Google Scholar] [CrossRef]
- Van Ooijen, J.W. MapQTL 6, Software for the Mapping of Quantitative Trait Loci in Experimental Populations of Diploid Species; Kyazma, B.V.: Wageningen, The Netherlands, 2009. [Google Scholar]
- Pamin, K.A. A hundred and fifty years of oil palm in Indonesia: From the Bogor Botanical Garden to the industry. In Proceedings of the International Oil Palm Conference ‘Commodity of the Past, Today and the Future’, Bali, Indonesia, 23–25 September 1998; pp. 3–23. [Google Scholar]
- Kushairi, A.; Rajanaidu, N. Breeding populations, seed production and nursery management. In Advances in Oil Palm Research; Basiron, Y., Jalani, B.S., Chan, K.W., Eds.; Malaysian Palm Oil Board: Kuala Lumpur, Malaysia, 2000; Volume 1, pp. 39–98. [Google Scholar]
- Rosenquist, E.A. The genetic base of oil palm breeding populations. In Proceedings of the International Workshop on Oil Palm Germplasm and Ultilization, Bangi, Malaysia, 26–27 March 1985; pp. 27–56. [Google Scholar]
- Beirnaert, A.; Vanderweyen, R. Contribution à l’étude génétique et biométrique des variétiés d’Elaeis guineensis Jacq. Publ. Inst. Nat. Etude Agron. Congo Belge Ser. Sci. 1941, 3, 95–110. [Google Scholar]
- Hardon, J.J.; Corley, R.H.V.; Lee, C.H. Breeding and Selecting the Oil Palm; Academic Press: London, UK, 1987; pp. 63–81. [Google Scholar]
- Mohd Basri, W.; Norman, K. Role and effectiveness of Elaeidobius kamerunicus, Thrips hawaiiensis and Pyroderces sp. in pollination of mature oil palm in Peninsular Malaysia. J. Oil Palm Res. 1997, 9, 1–16. [Google Scholar]
- Cik Mohd Rizuan, Z.A.; Noor Hisham, H.; Samsudin, A. Role of pollinating weevil (Elaeidobius kamerunicus), seasonal effect and its relation to fruit set in oil palm area of FELDA. In Proceedings of the PIPOC 2013 Conference, Kuala Lumpur, Malaysia, 19–21 November 2013. [Google Scholar]
- Henson, I.E. Measuring and modelling oil palm trunk growth. Oil Palm Bull. 2006, 52, 1–24. [Google Scholar]
- Jacquemard, J.C. Contribution to the study of the height growth of the stems of Elaeis guineensis Jacq. Study of the L2T × D10D cross. Oléagineux 1979, 34, 492–497. [Google Scholar]
- Corley, R.H.V.; Tinker, P.B. Growth, flowring and yield. In The Oil Palm, 4th ed.; Blackwell Science Ltd.: Oxford, UK, 2016; pp. 89–134. [Google Scholar]
- Kwan, B.K.W. The effect of planting density on the first fifteen years of growth and yield of oil palm in Sabah. Tech. Bull. 1994, 11, 26. [Google Scholar]
- Henson, I.E.; Mohd Tayeb, D. Physiological analysis of an oil palm density trial on a peat soil. J. Oil Palm Res. 2003, 15, 1–27. [Google Scholar]
- Weraduwage, S.M.; Chen, J.; Anozie, F.C.; Morales, A.; Weise, S.E.; Sharkey, T.D. The relationship between leaf area growth and biomass accumulation in Arabidopsis thaliana. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Henson, I.E. Photosynthesis and source-sink relationship in oil palm. Trans. Malays. Soc. Plant Physiol. 1990, 1, 165–171. [Google Scholar]
- Henson, I.E. Age-related changes in stomatal and photosynthetic characteristics of leaves of oil palm (Elaeis guineensis Jacq.). Elaeis 1991, 4, 51–59. [Google Scholar]
- Ong, A.-L.; Teh, C.-K.; Kwong, Q.-B.; Tangaya, P.; Appleton, D.R.; Massawe, F.; Mayes, S. Linkage-based genome assembly improvement of oil palm (Elaeis guineensis). Sci. Rep. 2019, 9, 6619. [Google Scholar] [CrossRef]
- Argyris, J.M.; Ruiz-Herrera, A.; Madriz-Masis, P.; Sanseverino, W.; Morata, J.; Pujol, M.; Ramos-Onsins, S.E.; Garcia-Mas, J. Use of targeted SNP selection for an improved anchoring of the melon (Cucumis melo L.) scaffold genome assembly. BMC Genom. 2015, 16, 4. [Google Scholar] [CrossRef]
- Mathew, L.S.; Spannagl, M.; Al-Malki, A.; George, B.; Torres, M.F.; Al-Dous, E.K.; Al-Azwani, E.K.; Hussein, E.; Mathew, S.; Mayer, K.F.X.; et al. A first genetic map of date palm (Phoenix dactylifera) reveals long-range genome structure conservation in the palms. BMC Genom. 2014, 15, 285. [Google Scholar] [CrossRef]
- Xu, Z.; Zou, F.; Vision, T.J. Improving quantitative trait loci mapping resolution in experimental crosses by the use of genotypically selected samples. Genetics 2005, 170, 401–408. [Google Scholar] [CrossRef]
- Balyan, H.S.; Singh, O. Pleiotropic effects of GA-insensitive Rht Genes on grain yield and its component characters in wheat. Cereal Res. Commun. 1994, 22, 195–200. [Google Scholar]
- Jobson, E.M.; Johnston, R.E.; Oiestad, A.J.; Martin, J.M.; Giroux, M.J. The Impact of the wheat Rht-B1b semi-dwarfing allele on photosynthesis and seed development under field conditions. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Pootakham, W.; Jomchai, N.; Ruang-areerate, P.; Shearman, J.R.; Sonthirod, C.; Sangsrakru, D.; Tragoonrung, S.; Tangphatsornruang, S. Genome-wide SNP discovery and identification of QTL associated with agronomic traits in oil palm using genotyping-by-sequencing (GBS). Genomics 2015, 105, 288–295. [Google Scholar] [CrossRef]
- Lee, M.; Xia, J.H.; Zou, Z.; Ye, J.; Rahmadsyah; Alfiko, Y.; Jin, J.; Lieando, J.V.; Purnamasari, M.I.; Lim, C.H.; et al. A consensus linkage map of oil palm and a major QTL for stem height. Sci. Rep. 2015, 5, 8232. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Trait | Trait Abbreviation | THT_2008 | THT_2014 |
|---|---|---|---|
| Fresh fruit bunch | FFB/kg | 0.423 * | 0.242 * |
| Bunch number | BNO | 0.364 * | 0.119 |
| Bunch weight | BWT | 0.248 * | 0.156 |
| Fruit-to-bunch ratio | F/B | −0.055 | −0.015 |
| Mean fruit weight | MFW | −0.068 | 0.107 |
| Mesocarp-to-fruit ratio | M/F | −0.150 | 0.103 |
| Shell-to-fruit ratio | S/F | 0.156 | −0.153 |
| Kernel-to-fruit ratio | K/F | 0.033 | 0.043 |
| Oil-to-dry mesocarp ratio | O/DM | 0.237 | 0.216 |
| Oil-to-wet mesocarp ratio | O/WM | −0.038 | −0.042 |
| Oil-to-bunch ratio | O/B | 0.126 | 0.069 |
| Kernel-to-bunch ratio | K/B | 0.081 | −0.005 |
| Total oil per palm | O/P | 0.294 * | 0.165 |
| Frond length | FL | 0.270 * | 0.301 * |
| Trunk height in 2008 | THT_2008 | NA | 0.595 * |
| Trunk height in 2014 | THT_2014 | NA | NA |
| No. | Group | Linkage Length (cM) | No. of Linked Markers | Mapping Interval (cM) | No. of Markers in the Physical Map | Linkage Length of Common Markers (cM) | Physical Length (Mb) | Average Recombination Rate (cM/Mb) |
|---|---|---|---|---|---|---|---|---|
| 1 | LG1a | 53.44 | 20 | 2.67 | 10 | 53.44 | 63.71 | 0.84 |
| 2 | LG1b | 38.99 | 20 | 1.95 | 18 | 38.99 | 61.63 | 0.63 |
| 3 | LG2 | 147.43 | 73 | 2.02 | 36 | 147.43 | 64.78 | 2.28 |
| 4 | LG3 | 86.94 | 48 | 1.81 | 27 | 86.94 | 58.66 | 1.48 |
| 5 | LG4 | 55.97 | 32 | 1.75 | 9 | 55.97 | 34.73 | 1.61 |
| 6 | LG5 | 52.11 | 34 | 1.53 | 20 | 52.11 | 37.29 | 1.40 |
| 7 | LG6 | 88.89 | 55 | 1.62 | 22 | 83.33 | 41.90 | 1.99 |
| 8 | LG7 | 107.34 | 49 | 2.19 | 27 | 105.61 | 42.27 | 2.50 |
| 9 | LG8 | 33.67 | 28 | 1.20 | 22 | 33.67 | 39.98 | 0.84 |
| 10 | LG9 | 69.28 | 38 | 1.82 | 23 | 68.16 | 34.66 | 1.97 |
| 11 | LG10 | 74.31 | 35 | 2.12 | 19 | 74.31 | 22.94 | 3.24 |
| 12 | LG11 | 72.36 | 37 | 1.96 | 13 | 72.36 | 26.05 | 2.78 |
| 13 | LG12 | 54.00 | 28 | 1.93 | 26 | 54.00 | 26.63 | 2.03 |
| 14 | LG13 | 37.90 | 11 | 3.45 | 7 | 23.12 | 25.51 | 0.91 |
| 15 | LG14 | 49.50 | 23 | 2.15 | 18 | 49.50 | 23.78 | 2.08 |
| 15 | LG15 | 51.95 | 22 | 2.36 | 16 | 45.24 | 22.25 | 2.03 |
| 17 | LG16 | 19.43 | 12 | 1.62 | 9 | 16.43 | 17.05 | 0.96 |
| GW | 1093.51 * | 565 * | 2.01 * | 322 * | 1060.61 * | 643.82 * | 1.74 ** |
| Trait | Linkage Group | Cofactor | Peak LOD | Peak Position (cM) | Trait Variation Explained (%) | KW |
|---|---|---|---|---|---|---|
| THT_2008 | 1a | rs795987799 | 4.55 | 48.04 | 7.6 | |
| 4 | rs795956088 | 2.67 | 30.22 | 4.3 | ||
| 7 | rs796035893 | 6.65 | 68.67 | 11.7 | ||
| 7 | ss1810504935 | 6.79 | 78.32 | 12.0 | ||
| 8 | rs795967221 | 5.02 | 0.00 | 8.5 | ||
| 9 | rs796032489 | 4.42 | 35.56 | 7.4 | ***** | |
| 11 | ss1810516343 | 9.11 | 33.55 | 17.0 | **** | |
| 15 | rs795995560 | 5.99 | 33.79 | 10.4 | ||
| O/P | 2 | DA290 NC | 3.74 | 81.14 | 7.6 | |
| 5 | rs796007949 NC | 2.88 | 39.55 | 6.0 | ||
| 6 | DA358 NC | 3.02 | 88.89 | 6.3 | ||
| 7 | ss1810302303 | 4.09 | 56.03 | 10.9 | ||
| 11 | rs796015770 | 4.56 | 43.79 | 12.4 | **** | |
| 12 | rs796017329 | 8.13 | 34.73 | 25.2 | **** | |
| 15 | rs795992318 | 8.92 | 16.25 | 28.5 | ||
| FL | 2 | rs795951455 NC | 2.69 | 79.40 | 10.2 | |
| 6 | rs795993306 NC | 3.11 | 2.26 | 11.7 | ||
| 6 | ss1810567844 NC | 3.11 | 2.38 | 11.7 | ||
| 11 | rs795990373 | 3.73 | 24.55 | 16.5 | ****** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teh, C.-K.; Ong, A.-L.; Mayes, S.; Massawe, F.; Appleton, D.R. Major QTLs for Trunk Height and Correlated Agronomic Traits Provide Insights into Multiple Trait Integration in Oil Palm Breeding. Genes 2020, 11, 826. https://doi.org/10.3390/genes11070826
Teh C-K, Ong A-L, Mayes S, Massawe F, Appleton DR. Major QTLs for Trunk Height and Correlated Agronomic Traits Provide Insights into Multiple Trait Integration in Oil Palm Breeding. Genes. 2020; 11(7):826. https://doi.org/10.3390/genes11070826
Chicago/Turabian StyleTeh, Chee-Keng, Ai-Ling Ong, Sean Mayes, Festo Massawe, and David Ross Appleton. 2020. "Major QTLs for Trunk Height and Correlated Agronomic Traits Provide Insights into Multiple Trait Integration in Oil Palm Breeding" Genes 11, no. 7: 826. https://doi.org/10.3390/genes11070826
APA StyleTeh, C.-K., Ong, A.-L., Mayes, S., Massawe, F., & Appleton, D. R. (2020). Major QTLs for Trunk Height and Correlated Agronomic Traits Provide Insights into Multiple Trait Integration in Oil Palm Breeding. Genes, 11(7), 826. https://doi.org/10.3390/genes11070826