RNA Editing and Modifications in Mood Disorders



Abstract
:1. Introduction
2. Adenosine to Inosine (A-to-I): The Most Common Form of RNA Editing
2.1. RNA Editing in MDD
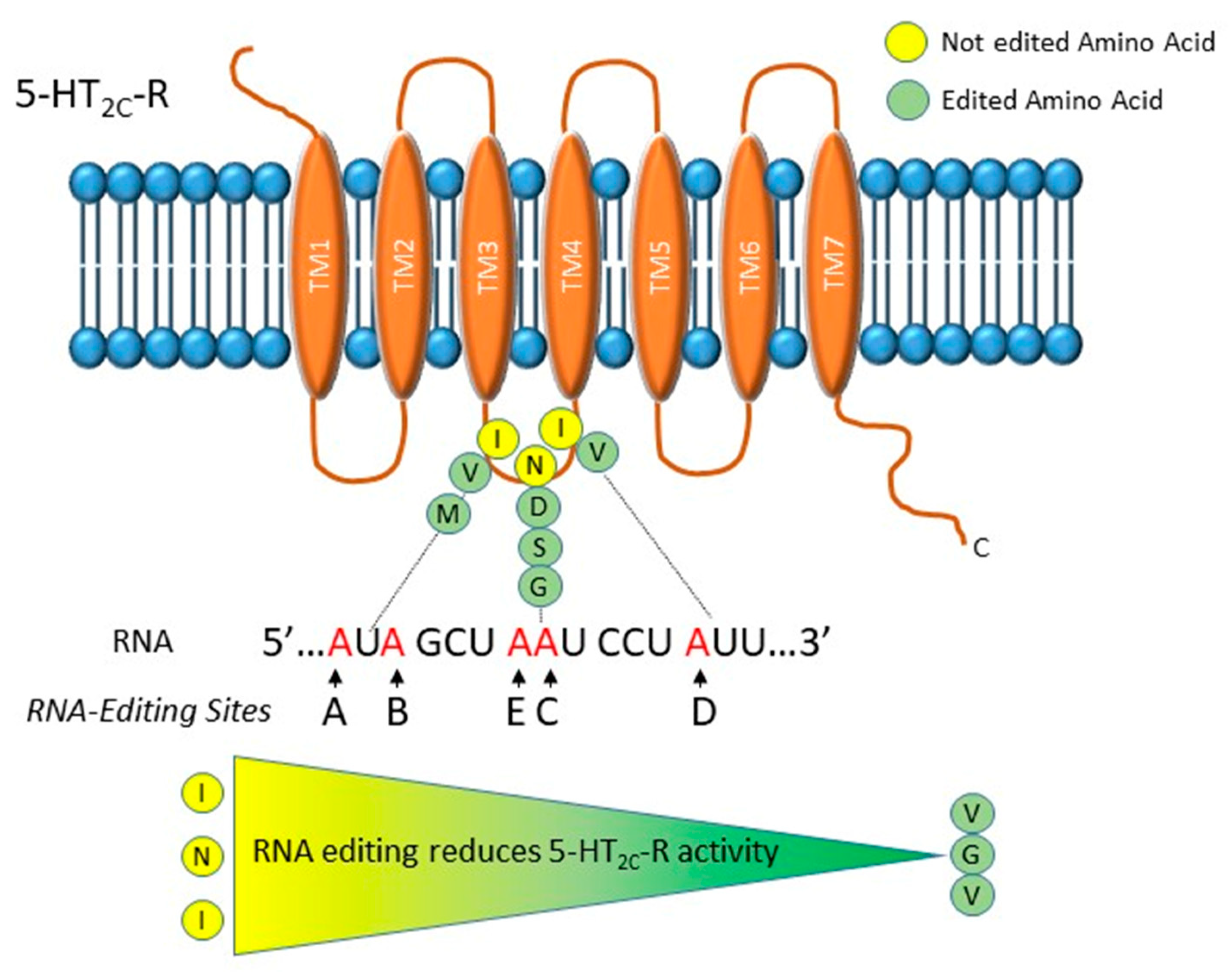
2.1.1. RNA Editing in the Serotonin Receptor 2C
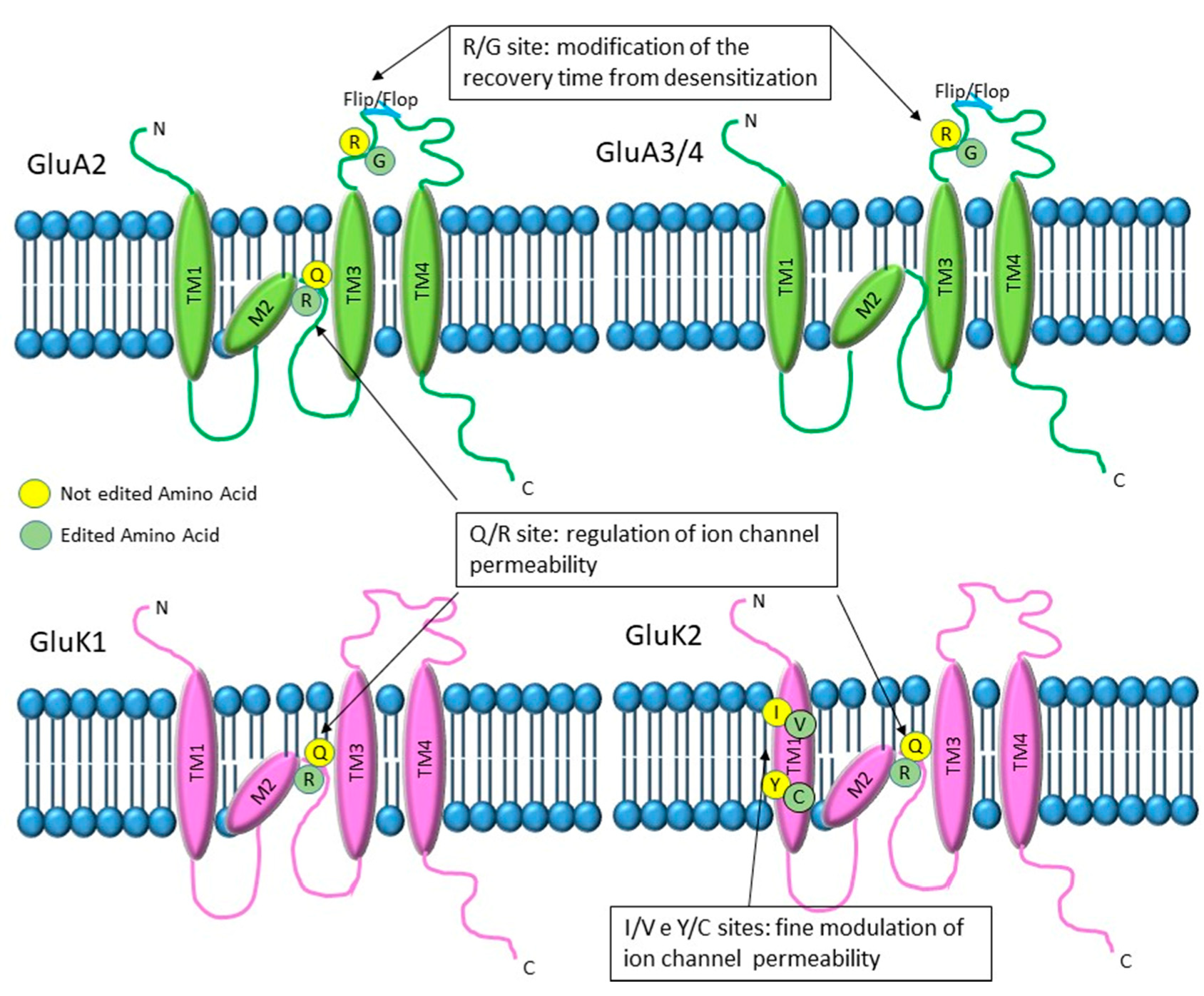
2.1.2. RNA Editing in Glutamate Receptor Transcripts
2.2. Stress Paradigm Affect RNA Editing Activity
2.3. Antidepressant Treatment Modulate RNA Editing Activity
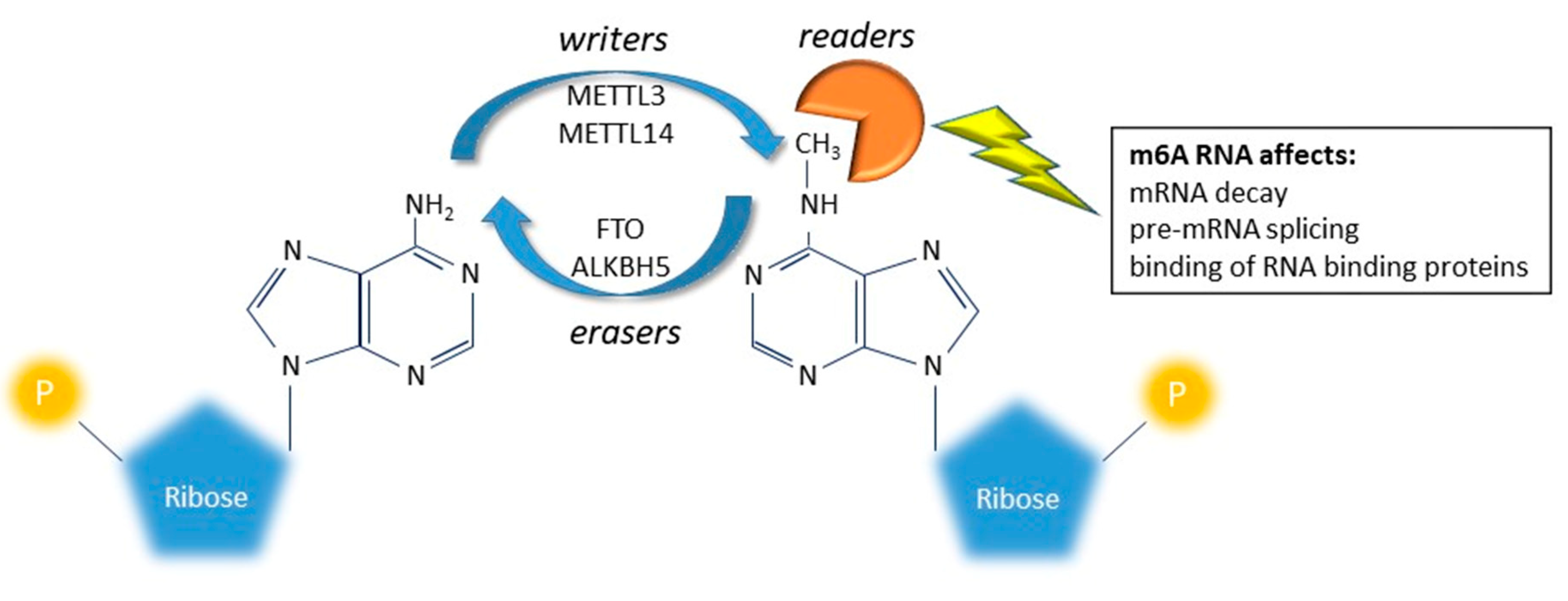
3. m6A RNA Modification
4. Conclusions and Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 354 Diseases and Injuries for 195 countries and territories, 1990-2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef] [Green Version]
- Purves, K.L.; Coleman, J.R.I.; Meier, S.M.; Rayner, C.; Davis, K.A.S.; Cheesman, R.; Bækvad-Hansen, M.; Børglum, A.D.; Wan Cho, S.; Jürgen Deckert, J.; et al. A major role for common genetic variation in anxiety disorders. Mol. Psychiatry 2019, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, S.M.; Trontti, K.; Purves, K.L.; Als, T.D.; Grove, J.; Laine, M.; Pedersen, M.G.; Bybjerg-Grauholm, J.; Bækved-Hansen, M.; Sokolowska, E.; et al. Genetic Variants Associated With Anxiety and Stress-Related Disorders: A Genome-Wide Association Study and Mouse-Model Study. JAMA Psychiatry 2019, 76, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Rosenblat, J.D.; Brietzke, E.; Pan, Z.; Lee, Y.; Cao, B.; Zuckerman, H.; Kalantarova, A.; McIntyre, R.S. Stress, epigenetics and depression: A systematic review. Neurosci. Biobehav. Rev. 2019, 102, 139–152. [Google Scholar] [CrossRef]
- Peña, C.J.; Nestler, E.J. Progress in Epigenetics of Depression. In Progress in Molecular Biology and Translational Science; Elsevier, B.V.: Amsterdam, The Netherlands, 2018; Volume 157, pp. 41–66. [Google Scholar]
- Jung, Y.; Goldman, D. Role of RNA modifications in brain and behavior. Genes Brain Behav. 2018, 17, e12444. [Google Scholar] [CrossRef] [Green Version]
- Licht, K.; Jantsch, M.F. Rapid and dynamic transcriptome regulation by RNA editing and RNA modifications. J. Cell Biol. 2016, 213, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.S.; Roundtree, I.A.; He, C. Post-transcriptional gene regulation by mRNA modifications. Nat. Rev. Mol. Cell Biol. 2017, 18, 31–42. [Google Scholar] [CrossRef]
- Gilbert, W.V.; Bell, T.A.; Schaening, C. Messenger RNA modifications: Form, distribution, and function. Science 2016, 352, 1408–1412. [Google Scholar] [CrossRef] [Green Version]
- Bajad, P.; Jantsch, M.F.; Keegan, L.; O’Connell, M. A to I editing in disease is not fake news. RNA Biol. 2017, 14, 1223–1231. [Google Scholar] [CrossRef]
- Benne, R.; Van den Burg, J.; Brakenhoff, J.P.; Sloof, P.; Van Boom, J.H.; Tromp, M.C. Major transcript of the frameshifted coxII gene from trypanosome mitochondria contains four nucleotides that are not encoded in the DNA. Cell 1986, 46, 819–826. [Google Scholar] [CrossRef]
- Daniel, C.; Lagergren, J.; Öhman, M. RNA editing of non-coding RNA and its role in gene regulation. Biochimie 2015, 117, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Nishikura, K. A-to-I editing of coding and non-coding RNAs by ADARs. Nat. Rev. Mol. Cell Biol. 2016, 17, 83–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livneh, I.; Moshitch-Moshkovitz, S.; Amariglio, N.; Rechavi, G.; Dominissini, D. The m6A epitranscriptome: Transcriptome plasticity in brain development and function. Nat. Rev. Neurosci. 2020, 21, 36–51. [Google Scholar] [CrossRef] [PubMed]
- Orlandi, C.; Barbon, A.; Barlati, S. Activity regulation of adenosine deaminases acting on RNA (ADARs). Mol. Neurobiol. 2012, 45, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Nishikura, K. Functions and regulation of RNA editing by ADAR deaminases. Annu. Rev. Biochem. 2010, 79, 321–349. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Li, X.; Qi, R.; Billiar, T. RNA Editing, ADAR1, and the Innate Immune Response. Genes (Basel) 2017, 8, 41. [Google Scholar] [CrossRef] [Green Version]
- Filippini, A.; Bonini, D.; Giacopuzzi, E.; La Via, L.; Gangemi, F.; Colombi, M.; Barbon, A. Differential Enzymatic Activity of Rat ADAR2 Splicing Variants Is Due to Altered Capability to Interact with RNA in the Deaminase Domain. Genes (Basel) 2018, 9, 79. [Google Scholar] [CrossRef] [Green Version]
- Gerber, A.; O’Connell, M.A.; Keller, W. Two forms of human double-stranded RNA-specific editase 1 (hRED1) generated by the insertion of an Alu cassette. RNA 1997, 3, 453–463. [Google Scholar]
- Daniel, C.; Behm, M.; Öhman, M. The role of Alu elements in the cis-regulation of RNA processing. Cell. Mol. Life Sci. 2015, 72, 4063–4076. [Google Scholar] [CrossRef]
- Tan, M.H.; Li, Q.; Shanmugam, R.; Piskol, R.; Kohler, J.; Young, A.N.; Liu, K.I.; Zhang, R.; Ramaswami, G.; Ariyoshi, K.; et al. Dynamic landscape and regulation of RNA editing in mammals. Nature 2017, 550, 249–254. [Google Scholar] [CrossRef]
- Picardi, E.; D’Erchia, A.M.; Lo Giudice, C.; Pesole, G. REDIportal: A comprehensive database of A-to-I RNA editing events in humans. Nucleic Acids Res. 2017, 45, D750–D757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramaswami, G.; Li, J.B. RADAR: A rigorously annotated database of A-to-I RNA editing. Nucleic Acids Res. 2014, 42, D109–D113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behm, M.; Öhman, M. RNA Editing: A Contributor to Neuronal Dynamics in the Mammalian Brain. Trends Genet. 2016, 32, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, J.J.C.; Seeburg, P.H. A-to-I RNA editing: Effects on proteins key to neural excitability. Neuron 2012, 74, 432–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlandi, C.; La Via, L.; Bonini, D.; Mora, C.; Russo, I.; Barbon, A.; Barlati, S. AMPA receptor regulation at the mRNA and protein level in rat primary cortical cultures. PLoS ONE 2011, 6, e25350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahlstedt, H.; Daniel, C.; Ensterö, M.; Ohman, M. Large-scale mRNA sequencing determines global regulation of RNA editing during brain development. Genome Res. 2009, 19, 978–986. [Google Scholar] [CrossRef] [Green Version]
- Gaisler-Salomon, I.; Kravitz, E.; Feiler, Y.; Safran, M.; Biegon, A.; Amariglio, N.; Rechavi, G. Hippocampus-specific deficiency in RNA editing of GluA2 in Alzheimer’s disease. Neurobiol. Aging 2014, 35, 1785–1791. [Google Scholar] [CrossRef]
- Gardner, O.K.; Wang, L.; Van Booven, D.; Whitehead, P.L.; Hamilton-Nelson, K.L.; Adams, L.D.; Starks, T.D.; Hofmann, N.K.; Vance, J.M.; Cuccaro, M.L.; et al. RNA editing alterations in a multi-ethnic Alzheimer disease cohort converge on immune and endocytic molecular pathways. Hum. Mol. Genet. 2019, 28, 3053–3061. [Google Scholar] [CrossRef]
- Moore, S.; Alsop, E.; Lorenzini, I.; Starr, A.; Rabichow, B.E.; Mendez, E.; Levy, J.L.; Burciu, C.; Reiman, R.; Chew, J.; et al. ADAR2 mislocalization and widespread RNA editing aberrations in C9orf72-mediated ALS/FTD. Acta Neuropathol. 2019, 138, 49–65. [Google Scholar] [CrossRef]
- Breen, M.S.; Dobbyn, A.; Li, Q.; Roussos, P.; Hoffman, G.E.; Stahl, E.; Chess, A.; Sklar, P.; Li, J.B.; Devlin, B.; et al. Global landscape and genetic regulation of RNA editing in cortical samples from individuals with schizophrenia. Nat. Neurosci. 2019, 22, 1402–1412. [Google Scholar] [CrossRef]
- Tran, S.S.; Jun, H.-I.; Bahn, J.H.; Azghadi, A.; Ramaswami, G.; Van Nostrand, E.L.; Nguyen, T.B.; Hsiao, Y.-H.E.; Lee, C.; Pratt, G.A.; et al. Widespread RNA editing dysregulation in brains from autistic individuals. Nat. Neurosci. 2019, 22, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Eran, A.; Li, J.B.; Vatalaro, K.; McCarthy, J.; Rahimov, F.; Collins, C.; Markianos, K.; Margulies, D.M.; Brown, E.N.; Calvo, S.E.; et al. Comparative RNA editing in autistic and neurotypical cerebella. Mol. Psychiatry 2013, 18, 1041–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanacora, G.; Zarate, C.A.; Krystal, J.H.; Manji, H.K. Targeting the glutamatergic system to develop novel, improved therapeutics for mood disorders. Nat. Rev. Drug Discov. 2008, 7, 426–437. [Google Scholar] [CrossRef] [Green Version]
- Sanacora, G.; Treccani, G.; Popoli, M. Towards a glutamate hypothesis of depression: An emerging frontier of neuropsychopharmacology for mood disorders. Neuropharmacology 2012, 62, 63–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoyer, D.; Clarke, D.E.; Fozard, J.R.; Hartig, P.R.; Martin, G.R.; Mylecharane, E.J.; Saxena, P.R.; Humphrey, P.P. International Union of Pharmacology classification of receptors for 5-hydroxytryptamine (Serotonin). Pharmacol. Rev. 1994, 46, 157–203. [Google Scholar] [PubMed]
- Sanders-Bush, E.; Canton, H. Serotonin Receptors: Signal Transduction Pathways. In Psychopharmacology: The Fourth Generation of Progress; Bloom, F.E., Kupfer, D.J., Eds.; Raven Press: New York, NY, USA, 1995; pp. 431–441. [Google Scholar]
- Burns, C.M.; Chu, H.; Rueter, S.M.; Hutchinson, L.K.; Canton, H.; Sanders-Bush, E.; Emeson, R.B. Regulation of serotonin-2C receptor G-protein coupling by RNA editing. Nature 1997, 387, 303–308. [Google Scholar] [CrossRef]
- Vitali, P.; Basyuk, E.; Le Meur, E.; Bertrand, E.; Muscatelli, F.; Cavaillé, J.; Huttenhofer, A. ADAR2-mediated editing of RNA substrates in the nucleolus is inhibited by C/D small nucleolar RNAs. J. Cell Biol. 2005, 169, 745–753. [Google Scholar] [CrossRef]
- Niswender, C.M.; Herrick-Davis, K.; Dilley, G.E.; Meltzer, H.Y.; Overholser, J.C.; Stockmeier, C.A.; Emeson, R.B.; Sanders-Bush, E. RNA editing of the human serotonin 5-HT2C receptor. alterations in suicide and implications for serotonergic pharmacotherapy. Neuropsychopharmacology 2001, 24, 478–491. [Google Scholar] [CrossRef]
- Iwamoto, K.; Kato, T. RNA editing of serotonin 2C receptor in human postmortem brains of major mental disorders. Neurosci. Lett. 2003, 346, 169–172. [Google Scholar] [CrossRef]
- Gurevich, I.; Tamir, H.; Arango, V.; Dwork, A.J.; Mann, J.J.; Schmauss, C. Altered editing of serotonin 2C receptor pre-mRNA in the prefrontal cortex of depressed suicide victims. Neuron 2002, 34, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Urban, D.J.; Blashka, J.; McPheeters, M.T.; Kroeze, W.K.; Mieczkowski, P.; Overholser, J.C.; Jurjus, G.J.; Dieter, L.; Mahajan, G.J.; et al. Quantitative analysis of focused a-to-I RNA editing sites by ultra-high-throughput sequencing in psychiatric disorders. PLoS ONE 2012, 7, e43227. [Google Scholar] [CrossRef] [PubMed]
- Lyddon, R.; Dwork, A.J.; Keddache, M.; Siever, L.J.; Dracheva, S. Serotonin 2c receptor RNA editing in major depression and suicide. World J. Biol. Psychiatry 2013, 14, 590–601. [Google Scholar] [CrossRef] [Green Version]
- Di Narzo, A.F.; Kozlenkov, A.; Roussos, P.; Hao, K.; Hurd, Y.; Lewis, D.A.; Sibille, E.; Siever, L.J.; Koonin, E.; Dracheva, S. A unique gene expression signature associated with serotonin 2C receptor RNA editing in the prefrontal cortex and altered in suicide. Hum. Mol. Genet. 2014, 23, 4801–4813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissmann, D.; van der Laan, S.; Underwood, M.D.; Salvetat, N.; Cavarec, L.; Vincent, L.; Molina, F.; Mann, J.J.; Arango, V.; Pujol, J.F. Region-specific alterations of A-to-I RNA editing of serotonin 2c receptor in the cortex of suicides with major depression. Transl. Psychiatry 2016, 6, e878. [Google Scholar] [CrossRef] [PubMed]
- Chimienti, F.; Cavarec, L.; Vincent, L.; Salvetat, N.; Arango, V.; Underwood, M.D.; Mann, J.J.; Pujol, J.-F.; Weissmann, D. Brain region-specific alterations of RNA editing in PDE8A mRNA in suicide decedents. Transl. Psychiatry 2019, 9, 91. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, R.T.; Emeson, R.B. Quantitative analysis of 5HT(2C) receptor RNA editing patterns in psychiatric disorders. Neurobiol. Dis. 2012, 45, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Karanović, J.; Šviković, S.; Pantović, M.; Durica, S.; Brajušković, G.; Damjanović, A.; Jovanović, V.; Ivković, M.; Romac, S.; Savić Pavićević, D. Joint effect of ADARB1 gene, HTR2C gene and stressful life events on suicide attempt risk in patients with major psychiatric disorders. World J. Biol. Psychiatry 2015, 16, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Karanović, J.; Ivković, M.; Jovanović, V.M.; Šviković, S.; Pantović-Stefanović, M.; Brkušanin, M.; Damjanović, A.; Brajušković, G.; Savić-Pavićević, D. Effect of childhood general traumas on suicide attempt depends on TPH2 and ADARB1 variants in psychiatric patients. J. Neural Transm. 2017, 124, 621–629. [Google Scholar] [CrossRef]
- Barbon, A.; Barlati, S. Glutamate receptor RNA editing in health and disease. Biochemistry (Mosc.) 2011, 76, 882–889. [Google Scholar] [CrossRef]
- Filippini, A.; Bonini, D.; La Via, L.; Barbon, A. The Good and the Bad of Glutamate Receptor RNA Editing. Mol. Neurobiol. 2017, 54, 6795–6805. [Google Scholar] [CrossRef] [Green Version]
- Kubota-Sakashita, M.; Iwamoto, K.; Bundo, M.; Kato, T. A role of ADAR2 and RNA editing of glutamate receptors in mood disorders and schizophrenia. Mol. Brain 2014, 7, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McEwen, B.S.; Akil, H. Revisiting the Stress Concept: Implications for Affective Disorders. J. Neurosci. 2020, 40, 12–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmauss, C. Serotonin 2C receptors: Suicide, serotonin, and runaway RNA editing. Neuroscientist 2003, 9, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Englander, M.T.; Dulawa, S.C.; Bhansali, P.; Schmauss, C. How stress and fluoxetine modulate serotonin 2C receptor pre-mRNA editing. J. Neurosci. 2005, 25, 648–651. [Google Scholar] [CrossRef] [Green Version]
- Iwamoto, K.; Nakatani, N.; Bundo, M.; Yoshikawa, T.; Kato, T. Altered RNA editing of serotonin 2C receptor in a rat model of depression. Neurosci. Res. 2005, 53, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Stasko, M.; Costa, A.C.; Davisson, M.T.; Gardiner, K.J. Editing of the serotonin 2C receptor pre-mRNA: Effects of the Morris Water Maze. Gene 2007, 391, 186–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhansali, P.; Dunning, J.; Singer, S.E.; David, L.; Schmauss, C. Early life stress alters adult serotonin 2C receptor pre-mRNA editing and expression of the α subunit of the heterotrimeric G-protein G q. J. Neurosci. 2007, 27, 1467–1473. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, Y.; Grimberg, A.; Teegarden, S.; Mombereau, C.; Liu, S.; Bale, T.L.; Blendy, J.A.; Nishikura, K. Dysregulated editing of serotonin 2C receptor mRNAs results in energy dissipation and loss of fat mass. J. Neurosci. 2008, 28, 12834–12844. [Google Scholar] [CrossRef] [Green Version]
- Mombereau, C.; Kawahara, Y.; Gundersen, B.B.; Nishikura, K.; Blendy, J.A. Functional relevance of serotonin 2C receptor mRNA editing in antidepressant- and anxiety-like behaviors. Neuropharmacology 2010, 59, 468–473. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.B.P.; Ramond, F.; Farrington, D.T.; Aguiar, A.S.; Chevarin, C.; Berthiau, A.-S.; Caussanel, S.; Lanfumey, L.; Herrick-Davis, K.; Hamon, M.; et al. RNA splicing and editing modulation of 5-HT(2C) receptor function: Relevance to anxiety and aggression in VGV mice. Mol. Psychiatry 2013, 18, 656–665. [Google Scholar] [CrossRef]
- Bombail, V.; Qing, W.; Chapman, K.E.; Holmes, M.C. Prevention of 5-hydroxytryptamine2C receptor RNA editing and alternate splicing in C57BL/6 mice activates the hypothalamic-pituitary-adrenal axis and alters mood. Eur. J. Neurosci. 2014, 40, 3663–3673. [Google Scholar] [CrossRef] [Green Version]
- Règue, M.; Poilbout, C.; Martin, V.; Franc, B.; Lanfumey, L.; Mongeau, R. Increased 5-HT2C receptor editing predisposes to PTSD-like behaviors and alters BDNF and cytokines signaling. Transl. Psychiatry 2019, 9, 100. [Google Scholar] [CrossRef] [Green Version]
- Brande-Eilat, N.; Golumbic, Y.N.; Zaidan, H.; Gaisler-Salomon, I. Acquisition of conditioned fear is followed by region-specific changes in RNA editing of glutamate receptors. Stress 2015, 18, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Bonini, D.; Mora, C.; Tornese, P.; Sala, N.; Filippini, A.; La Via, L.; Milanese, M.; Calza, S.; Bonanno, G.; Racagni, G.; et al. Acute Footshock Stress Induces Time-Dependent Modifications of AMPA/NMDA Protein Expression and AMPA Phosphorylation. Neural Plast. 2016, 2016, 7267865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elhussiny, E.A.M.; Carini, G.; Mingardi, J.; Tornese, P.; Sala, N.; Bono, F.; Fiorentini, C.; La Via, L.; Popoli, M.; Musazzi, L.; et al. Modulation by chronic stress and ketamine of ionotropic AMPA/NMDA and metabotropic glutamate receptors in the rat hippocampus. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 104, 110033. [Google Scholar] [CrossRef]
- Dick, A.L.W.; Khermesh, K.; Paul, E.; Stamp, F.; Levanon, E.Y.; Chen, A. Adenosine-to-Inosine RNA Editing Within Corticolimbic Brain Regions Is Regulated in Response to Chronic Social Defeat Stress in Mice. Front. Psychiatry 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gal-Mark, N.; Shallev, L.; Sweetat, S.; Barak, M.; Billy Li, J.; Levanon, E.Y.; Eisenberg, E.; Behar, O. Abnormalities in A-to-I RNA editing patterns in CNS injuries correlate with dynamic changes in cell type composition. Sci. Rep. 2017, 7, 43421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picardi, E.; Horner, D.S.; Pesole, G. Single-cell transcriptomics reveals specific RNA editing signatures in the human brain. RNA 2017, 23, 860–865. [Google Scholar] [CrossRef] [Green Version]
- Zaidan, H.; Ramaswami, G.; Golumbic, Y.N.; Sher, N.; Malik, A.; Barak, M.; Galiani, D.; Dekel, N.; Li, J.B.; Gaisler-Salomon, I. A-to-I RNA editing in the rat brain is age-dependent, region-specific and sensitive to environmental stress across generations. BMC Genom. 2018, 19, 28. [Google Scholar] [CrossRef] [Green Version]
- Abbas, A.I.; Urban, D.J.; Jensen, N.H.; Farrell, M.S.; Kroeze, W.K.; Mieczkowski, P.; Wang, Z.; Roth, B.L. Assessing serotonin receptor mRNA editing frequency by a novel ultra high-throughput sequencing method. Nucleic Acids Res. 2010, 38, e118. [Google Scholar] [CrossRef]
- Barbon, A.; Orlandi, C.; La Via, L.; Caracciolo, L.; Tardito, D.; Musazzi, L.; Mallei, A.; Gennarelli, M.; Racagni, G.; Popoli, M.; et al. Antidepressant treatments change 5-HT2C receptor mRNA expression in rat prefrontal/frontal cortex and hippocampus. Neuropsychobiology 2011, 63, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Aoki, M.; Watanabe, Y.; Yoshimoto, K.; Tsujimura, A.; Yamamoto, T.; Kanamura, N.; Tanaka, M. Involvement of serotonin 2C receptor RNA editing in accumbal neuropeptide Y expression and behavioural despair. Eur. J. Neurosci. 2016, 43, 1219–1228. [Google Scholar] [CrossRef] [PubMed]
- Zaidan, H.; Ramaswami, G.; Barak, M.; Li, J.B.; Gaisler-Salomon, I. Pre-reproductive stress and fluoxetine treatment in rats affect offspring A-to-I RNA editing, gene expression and social behavior. Environ. Epigenetics 2018, 4, dvy021. [Google Scholar] [CrossRef] [PubMed]
- Barbon, A.; Popoli, M.; La Via, L.; Moraschi, S.; Vallini, I.; Tardito, D.; Tiraboschi, E.; Musazzi, L.; Giambelli, R.; Gennarelli, M.; et al. Regulation of editing and expression of glutamate α-amino-propionic-acid (AMPA)/kainate receptors by antidepressant drugs. Biol. Psychiatry 2006, 59, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Barbon, A.; Caracciolo, L.; Orlandi, C.; Musazzi, L.; Mallei, A.; La Via, L.; Bonini, D.; Mora, C.; Tardito, D.; Gennarelli, M.; et al. Chronic antidepressant treatments induce a time-dependent up-regulation of AMPA receptor subunit protein levels. Neurochem. Int. 2011, 59, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Sawada, J.; Yamashita, T.; Aizawa, H.; Aburakawa, Y.; Hasebe, N.; Kwak, S. Effects of antidepressants on GluR2 Q/R site-RNA editing in modified HeLa cell line. Neurosci. Res. 2009, 64, 251–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Zhang, S.; Zhang, H.; Hertz, L.; Peng, L. Fluoxetine affects GluK2 editing, glutamate-evoked Ca(2+) influx and extracellular signal-regulated kinase phosphorylation in mouse astrocytes. J. Psychiatry Neurosci. 2011, 36, 322–338. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Dong, L.; Wang, B.; Cai, L.; Jiang, N.; Peng, L. Cell type-specific gene expression and editing responses to chronic fluoxetine treatment in the in vivo mouse brain and their relevance for stress-induced anhedonia. Neurochem. Res. 2012, 37, 2480–2495. [Google Scholar] [CrossRef]
- Koranda, J.L.; Dore, L.; Shi, H.; Patel, M.J.; Vaasjo, L.O.; Rao, M.N.; Chen, K.; Lu, Z.; Yi, Y.; Chi, W.; et al. Mettl14 Is Essential for Epitranscriptomic Regulation of Striatal Function and Learning. Neuron 2018, 99, 283–292.e5. [Google Scholar] [CrossRef] [Green Version]
- Merkurjev, D.; Hong, W.-T.; Iida, K.; Oomoto, I.; Goldie, B.J.; Yamaguti, H.; Ohara, T.; Kawaguchi, S.-Y.; Hirano, T.; Martin, K.C.; et al. Synaptic N6-methyladenosine (m6A) epitranscriptome reveals functional partitioning of localized transcripts. Nat. Neurosci. 2018, 21, 1004–1014. [Google Scholar] [CrossRef]
- Weng, Y.-L.; Wang, X.; An, R.; Cassin, J.; Vissers, C.; Liu, Y.; Liu, Y.; Xu, T.; Wang, X.; Wong, S.Z.H.; et al. Epitranscriptomic m6A Regulation of Axon Regeneration in the Adult Mammalian Nervous System. Neuron 2018, 97, 313–325.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, K.-J.; Ringeling, F.R.; Vissers, C.; Jacob, F.; Pokrass, M.; Jimenez-Cyrus, D.; Su, Y.; Kim, N.-S.; Zhu, Y.; Zheng, L.; et al. Temporal Control of Mammalian Cortical Neurogenesis by m6A Methylation. Cell 2017, 171, 877–889.e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Yang, X.; Qi, Z.; Sang, Y.; Liu, Y.; Xu, B.; Liu, W.; Xu, Z.; Deng, Y. The role of mRNA m6A methylation in the nervous system. Cell Biosci. 2019, 9, 66. [Google Scholar] [CrossRef] [PubMed]
- Samaan, Z.; Anand, S.S.; Anand, S.; Zhang, X.; Desai, D.; Rivera, M.; Pare, G.; Thabane, L.; Xie, C.; Gerstein, H.; et al. The protective effect of the obesity-associated rs9939609 A variant in fat mass- and obesity-associated gene on depression. Mol. Psychiatry 2013, 18, 1281–1286. [Google Scholar] [CrossRef]
- Milaneschi, Y.; Lamers, F.; Mbarek, H.; Hottenga, J.-J.; Boomsma, D.I.; Penninx, B.W.J.H. The effect of FTO rs9939609 on major depression differs across MDD subtypes. Mol. Psychiatry 2014, 19, 960–962. [Google Scholar] [CrossRef] [Green Version]
- Du, T.; Rao, S.; Wu, L.; Ye, N.; Liu, Z.; Hu, H.; Xiu, J.; Shen, Y.; Xu, Q. An association study of the m6A genes with major depressive disorder in Chinese Han population. J. Affect. Disord. 2015, 183, 279–286. [Google Scholar] [CrossRef]
- Engel, M.; Eggert, C.; Kaplick, P.M.; Eder, M.; Röh, S.; Tietze, L.; Namendorf, C.; Arloth, J.; Weber, P.; Rex-Haffner, M.; et al. The Role of m6A/m-RNA Methylation in Stress Response Regulation. Neuron 2018, 99, 389–403.e9. [Google Scholar] [CrossRef]
- Huang, R.; Zhang, Y.; Bai, Y.; Han, B.; Ju, M.; Chen, B.; Yang, L.; Wang, Y.; Zhang, H.; Zhang, H.; et al. N6-Methyladenosine Modification of Fatty Acid Amide Hydrolase Messenger RNA in Circular RNA STAG1-Regulated Astrocyte Dysfunction and Depressive-like Behaviors. Biol. Psychiatry 2020. [Google Scholar] [CrossRef] [Green Version]
- Yoon, K.-J.; Ming, G.-L.; Song, H. Epitranscriptomes in the Adult Mammalian Brain: Dynamic Changes Regulate Behavior. Neuron 2018, 99, 243–245. [Google Scholar] [CrossRef] [Green Version]
- De Deurwaerdère, P.; Navailles, S.; Berg, K.A.; Clarke, W.P.; Spampinato, U. Constitutive activity of the serotonin2C receptor inhibits in vivo dopamine release in the rat striatum and nucleus accumbens. J. Neurosci. 2004, 24, 3235–3241. [Google Scholar] [CrossRef]
- Heshmati, M.; Christoffel, D.J.; LeClair, K.; Cathomas, F.; Golden, S.A.; Aleyasin, H.; Turecki, G.; Friedman, A.K.; Han, M.-H.; Menard, C.; et al. Depression and social defeat stress are associated with inhibitory synaptic changes in the nucleus accumbens. J. Neurosci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Giacopuzzi, E.; Gennarelli, M.; Sacco, C.; Filippini, A.; Mingardi, J.; Magri, C.; Barbon, A. Genome-wide analysis of consistently RNA edited sites in human blood reveals interactions with mRNA processing genes and suggests correlations with cell types and biological variables. BMC Genom. 2018, 19, 963. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Disease | Brain Area | Single Site Analysis | Edited Isoform | Reference |
|---|---|---|---|---|
| MDD | FC | not altered (sites A, C, D) | nd | [40] |
| Suicide victims (SCZ and MDD) | FC | ↑ A, = C, ↑ D (trend) | nd | [40] |
| MDD | FC | = A, ↑ D (trend) | nd | [41] |
| Suicide victims (MDD) | FC | = A, = B,↑ E, ↓ D, ↑ C (trend) | ↑ VGI, ↓ VNI | [42] |
| MDD | Cortex | ↓ E, ↓ D, ↓ C (all trend) | not altered | [43] |
| Suicide victims (MDD) | PFC | nd | ↑VSV | [45] |
| Suicide victims (MDD) | ACC | nd | ↑VNI ↓INV↑ VDV | [46] |
| Suicide victims (MDD) | FC | nd | ↑ VNI | [46] |
| Animal Model | Brain Region | Treatment | Effect | Reference |
|---|---|---|---|---|
| Mouse C57BL/6 | Striatum and HI | Chronic fluoxetine | ↑A and B | [72] |
| Mouse C57BL/6 | Cortex | Chronic fluoxetine | No Effects | [56,72] |
| Mouse BALB/c | FC | Chronic fluoxetine | ↑A, B, C, D = E | [56] |
| Rat Sprague-Dawley | FC | Reboxetine | ↓ D, E | [73] |
| Rat Sprague-Dawley | FC | Fluoxetine | Non effects | [73] |
| Mouse C57BL/6 | Striatum and HI | Tricyclic, amitriptyline, SB-206553 | ↑A and B | [72] |
| Rat Sprague-Dawley | Amygdala of the offspring at birth | Fluoxetine | ↓ A;B and C | [75] |
| Rat Sprague-Dawley | PFC | Fluoxetine | = GluR2 Q/R, ↑ GluR2 R/G (flip isoform only); GluR3R/G, GluR4 R/G, GluR5 Q/R, GluR6 I/V, GluR6 Y/C (no alteration) ↓GluR6 Q/R | [76] |
| Rat Sprague-Dawley | PFC | Desipramine | ↑ GluR2 R/G (flop isoform only), ↑ GluR4 R/G GluR3R/G, GluR5 Q/R, GluR6 I/V, GluR6 Y/C, GluR6 Q/R (no alteration) | [76] |
| Rat Sprague-Dawley | PFC | Reboxetine | = GluR2 Q/R, ↑ GluR2 R/G (flip isoform only), ↑ GluR4 R/G GluR3R/G, GluR5 Q/R, GluR6 I/V, GluR6 Y/C (no alteration) ↓GluR6 Q/R | [76] |
| Rat Sprague-Dawley | HI | Fluoxetine | GluR2 Q/R, GluR2 R/G, GluR3R/G, GluR4 R/G, GluR5 Q/R, GluR6 I/V, GluR6 Y/C (no alteration) ↓GluR6 Q/R | [76] |
| Rat Sprague-Dawley | HI | Desipramine | ↓ GluR3R/G (flop isoform only) GluR2 Q/R, GluR2 R/G, GluR3R/G, GluR4 R/G, GluR5 Q/R, GluR6 I/V, GluR6 Y/C, GluR6 Q/R (no alteration) | [76] |
| Rat Sprague-Dawley | HI | Reboxetine | ↓ GluR3R/G (flip isoform only) GluR2 Q/R, GluR2 R/G, GluR4 R/G, GluR5 Q/R, GluR6 I/V, GluR6 Y/C (no alteration) ↓GluR6 Q/R | [76] |
| Rat Sprague-Dawley | P/FC | Fluoxetine | ↓ GluA2 R/G (flop isoform) observed after 3 weeks of treatment and washout; ↓GluA3 R/G (flip variant) observed after 3 weeks of treatment ↑ GluA4 R/G (flip isoform) observed after 3 weeks of treatment | [77] |
| Rat Sprague-Dawley | P/FC | Reboxetine | ↓ GluA2 R/G (flop isoform) observed after 1 and 3 weeks of treatment and washout; ↑ GluA4 R/G (flip isoform) observed at all point tested | [77] |
| Rat Sprague-Dawley | HI | Fluoxetine/Reboxetine | ↑GluA2 R/G (flip and flop isoforms) observed after 2 and 3 weeks of treatment GluA3 and GluA4 R/G sites were unchanged | [77] |
| TetHeLaG2m cells | Fluvoxamine, fluoxetine, paroxetine, milnacipran, reboxetine, amitriptyline, desipramine, imipramine | ↑ GluR2 Q/R after incubation with each antidepressant except reboxentine | [78] | |
| Male adult CD-1 | cultured astrocytes intact brain | fluoxetine | ↑ GluK2 I/V, GluK2 Y/C, GluK2 Q/R | [79] |
| Male and female FVB/NTg(GFAP-GFP)14Mes/J B6.Cg-Tg(Thy1-YFPH)2Jrs/J mice | cultured astrocytes | fluoxetine | ↑ GluK2 I/V, GluK2 Y/C, GluK2 Q/R | [80] |
| Male and female FVB/NTg(GFAP-GFP)14Mes/J B6.Cg-Tg(Thy1-YFPH)2Jrs/J | cultured neurons | fluoxetine | No alteration in any GluK2 editing sites | [80] |
| Rat Sprague-Dawley | HI | Ketamine | No alteration in any AMPA receptor editing sites | [67] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbon, A.; Magri, C. RNA Editing and Modifications in Mood Disorders. Genes 2020, 11, 872. https://doi.org/10.3390/genes11080872
Barbon A, Magri C. RNA Editing and Modifications in Mood Disorders. Genes. 2020; 11(8):872. https://doi.org/10.3390/genes11080872
Chicago/Turabian StyleBarbon, Alessandro, and Chiara Magri. 2020. "RNA Editing and Modifications in Mood Disorders" Genes 11, no. 8: 872. https://doi.org/10.3390/genes11080872
APA StyleBarbon, A., & Magri, C. (2020). RNA Editing and Modifications in Mood Disorders. Genes, 11(8), 872. https://doi.org/10.3390/genes11080872