Development of Chromosome Segment Substitution Lines (CSSLs) Derived from Guangxi Wild Rice (Oryza rufipogon Griff.) under Rice (Oryza sativa L.) Background and the Identification of QTLs for Plant Architecture, Agronomic Traits and Cold Tolerance


 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Phenotyping
2.3. Evaluation of Cold Tolerance at Seedling Stage
2.4. Construction of CSSLs and Genome Sequencing and Development of InDel Markers
2.5. DNA Isolation and PCR Amplification
2.6. QTL Mapping and Data Analysis
- (1)
- Additive effect (a) = phenotypic value of CSSL (CSSL) − phenotypic value of 93-11(93-11);
- (2)
- Dominant effect (d) = 93-11 × CSSL − 93-11.
3. Results
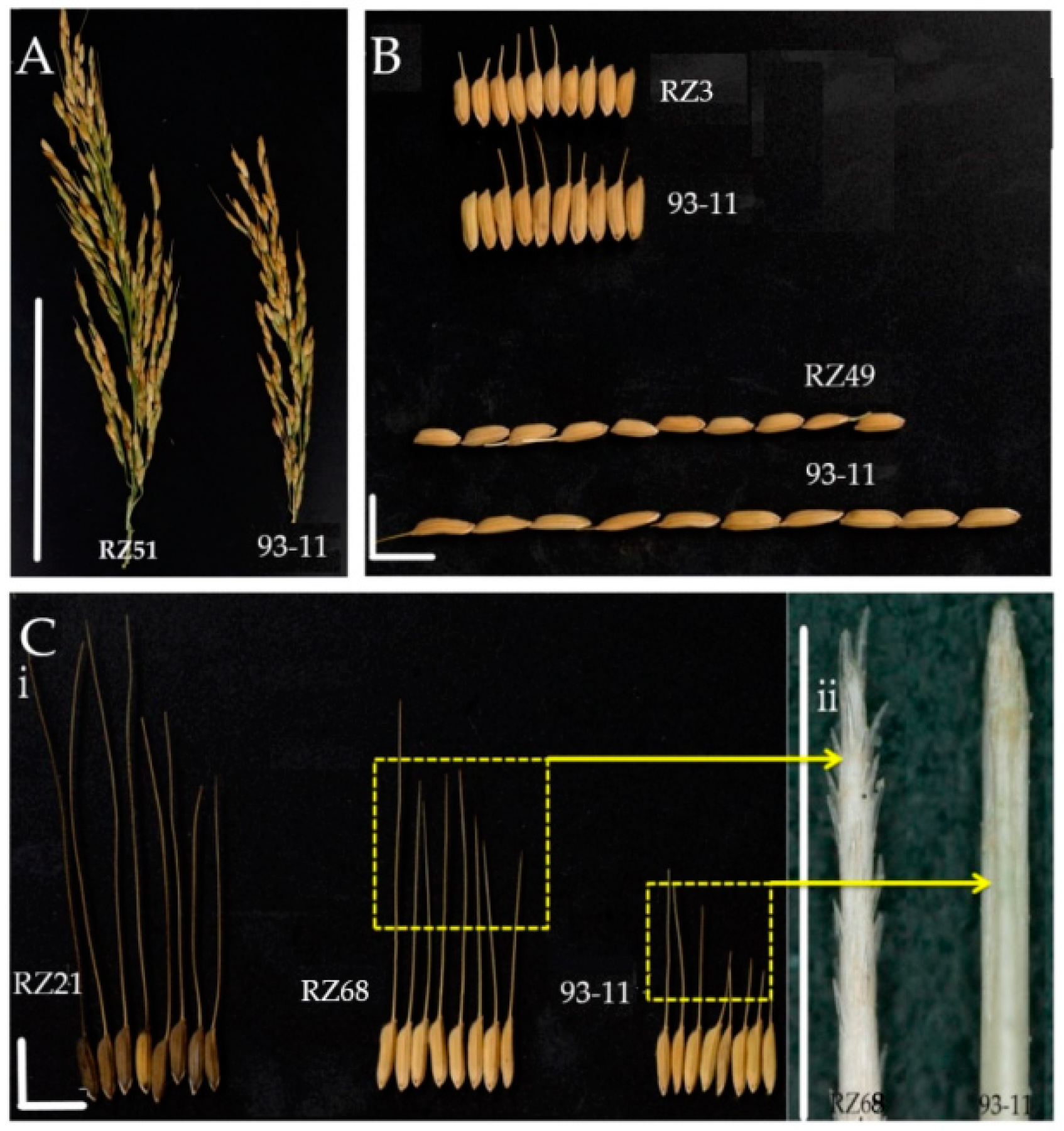
3.1. Phenotypic Variation of Plant Architecture, Agronomic Traits and Cold Tolerance in DP30-CSSLs
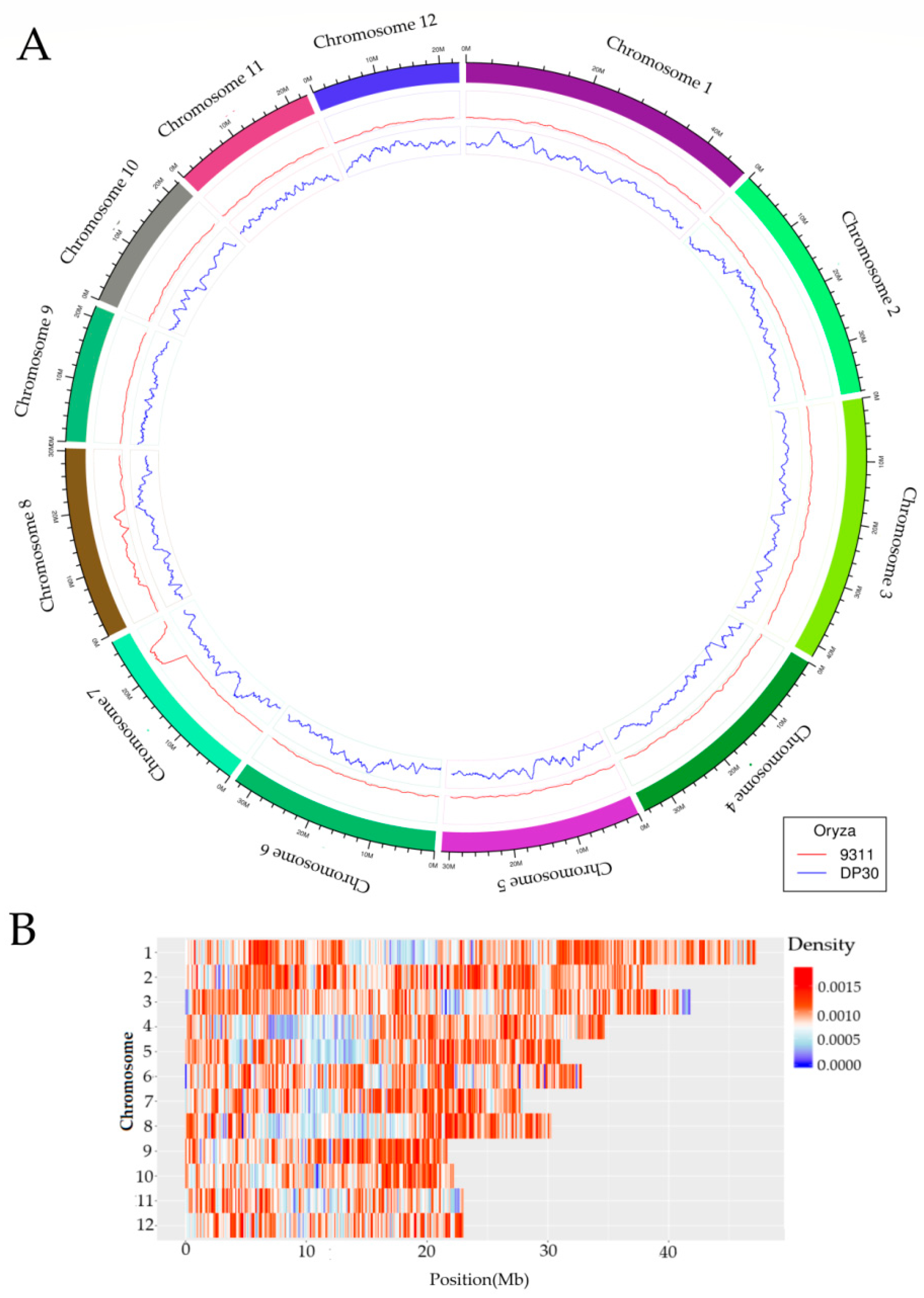
3.2. Genome Re-Sequencing and Selection of InDel Markers
3.3. Development and Substitution Segment Analysis of DP30-CSSLs
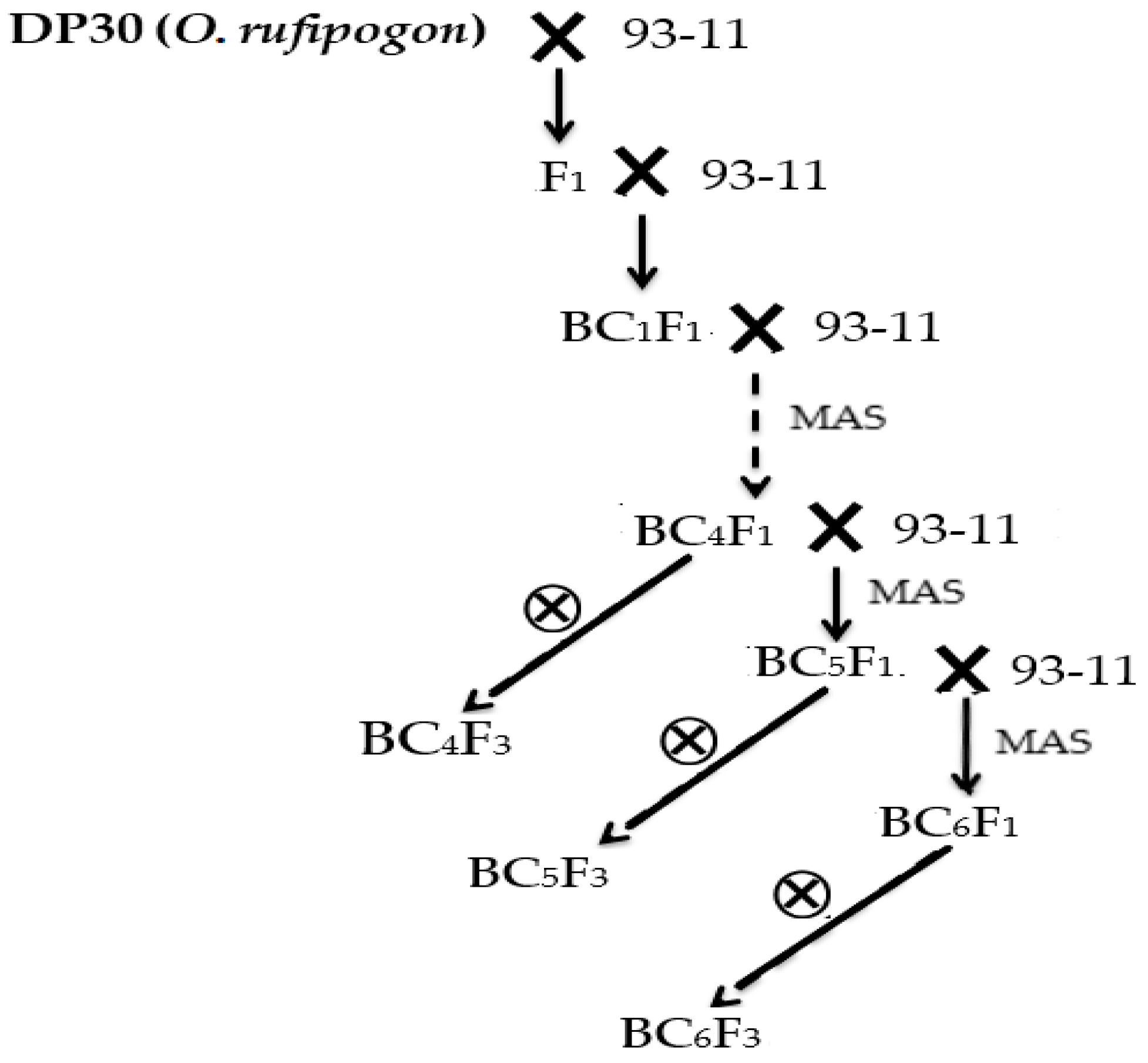
3.3.1. Development of the DP30-CSSLs
3.3.2. Substitution Segments of DP30-CSSLs
3.4. Wild rice QTLs in the DP30-CSSLs
3.4.1. Tiller Angle (TA) and Heading Date (HD)
3.4.2. Plant Height (PH)
3.4.3. Number of Grains per Panicle (NGPP) and 100 Grain Weight (GWT)
3.4.4. Grain Length (GL), Grain Width (GW) and Grain Length to Width Ratio (GLWR)
3.4.5. Awn Length (AL)
3.4.6. Grain Shattering (SH) and Cold Tolerance (CT)
3.4.7. Identification of Loci Related to Quality Traits
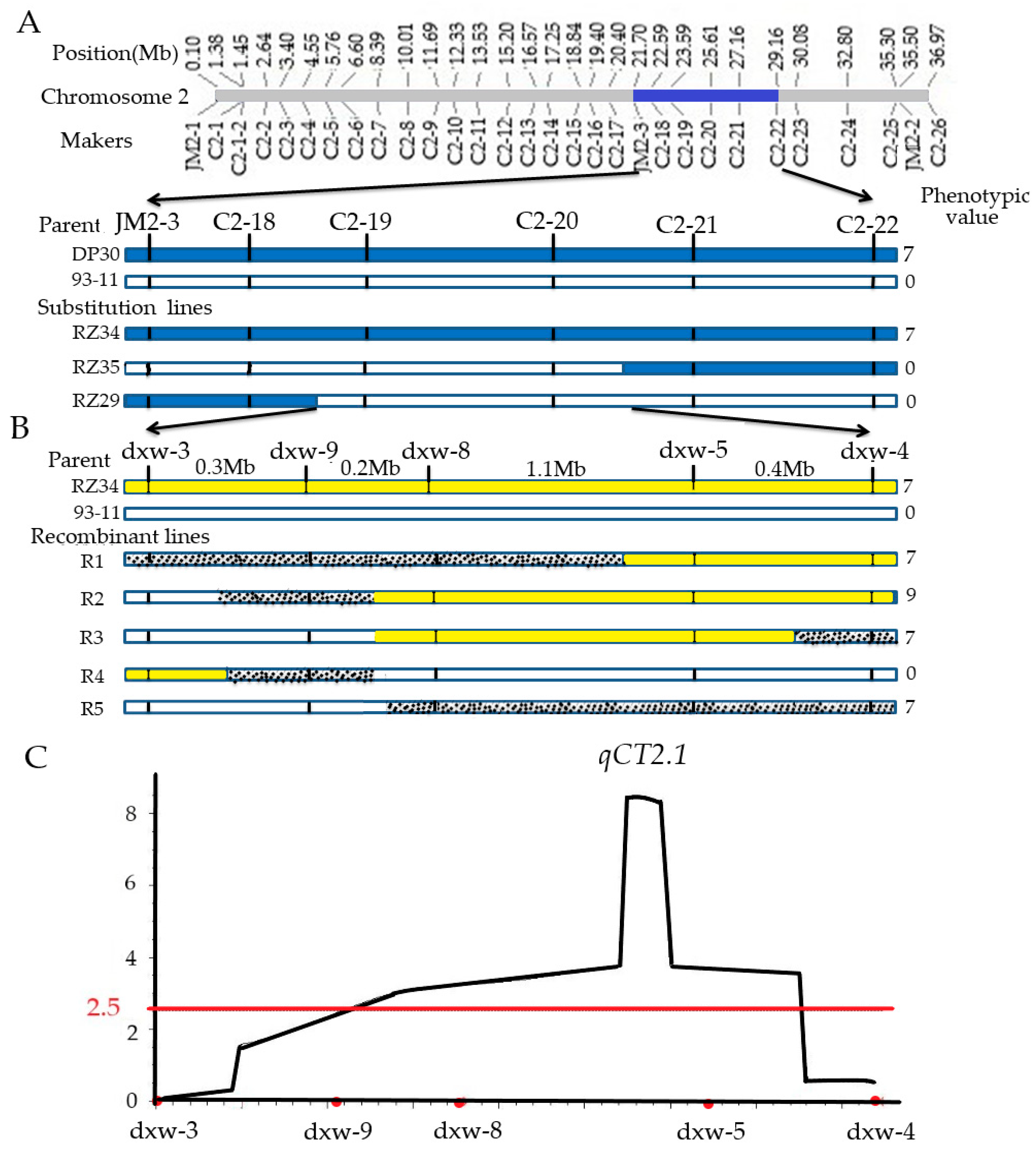
3.5. Construction of a Secondary Population and Mapping of qCT2.1
4. Discussion
4.1. Constructing CSSLs in Guangxi Wild Rice (O. rufipogon Griff.) DP30
4.2. Identification of Quantitative Trait Loci and Measurement of Various Traits in Fall and Spring
4.3. Construction of Secondary Population and Mapping of qCT2.1
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Doebley, J.F.; Gaut, B.S. The molecular genetics of crop domestication. Cell 2006, 127, 1309–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, M.; Sasaki, T. Genetic and molecular dissection of quantitative traits in rice. In Oryza: From Molecule to Plant; Springer: Dordrecht, The Netherlands, 1997; pp. 145–153. [Google Scholar]
- Li, Z.; Pinson, S. Identification of quantitative trait loci (QTLs) for heading date and plant height in cultivated rice (Oryza sativa L.). Theor. Appl. Genet. 1995, 91, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Ray, J.; Yu, L.; McCouch, S. Mapping quantitative trait loci associated with root penetration ability in rice (Oryza sativa L.). Theor. Appl. Genet. 1996, 92, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Redona, E.; Mackill, D. Mapping quantitative trait loci for seedling vigor in rice using RFLPs. Theor. Appl. Genet. 1996, 92, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Yano, M. Genetic and molecular dissection of naturally occurring variation. Curr. Opin. Plant Biol. 2001, 4, 130–135. [Google Scholar] [CrossRef]
- Fujita, D.; Ebron, L.A. Fine mapping of a gene for low-tiller number, Ltn, in japonica rice (Oryza sativa L.) variety Aikawa 1. Theor. Appl. Genet. 2010, 120, 1233–1240. [Google Scholar] [CrossRef]
- Eshed, Y.; Zamir, D. Less-than-additive epistatic interactions of quantitative trait loci in tomato. Genetics 1996, 143, 1807–1817. [Google Scholar] [CrossRef]
- Furuta, T.; Uehara, K. Development and evaluation of chromosome segment substitution lines (cssls) carrying chromosome segments derived from oryza rufipogon in the genetic background of Oryza Sativa L. Breed. Sci. 2014, 63, 468–475. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Sun, L. Genetic characterization of the chromosome single-segment substitution lines of O. glumaepatula and O. barthii and identification of QTLs for yield-related traits. Mol. Breed. 2019, 39, 51. [Google Scholar] [CrossRef]
- Wu, W.; Liu, X. A single-nucleotide polymorphism causes smaller grain size and loss of seed shattering during African rice domestication. Nat. Plants 2017, 3, 17064. [Google Scholar] [CrossRef]
- Lin, Y.; Tan, L. RLS3, a protein with AAAt domain localized in chloroplast, sustains leaf longevity in rice. J. Integr. Plant Biol. 2016, 58, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Jin, L. Analysis of epistasis among QTLs on heading date based on single segment substitution lines in rice. Sci. Rep. 2018, 8, 2071. [Google Scholar] [CrossRef] [PubMed]
- Kaneda, C.; Beachell, H.M. Response of indica-japonica rice hybrids to low temperatures. SABRAO J. 1973, 6, 17–32. [Google Scholar]
- Xiao, N.; Gao, Y.; Qian, H.; Gao, Q.; Wu, Y.; Zhang, D.; Zhang, X.; Yu, L.; Li, Y.; Pan, C. Identification of genes related to cold tolerance and a functional allele that confers cold tolerance. Plant Physiol. 2018, 177, 1108–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yiting, S.; Zhizhong, G. One SNP in cold1 determines cold tolerance during rice domestication. J. Genet. Genom. 2015, 42, 133–134. [Google Scholar] [CrossRef]
- Tan, L.; Li, X. Control of a key transition from prostrate to erect growth in rice domestication. Nat. Genet. 2008, 40, 1360–1364. [Google Scholar] [CrossRef]
- Wang, J.; Wang, J. Proteomic response of hybrid wild rice to cold stress at the seedling stage. PLoS ONE 2018, 13, e0198675. [Google Scholar] [CrossRef]
- Ando, T.; Yamamoto, T. Genetic dissection and pyramiding of quantitative traits for panicle architecture by using chromosomal segment substitution lines in rice. Theor. Appl. Genet. 2008, 116, 881–890. [Google Scholar] [CrossRef]
- Jiang, J.; Tan, L. Molecular evolution of the TAC1 gene from rice (Oryza sativa L.). J. Genet. Genom. 2012, 39, 551–560. [Google Scholar] [CrossRef]
- Saito, K.; Hayano-Saito, Y. Physical mapping and putative candidate gene identification of a quantitative trait locus Ctb1 for cold tolerance at the booting stage of rice. Theor. Appl. Genet. 2004, 109, 515–522. [Google Scholar] [CrossRef]
- Zhu, H.; Liu, Z. Detection and characterization of epistasis between qtls on plant height in rice using single segment substitution lines. Breed. Sci. 2015, 65, 192–200. [Google Scholar] [CrossRef] [Green Version]
- Ookawa, T.; Aoba, R. Precise estimation of genomic regions controlling lodging resistance using a set of reciprocal chromosome segment substitution lines in rice. Sci. Rep. 2016, 6, 30572. [Google Scholar] [CrossRef]
- Wingett, S.W.; Andrews, S. FastQ screen: A tool for multi-genome mapping and quality control. F1000Research 2018, 7, 1338. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows—Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawahara, Y.; de la Bastide, M. Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data. Rice 2013, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenna, A.; Hanna, M. The genome analysis toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, S.O.; Bendich, A.J. Extraction of DNA from plant tissues. In Plant Molecular Biology Manual; Springer: Dordrecht, The Netherlands, 1989; pp. 73–83. [Google Scholar]
- Qiao, W.; Qi, L. Development and characterization of chromosome segment substitution lines derived from Oryza rufipogon in the genetic background of O. sativa spp. indica cultivar 93-11. BMC. Genom. 2016, 17, 580. [Google Scholar] [CrossRef] [Green Version]
- Panaud, O.; Chen, X. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSLP) in rice (Oryza sativa L.). Mol. Gen. Genet. 1996, 252, 597–607. [Google Scholar] [CrossRef]
- Young, N.; Tanksley, S. Restriction segment length polymorphism maps and the concept of graphical genotypes. Theor. Appl. Genet. 1989, 77, 95–101. [Google Scholar] [CrossRef]
- Wan, X.; Weng, J. Quantitative trait loci (QTL) analysis for rice grain width and fine mapping of an identified QTL allele gw-5 in a recombination hotspot region on chromosome5. Genet 2008, 179, 2239–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCouch, S.R. Gene nomenclature system for rice. Rice 2008, 1, 72–84. [Google Scholar] [CrossRef] [Green Version]
- Voorrips, R. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, L.; Li, H.; Zhang, L.; Wang, J. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J. 2015, 3, 269–283. [Google Scholar] [CrossRef] [Green Version]
- Ye, H.; Feng, J. Map-based cloning of seed dormancy1-2 identified a gibberellin synthesis gene regulating the development of endosperm-imposed dormancy in rice. Plant Physiol. 2015, 169, 2152–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Jiao, G. GRAIN INCOMPLETE FILLING 2 regulates grain filling and starch synthesis during rice caryopsis development. J. Integr. Plant Biol. 2017, 59, 134–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, H.; Tanaka, A. Short grain1 decreases organ elongation and brassinosteroid response in rice. Plant. Physiol. 2012, 158, 1208–1219. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Liu, H. An-1 encodes a basic helix-loop-helix protein that regulates awn development, grain size, and grain number in rice. Plant. Cell 2013, 25, 3360–3376. [Google Scholar] [CrossRef] [Green Version]
- Gu, B.; Zhou, T. An-2 encodes a cytokinin synthesis enzyme that regulates awn length and grain production in rice. Mol. Plant 2015, 8, 1635–1650. [Google Scholar] [CrossRef]
- Lin, Z.; Griffith, M.E. Origin of seed shattering in rice (Oryza sativa L.). Planta 2007, 226, 11–20. [Google Scholar] [CrossRef]
- Xu, P.; Cai, W. RAN1 is involved in plant cold resistance and development in rice (Oryza sativa). J. Exp. Bot. 2014, 65, 3277–3287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanneganti, V.; Gupta, A.K. Overexpression of OsiSAP8, a member of stress associated protein (SAP) gene family of rice confers tolerance to salt, drought and cold stress in transgenic tobacco and rice. Plant Mol. Biol. 2008, 66, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Hwang, H. A rice orthologous of the ABA receptor, OsPYL/RCAR5, is a positive regulator of the ABA signal transduction pathway in seed germination and early seedling growth. J. Exp. Bot. 2011, 63, 1013–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandraratna, M.F. A gene for photoperiod sensitivity in rice linked with apiculus colour. Nature 1953, 171, 1162–1163. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.F.; Si, L. Genetic control of a transition from black to straw-white seed hull in rice domestication. Plant Phys. 2011, 155, 1301–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furukawa, T.; Maekawa, M. The Rc and Rd genes are involved in proanthocyanidin synthesis in rice pericarp. Plant J. 2007, 49, 91–102. [Google Scholar] [CrossRef]
- Ma, X.D.; Han, B. Construction of chromosome segment substitution lines of Dongxiang common wild rice (Oryza rufipogon Griff.) in the background of the japonica rice cultivar Nipponbare (Oryza sativa L.). Plant Physiol. Biochem. 2019, 144, 274–282. [Google Scholar] [CrossRef]
- Tan, L.B.; Zhang, P.J. Identification of quantitative trait loci controlling plant height and days to heading from Yuanjiang common wild rice (Oryza rufipogon Griff.). Acta Genet. Sinica 2004, 31, 1123–1128. [Google Scholar] [CrossRef]
- Andaya, V.; Mackill, D. Qtls conferring cold tolerance at the booting stage of rice using recombinant inbred lines from a japonica × indica cross. Theor. Appl. Genet. 2003, 106, 1084–1090. [Google Scholar] [CrossRef]
- Koji, S.; Yuriko, H.S. Map-based cloning of the rice cold tolerance gene ctb1. Plant Sci. 2010, 179, 97–102. [Google Scholar] [CrossRef]
- Zheng, J.X.; Ma, Z.F. Identification and mapping of QTLs for cold tolerance at the seedling stage in common wild rice (Oryza rufipogon). China J. Rice Sci. 2011, 25, 52–58. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chromosome | SNP No. (bp) | Chr. Length (bp) | SNP Percentage | Length (Mb) | Marker No. |
|---|---|---|---|---|---|
| 1 | 232,372 | 47,283,185 | 0.49% | 1.32 | 38 |
| 2 | 210,389 | 38,103,930 | 0.55% | 1.15 | 30 |
| 3 | 194,967 | 41,884,883 | 0.47% | 1.33 | 32 |
| 4 | 155,761 | 34,718,618 | 0.45% | 1.21 | 25 |
| 5 | 164,450 | 31,240,961 | 0.53% | 1.34 | 24 |
| 6 | 167,985 | 32,913,967 | 0.51% | 1.22 | 25 |
| 7 | 153,895 | 27,957,088 | 0.55% | 1.1 | 19 |
| 8 | 154,296 | 30,396,518 | 0.51% | 1.2 | 21 |
| 9 | 119,958 | 21,757,032 | 0.55% | 1.26 | 18 |
| 10 | 113,121 | 22,204,031 | 0.51% | 1.21 | 17 |
| 11 | 107,638 | 23,035,369 | 0.47% | 1.24 | 18 |
| 12 | 119,271 | 23,049,917 | 0.52% | 1.18 | 18 |
| Average | 157,842 | 31,212,125 | 0.51% | 1.23 | 24 |
| Total | 1,894,103 | 374,545,499 | 6.10% | 285 |
| Chromosome | No. of CSSLs | Total Length of Target Segment (Mb) | Length of Chromosome (Mb) | Coverage Length (Mb) | Coverage Rate (%) |
|---|---|---|---|---|---|
| 1 | 17 | 72 | 47 | 44.5 | 94.7 |
| 2 | 17 | 61 | 38 | 36.5 | 96.1 |
| 3 | 17 | 52.5 | 41 | 38.1 | 93.3 |
| 4 | 15 | 66.6 | 35 | 31.9 | 91.2 |
| 5 | 11 | 41.9 | 32 | 29.4 | 92.3 |
| 6 | 15 | 56.8 | 33 | 28.7 | 87.9 |
| 7 | 6 | 31.9 | 28 | 26.6 | 95.5 |
| 8 | 11 | 29.6 | 30 | 24.9 | 83.3 |
| 9 | 5 | 21.7 | 22 | 19.1 | 87.5 |
| 10 | 7 | 31.6 | 22 | 19.4 | 88.6 |
| 11 | 4 | 26.2 | 23 | 21.4 | 93.1 |
| 12 | 7 | 24.9 | 23 | 21.7 | 93.6 |
| Traits | QTL | LOD | Position | Spring | Fall | Cloned Gene | ||
|---|---|---|---|---|---|---|---|---|
| a | ap (%) | a | ap (%) | |||||
| TA | qTA1.1 | 2.4 | C1-26–C1-27 | −1.6 | 9.7 | 0.7 | 9.7 | |
| TA | qTA7.1 | 2.5 | C7-2 | −2.4 | 17.2 | −1.2 | 14 | PROG1 [17] |
| TA | qTA9.1 | 2.7 | C9-14–C9-15 | −1.4 | 16.5 | −1.4 | 16.5 | TAC1 [20] |
| TA | qTA2.1 | 2.6 | C2-25 | 0.4 | 4.3 | 0.6 | 7.3 | |
| HD | qHD11.1 | 3 | C11-4 | −5 | 4.1 | −3.72 | 2.9 | |
| PH | qPH1.1 | 3.1 | JM1-5 | 31.8 | 17.6 | 30.7 | 28.8 | Sd1 [37] |
| PH | qPH6.1 | 3 | C6-8 | 28.1 | 24.6 | 27.3 | 25.6 | |
| PH | qPH7.1 | 2.6 | C7-5 | 25.9 | 19.5 | 26.7 | 25 | |
| NGPP | qNGPP3.1 | 2.8 | C3-19 | 34.2 | 19.8 | 33.1 | 20.1 | |
| NGPP | qNGPP4.1 | 5.2 | C4-12 | 25.2 | 14.6 | 25 | 15.2 | |
| NGPP | qNGPP12.1 | 2.9 | C12-2 | 40.3 | 23.3 | 37.3 | 22.6 | |
| GWT | qGWT1.1 | 3.1 | C1-19 | −3 | 10.2 | −0.3 | 11.1 | OsAGPL2 [38] |
| GWT | qGWT2.1 | 4.2 | C2-19 | −0.2 | 8 | −0.2 | 7.8 | |
| GWT | qGWT3.1 | 4.4 | C3-25 | −0.3 | 8.3 | −0.3 | 8.3 | |
| GWT | qGWT4.1 | 3.4 | C4-15 | −0.3 | 10 | −0.3 | 9.9 | |
| GWT | qGWT5.1 | 4 | C5-23 | −0.3 | 10 | −0.3 | 10.1 | |
| GL | qGL3.1 | 3.8 | C3-19 | −0.9 | 9 | 1 | 10 | |
| GL | qGL9.1 | 3.4 | C9-12 | −0.9 | 8.5 | −0.9 | 8.7 | SG1 [39] |
| GW | qGW9.1 | 3.2 | C9-8 | −0.1 | 3.2 | −0.1 | 2.7 | |
| GW | qGW10.1 | 3.1 | C10-3 | −0.1 | 3.4 | −0.1 | 3.1 | |
| GLWR | qGLWR3.1 | 3.5 | C3-26 | 0.3 | 11.4 | 0.3 | 11.1 | |
| GLWR | qGLWR8.1 | 3.9 | C8-2 | 0.3 | 10.9 | 0.3 | 11.2 | |
| AL | qAL1.1 | 3 | C1-7 | 5.6 | 26.9 | 5.6 | 32.7 | |
| AL | qAL4.1 | 2.7 | C4-8–C4-10 | 9.8 | 47.6 | 10.1 | 59.3 | An-1 [40] |
| AL | qAL4.2 | 4.3 | C4-19 | 10.3 | 49.6 | 10.3 | 60.8 | An-2 [41] |
| AL | qAL8.1 | 2.6 | C8-11 | 6.8 | 33.1 | 6.4 | 37.7 | |
| AL | qAL11.1 | 3.9 | C11-2 | 6.3 | 30.6 | 6.9 | 40.5 | |
| SH | qSH4.1 | 3.4 | C4-22 | −12.5 | −8.3 | −12 | −9 | SH4 [42] |
| SH | qSH11.1 | 2 | C11-5~C11-8 | −40 | −26.7 | −42 | −27.7 | |
| CT | qCT1.1 | 2.6 | C1-16 | 1.3 | 48.5 | 1.2 | 53.3 | OsRAN1 [43] |
| CT | qCT2.1 | 2.6 | C2-19 | 1.2 | 46.2 | 0.9 | 42.9 | |
| CT | qCT3.1 | 2.9 | C3-2 | 1.2 | 45.8 | 1.1 | 48.1 | |
| CT | qCT5.1 | 2.1 | C5-20 | 1.8 | 67.3 | 1.4 | 65.2 | OsiSAP8 [44] |
| CT | qCT6.1 | 2.4 | C6-20 | 1.8 | 68.4 | 1.4 | 61.4 | OsPYL9 [45] |
| CT | qCT10.1 | 3 | C10-3 | 1.4 | 51.9 | 1.4 | 64.8 | |
| CT | qCT12.1 | 2.2 | C12-11 | 1.2 | 46.8 | 1 | 46.5 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, R.; Zhao, N.; Usman, B.; Luo, L.; Liao, S.; Qin, Y.; Nawaz, G.; Li, R. Development of Chromosome Segment Substitution Lines (CSSLs) Derived from Guangxi Wild Rice (Oryza rufipogon Griff.) under Rice (Oryza sativa L.) Background and the Identification of QTLs for Plant Architecture, Agronomic Traits and Cold Tolerance. Genes 2020, 11, 980. https://doi.org/10.3390/genes11090980
Yuan R, Zhao N, Usman B, Luo L, Liao S, Qin Y, Nawaz G, Li R. Development of Chromosome Segment Substitution Lines (CSSLs) Derived from Guangxi Wild Rice (Oryza rufipogon Griff.) under Rice (Oryza sativa L.) Background and the Identification of QTLs for Plant Architecture, Agronomic Traits and Cold Tolerance. Genes. 2020; 11(9):980. https://doi.org/10.3390/genes11090980
Chicago/Turabian StyleYuan, Ruizhi, Neng Zhao, Babar Usman, Liang Luo, Shanyue Liao, Yufen Qin, Gul Nawaz, and Rongbai Li. 2020. "Development of Chromosome Segment Substitution Lines (CSSLs) Derived from Guangxi Wild Rice (Oryza rufipogon Griff.) under Rice (Oryza sativa L.) Background and the Identification of QTLs for Plant Architecture, Agronomic Traits and Cold Tolerance" Genes 11, no. 9: 980. https://doi.org/10.3390/genes11090980
APA StyleYuan, R., Zhao, N., Usman, B., Luo, L., Liao, S., Qin, Y., Nawaz, G., & Li, R. (2020). Development of Chromosome Segment Substitution Lines (CSSLs) Derived from Guangxi Wild Rice (Oryza rufipogon Griff.) under Rice (Oryza sativa L.) Background and the Identification of QTLs for Plant Architecture, Agronomic Traits and Cold Tolerance. Genes, 11(9), 980. https://doi.org/10.3390/genes11090980