
Sorghum Brown Midrib19 (Bmr19) Gene Links Lignin Biosynthesis to Folate Metabolism

 , , ,
, , ,
Abstract
:1. Introduction
1.1. Biological Role of Lignin
1.2. Significance of Lignin in Biofuel Production
1.3. Biosynthesis of Lignin, Composition of Monolignols and Their Polymerization
1.4. Brown Midrib Mutations
1.5. Allelic Groups among the bmr Mutants of Sorghum
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction and Construction of the Bulked Pools
2.3. Construction of Libraries and Illumina Sequencing
2.4. Construction of Reference-Based Assemblies
2.5. Indel Markers of the Bmr12 Gene
2.6. Validation of Identified bmr19 Mutant SNP
2.7. Alternate bmr19 Alleles
2.8. Biomass Composition Analysis
3. Results
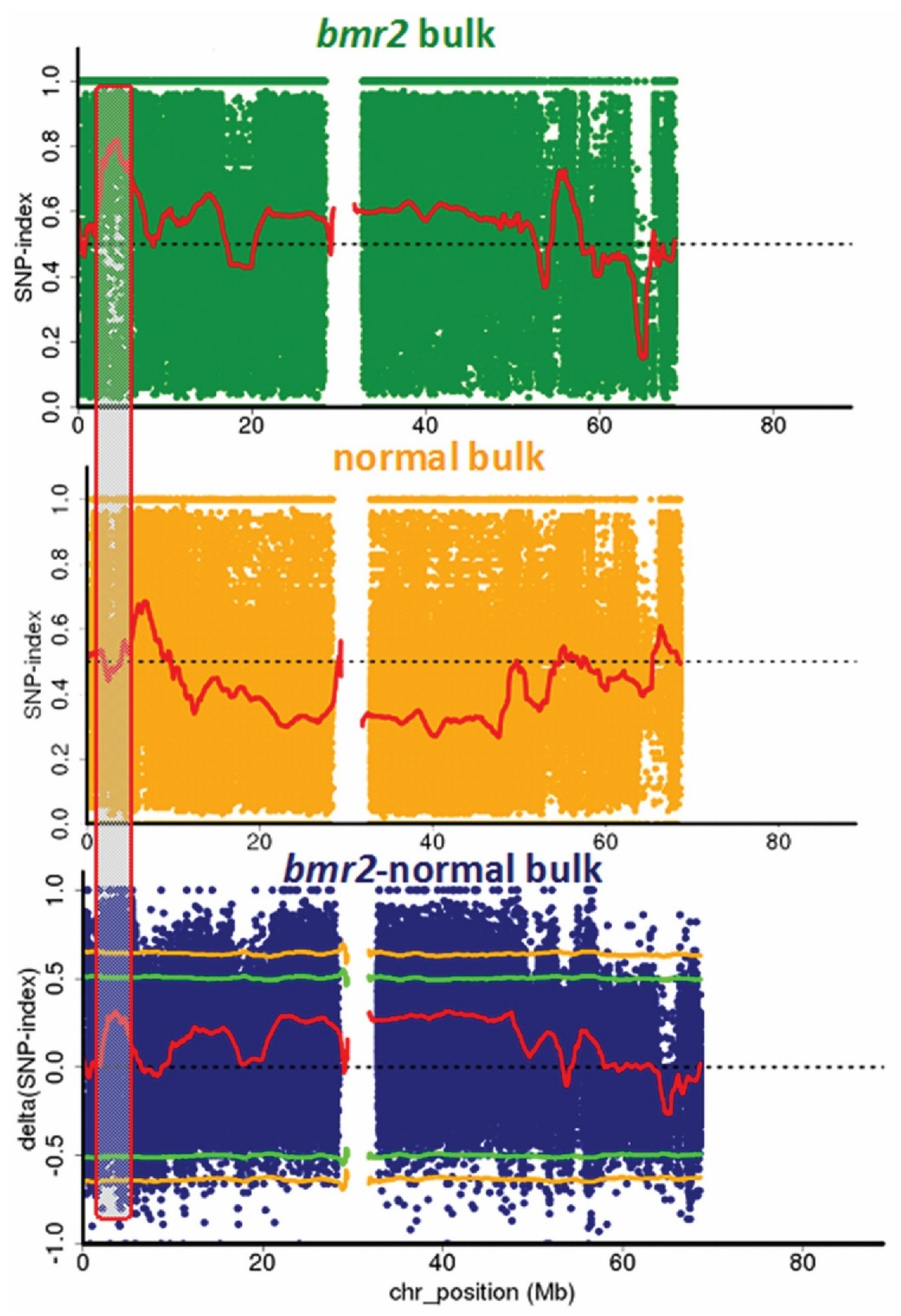
3.1. Confirmation of the bmr2-ref Allele in the bmr2 Bulk
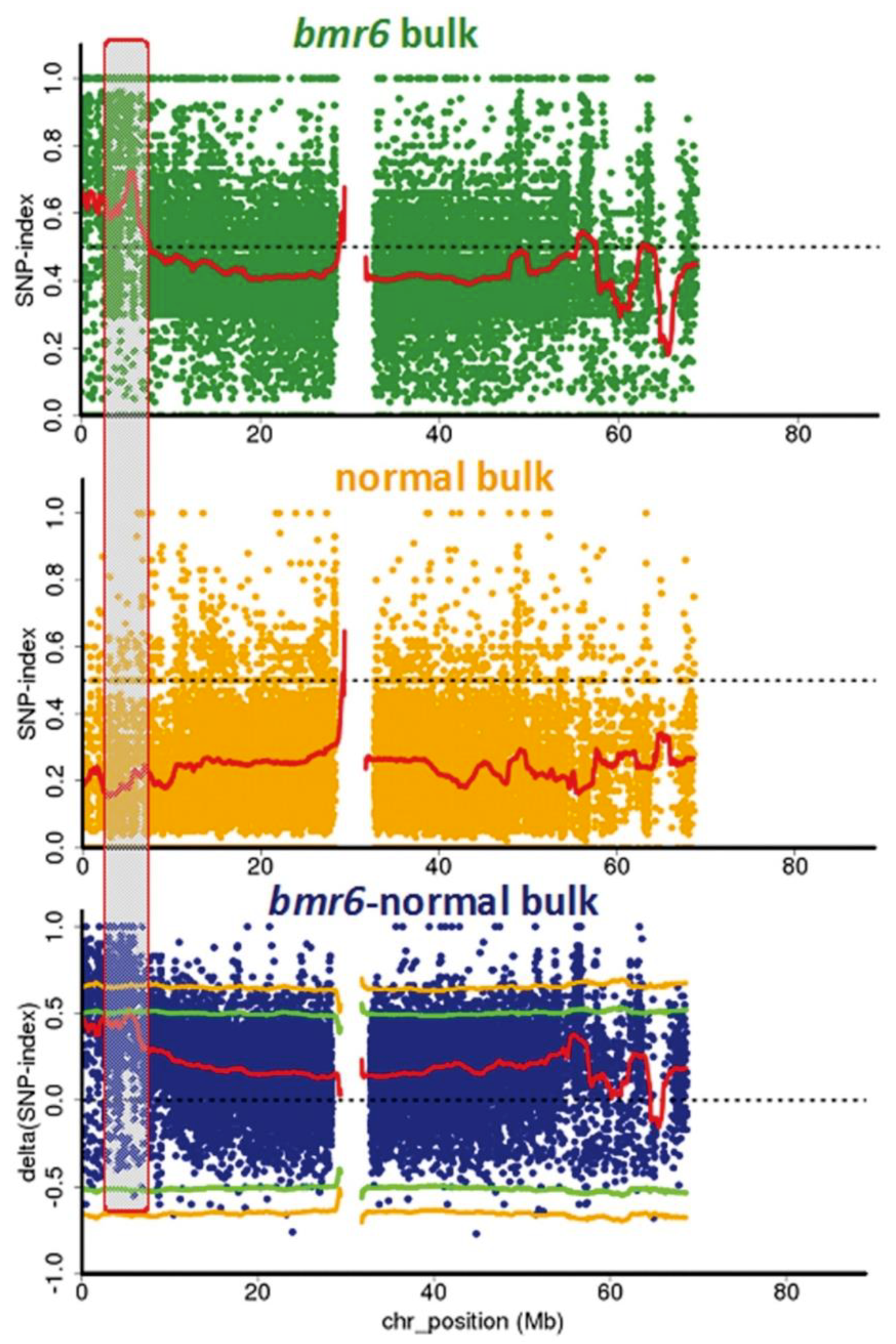
3.2. Confirmation of the bmr6-ref Allele in the bmr6 Bulk
3.3. Comparison of the bmr12 and Normal Bulks
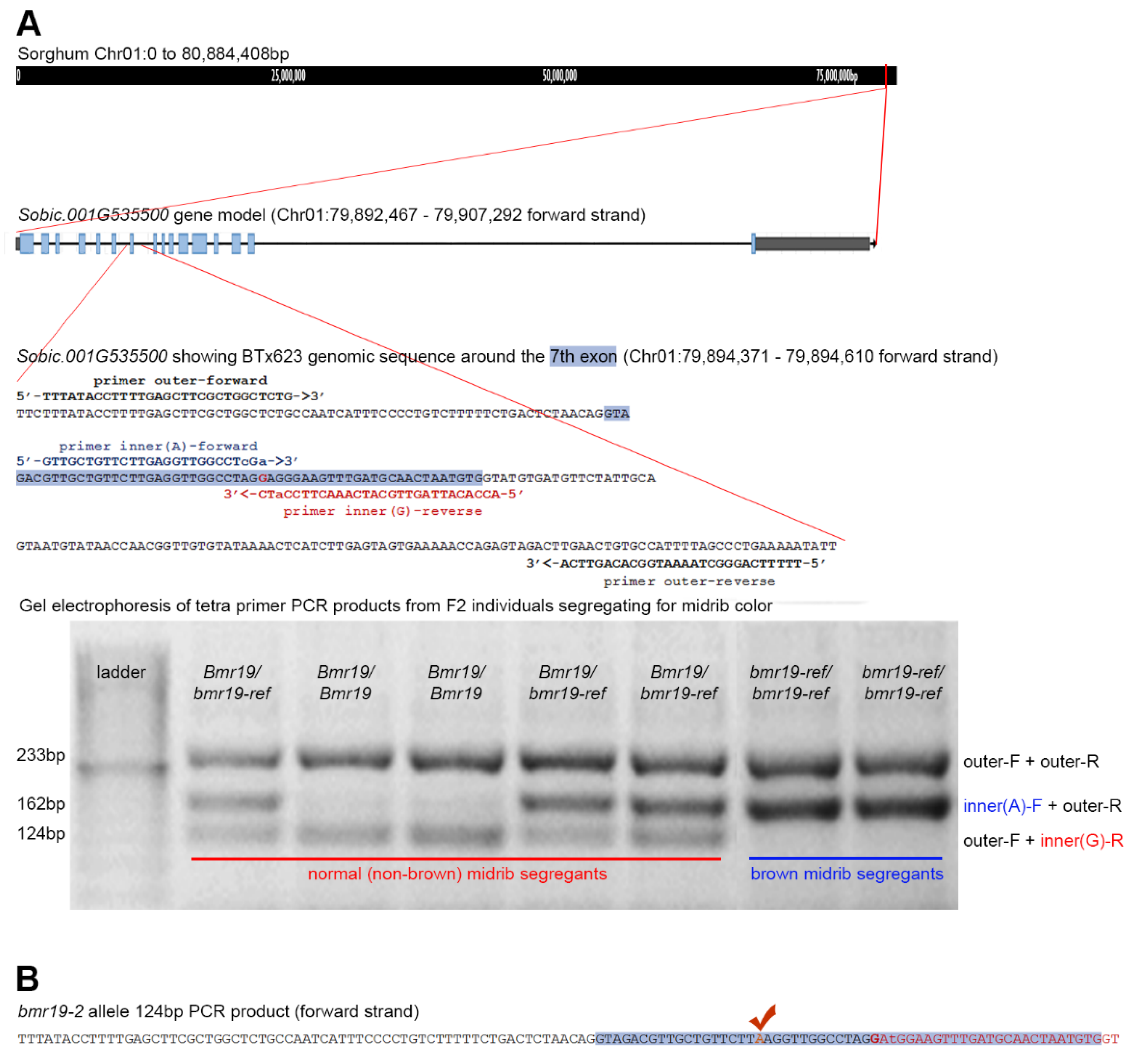
3.4. Identification of the Bmr19 Gene
3.5. Association of the bmr19-ref Allele with the Brown Midrib Phenotype Outside the Mapping Population
3.6. Alternate Alleles
3.7. Effects on Feedstock
4. Discussion
4.1. Power of the Analysis of Whole Genome Resequenced Bulk Comparisons
4.2. The Bmr19 Locus
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cesarino, I.; Simões, M.S.; dos Santos Brito, M.; Fanelli, A.; da Franca Silva, T.; Romanel, E. Building the wall: Recent advances in understanding lignin metabolism in grasses. Acta Physiol. Plant 2016, 38, 269–282. [Google Scholar] [CrossRef]
- Hatfield, R.D.; Marita, J.M.; Hatfield, R.D.; Marita, J.M. Maize development: Cell wall changes in leaves and sheaths. Am. J Plant Sci. 2017, 8, 1248–1263. [Google Scholar] [CrossRef]
- Li, H.; Huang, Y. Expression of brown-midrib in a spontaneous sorghum mutant is linked to a 5′-UTR deletion in lignin biosynthesis gene SbCAD2. Sci. Rep. 2017, 7, 11664. [Google Scholar] [CrossRef] [Green Version]
- Börjesson, P.; Tufvesson, L.M. Agricultural crop-based biofuels—Resource efficiency and environmental performance including direct land use changes. J. Clean. Prod. 2011, 19, 108–120. [Google Scholar] [CrossRef]
- Li, X.; Zheng, Y. Lignin-enzyme interaction: Mechanism, mitigation approach, modeling, and research prospects. Biotechnol. Adv. 2017, 35, 466–489. [Google Scholar] [CrossRef] [PubMed]
- Saballos, A.; Ejeta, G.; Sanchez, E.; Kang, C.; Vermerris, W. A genomewide analysis of the cinnamyl alcohol dehydrogenase family in sorghum [Sorghum bicolor (L.) Moench] identifies SbCAD2 as the brown midrib6 gene. Genetics 2009, 181, 783–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saballos, A.; Sattler, S.E.; Sanchez, E.; Foster, T.P.; Xin, Z.; Kang, C.; Pedersen, J.F.; Vermerris, W. Brown midrib2 (Bmr2) encodes the major 4-coumarate: Coenzyme A ligase involved in lignin biosynthesis in sorghum (Sorghum bicolor (L.) Moench). Plant J. 2012, 70, 818–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, B.; Li, C.; Tarczynski, M.C. High free-methionine and decreased lignin content result from a mutation in the Arabidopsis S-adenosyl-L-methionine synthetase 3 gene. Plant J. 2002, 29, 371–380. [Google Scholar] [CrossRef]
- Wang, Y.; Chantreau, M.; Sibout, R.; Hawkins, S. Plant cell wall lignification and monolignol metabolism. Front. Plant Sci. 2013, 4, 220. [Google Scholar] [CrossRef] [Green Version]
- Sattler, S.E.; Funnell-Harris, D.L.; Pedersen, J.F. Brown midrib mutations and their importance to the utilization of maize, sorghum, and pearl millet lignocellulosic tissues. Plant Sci. 2010, 178, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Umezawa, T.; Sakamoto, M.; Koshiba, T. Cinnamyl alcohol dehydrogenase deficiency causes the brown midrib phenotype in rice. In Biofuels: Greenhouse Gas Mitigation and Global Warming; Kumar, A., Ogita, S., Yau, Y., Eds.; Springer: New Delhi, India, 2018; pp. 375–385. [Google Scholar]
- Koshiba, T.; Murakami, S.; Hattori, T.; Mukai, M.; Takahashi, A.; Miyao, A.; Hirochika, H.; Suzuki, S.; Sakamoto, M.; Umezawa, T. CAD2 deficiency causes both brown midrib and gold hull and internode phenotypes in Oryza sativa L. cv. Nipponbare. Plant Biotechnol. 2013, 30, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Porter, K.S.; Axtell, J.D.; Lechtenberg, V.L.; Colenbrander, V.F. Phenotype, fiber composition, and in vitro dry matter disappearance of chemically induced brown midrib (bmr) mutants of sorghum. Crop. Sci. 1978, 18, 205–208. [Google Scholar] [CrossRef]
- Saballos, A.; Vermerris, W.; Rivera, L.; Ejeta, G. Allelic association, chemical characterization and saccharification properties of brown midrib mutants of sorghum (Sorghum bicolor (L.) Moench). BioEnergy Res. 2008, 1, 193–204. [Google Scholar] [CrossRef]
- Sattler, S.E.; Saballos, A.; Xin, Z.; Funnell-Harris, D.L.; Vermerris, W.; Pedersen, J.F. Characterization of novel sorghum brown midrib mutants from an EMS-mutagenized population. G3 2014, 4, 2115–2124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halpin, C.; Holt, K.; Chojecki, J.; Oliver, D.; Chabbert, B.; Monties, B.; Edwards, K.; Barakate, A.; Foxon, G.A. Brown-midrib maize (bm1)—A mutation affecting the cinnamyl alcohol dehydrogenase gene. Plant J. 1998, 14, 545–553. [Google Scholar] [CrossRef]
- Sattler, S.E.; Saathoff, A.J.; Haas, E.J.; Palmer, N.A.; Funnell-Harris, D.L.; Sarath, G.; Pedersen, J.F. A nonsense mutation in a cinnamyl alcohol dehydrogenase gene is responsible for the sorghum brown midrib6 phenotype. Plant Physiol. 2009, 150, 584–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorthy, S.; Mayandi, K.; Faldu, D.; Dalal, M. Molecular characterization of allelic variation in spontaneous brown midrib mutants of sorghum (Sorghum bicolor (L.) Moench). Mol. Breed. 2013, 31, 795–803. [Google Scholar] [CrossRef]
- Scully, E.D.; Gries, T.; Funnell-Harris, D.L.; Xin, Z.; Kovacs, F.A.; Vermerris, W.; Sattler, S.E. Characterization of novel Brown midrib 6 mutations affecting lignin biosynthesis in sorghum. J. Integr. Plant Biol. 2016, 58, 136–149. [Google Scholar] [CrossRef] [Green Version]
- Dien, B.S.; Sarath, G.; Pedersen, J.F.; Sattler, S.E.; Chen, H.; Funnell-Harris, D.L.; Nichols, N.N.; Cotta, M.A. Improved sugar conversion and ethanol yield for forage sorghum (Sorghum bicolor L. Moench) lines with reduced lignin contents. BioEnergy Res. 2009, 2, 153–164. [Google Scholar] [CrossRef]
- Bout, S.; Vermerris, W. A candidate-gene approach to clone the sorghum Brown midrib gene encoding caffeic acid O-methyltransferase. Mol. Gen. Genet. 2003, 269, 205–214. [Google Scholar] [CrossRef]
- Vignols, F.; Rlgau, J.; Torres, M.A.; Capellades, M.; Puigdomènech, P. The brown midrib3 (bm3) mutation in maize occurs in the gene encoding caffeic acid O-methyltransferase. Plant Cell 1995, 7, 407–416. [Google Scholar] [PubMed] [Green Version]
- Bittinger, T.S.; Cantrell, R.P.; Axtell, J.D. Allelism tests of the brown- midrib mutants of sorghum. J. Hered. 1981, 72, 147–148. [Google Scholar] [CrossRef]
- Xin, Z.; Wang, M.L.; Burow, G.; Burke, J. An induced sorghum mutant population suitable for bioenergy research. BioEnergy Res. 2009, 2, 10–16. [Google Scholar] [CrossRef]
- Sattler, S.; Palmer, N.; Saballos, A.; Greene, A.M.; Xin, Z.; Sarath, G.; Vermerris, W.; Pedersen, J.F. Identification and characterization of four missense mutations in Brown midrib 12 (Bmr12), the caffeic O-methyltranferase (COMT) of sorghum. BioEnergy Res. 2012, 5, 855–865. [Google Scholar] [CrossRef] [Green Version]
- Eudes, A.; Dutta, T.; Deng, K.; Jacquet, N.; Sinha, A.; Benites, V.T.; Baidoo, E.E.; Richel, A.; Sattler, S.E.; Northen, T.R.; et al. SbCOMT (Bmr12) is involved in the biosynthesis of tricin-lignin in sorghum. PLoS ONE 2017, 12, e0178160. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Kosugi, S.; Natsume, S.; Yoshida, K.; MacLean, D.; Cano, L.; Kamoun, S.; Terauchi, R. Coval: Improving alignment quality and variant calling accuracy for next-generation sequencing data. PLoS ONE 2013, 8, e75402. [Google Scholar] [CrossRef] [Green Version]
- Abe, A.; Kosugi, S.; Yoshida, K.; Natsume, S.; Takagi, H.; Kanzaki, H.; Matsumura, H.; Yoshida, K.; Mitsuoka, C.; Tamiru, M.; et al. Genome sequencing reveals agronomically important loci in rice using MutMap. Nat. Biotechnol. 2012, 30, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, M.; Huber, W.; Pages, H.; Aboyoun, P.; Carlson, M.; Gentleman, R.; Morgan, M.T.; Carey, V.J. Software for computing and annotating genomic ranges. PLoS Comput. Biol. 2013, 9, e1003118. [Google Scholar] [CrossRef]
- Obenchain, V.; Lawrence, M.; Carey, V.; Gogarten, S.; Shannon, P.; Morgan, M. VariantAnnotation: A Bioconductor package for exploration and annotation of genetic variants. Bioinformatics 2014, 30, 2076–2078. [Google Scholar] [CrossRef] [Green Version]
- Mace, E.S.; Tai, S.; Gilding, E.K.; Li, Y.; Prentis, P.J.; Bian, L.; Cruickshank, A.; Campbell, B.C.; Hu, W.; Innes, D.J.; et al. Whole-genome sequencing reveals untapped genetic potential in Africa’s indigenous cereal crop sorghum. Nat. Commun. 2013, 4, 2320. [Google Scholar] [CrossRef] [Green Version]
- Paterson, A.H.; Bowers, J.E.; Bruggmann, R.; Grimwood, J.; Gundlach, H.; Haberer, G.; Hellsten, U.; Mitros, T.; Poliakov, A.; Schmutz, J.; et al. The Sorghum bicolor genome and the diversification of grasses. Nature 2008, 457, 551–556. [Google Scholar] [CrossRef] [Green Version]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative Genomics Viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [Green Version]
- Rivera Burgos, L.A. Genetic, Agronomic and Compositional Characterization of Brown Midrib Sweet Sorghum Lignocellulosic Biomass for Ethanol Production. Ph.D. Dissertation, Purdue University, West Lafayette, IN, USA, 2015. [Google Scholar]
- Medrano, R.F.; de Oliveira, C.A. Guidelines for the tetra-primer ARMS-PCR technique development. Mol. Biotechnol. 2014, 56, 599–608. [Google Scholar] [CrossRef]
- Vogel, K.P.; Pedersen, J.F.; Masterson, S.D.; Toy, J.J. Evaluation of a filter bag system for NDF, ADF, and IVDMD forage analysis. Crop. Sci. 1999, 39, 276–279. [Google Scholar] [CrossRef]
- Sarath, G.; Akin, D.E.; Mitchell, R.B.; Vogel, K.P. Cell-wall composition and accessibility to hydrolytic enzymes is differentially altered in divergently bred switchgrass (Panicum virgatum L.) genotypes. Appl. Biochem. Biotechnol. 2008, 150, 1–14. [Google Scholar] [CrossRef]
- Palmer, N.A.; Sattler, S.E.; Saathoff, A.J.; Funnell, D.; Pedersen, J.F.; Sarath, G. Genetic background impacts soluble and cell wall-bound aromatics in brown midrib mutants of sorghum. Planta 2008, 229, 115–127. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Hill-Skinner, S.; Liu, S.; Beuchle, D.; Tang, H.M.; Yeh, C.T.; Nettleton, D.; Schnable, P.S. The maize brown midrib4 (bm4) gene encodes a functional folylpolyglutamate synthase. Plant J. 2015, 81, 493–504. [Google Scholar] [CrossRef] [Green Version]
- Marita, J.M.; Vermerris, W.; Ralph, J.; Hatfield, R.D. Variations in the cell wall composition of maize brown midrib mutants. J. Agric. Food Chem. 2003, 51, 1313–1321. [Google Scholar] [CrossRef]
- Srivastava, A.C.; Chen, F.; Ray, T.; Pattathil, S.; Peña, M.J.; Avci, U.; Li, H.; Huhman, D.V.; Backe, J.; Urbanowicz, B.; et al. Loss of function of folylpolyglutamate synthetase 1 reduces lignin content and improves cell wall digestibility in Arabidopsis. Biotechnol. Biofuels 2015, 8, 224. [Google Scholar] [CrossRef] [Green Version]
- Mehrshahi, P.; Gonzalez-Jorge, S.; Akhtar, T.A.; Ward, J.L.; Santoyo-Castelazo, A.; Marcus, S.E.; Lara-Núñez, A.; Ravanel, S.; Hawkins, N.D.; Beale, M.H.; et al. Functional analysis of folate polyglutamylation and its essential role in plant metabolism and development. Plant J. 2010, 64, 267–279. [Google Scholar] [CrossRef] [Green Version]
- Hanson, A.D.; Roje, S. One-carbon metabolism in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 119–137. [Google Scholar] [CrossRef] [PubMed]
- Barrière, Y.; Ralph, J.; Méchin, V.; Guillaumie, S.; Grabber, J.H.; Argillier, O.; Chabbert, B.; Lapierre, C. Genetic and molecular basis of grass cell wall biosynthesis and degradability. II. Lessons from brown-midrib mutants. Comptes Rendus Biol. 2004, 327, 847–860. [Google Scholar] [CrossRef]
- Mazarei, M.; Baxter, H.L.; Srivastava, A.; Li, G.; Xie, H.; Dumitrache, A.; Rodriguez, M., Jr.; Natzke, J.M.; Zhang, J.Y.; Turner, G.B.; et al. Silencing folylpolyglutamate synthetase1 (FPGS1) in switchgrass (Panicum virgatum L.) improves lignocellulosic biofuel production. Front. Plant Sci. 2020, 11, 843. [Google Scholar] [CrossRef]
- Xie, H.; Engle, N.L.; Venketachalam, S.; Yoo, C.G.; Barros, J.; Lecoultre, M.; Howard, N.; Li, G.; Sun, L.; Srivastava, A.C.; et al. Combining loss of function of FOLYLPOLYGLUTAMATE SYNTHETASE1 and CAFFEOYL-COA 3-O-METHYLTRANSFERASE1 for lignin reduction and improved saccharification efficiency in Arabidopsis thaliana. Biotechnol. Biofuels 2019, 12, 108–127. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal (Non-Brown) Bulk | bmr2 Bulk | bmr6 Bulk | bmr12 Bulk | bmr19 Bulk | |||||
|---|---|---|---|---|---|---|---|---|---|
| Pedigree | Relevant Alleles in Homozygous State | Pedigree | Relevant Alleles | Pedigree | Relevant Alleles | Pedigree | Relevant Alleles | Pedigree | Relevant Alleles |
| PP290 | Bmr2, Bmr6, Bmr12, Bmr19 | (bmr2ms3 × PP290)BC3F5 | bmr2-ref | (bmr6ms3 × PP290)BC3F5 | bmr6-ref | (bmr18ms3 × PP290)BC3F5 | bmr12-18 | (bmr19ms3 × PP290)BC2F5 | bmr19-ref |
| PRL983999 | Bmr2, Bmr6, Bmr12, Bmr19 | (bmr2ms3 × PRL983999)BC3F4 | bmr2-ref | (bmr6ms3 × PRL983999)BC3F4 | bmr6-ref | (bmr12ms3 × PRL983999)F4 | bmr12-ref | (bmr19ms3 × PRL983999)F4 | bmr19-ref |
| Sudan Zera-Zera | Bmr2, Bmr6, Bmr12, Bmr19 | (bmr2ms3 × Sudan Zera-Zera)F4 | bmr2-ref | (bmr6ms3 × Sudan Zera-Zera)BC3F4 | bmr6-ref | (bmr18ms3 × Sudan Zera-Zera)BC3F4 | bmr12-18 | (bmr19ms3 × Sudan Zera-Zera)BC2F5 | bmr19-ref |
| PU216B | Bmr2, Bmr6, Bmr12, Bmr19 | (bmr2ms3 × PU216B)BC2F5 | bmr2-ref | (bmr6ms3 × PU216B)BC3F5 | bmr6-ref | (bmr12ms3 × PU216B)BC3F5 | bmr12-ref | (bmr19ms3 × PU216B)BC3F3 | bmr19-ref |
| P942242B | Bmr2, Bmr6, Bmr12, Bmr19 | (bmr2ms3 × P942242B)BC3F4 | bmr2-ref | (bmr6ms3 × P942242B)BC3F4 | bmr6-ref | (bmr18ms3 × P942242B)BC3F4 | bmr12-18 | (bmr19ms3 × P942242B)BC2F4 | bmr19-ref |
| P90344 | Bmr2, Bmr6, Bmr12, Bmr19 | (bmr6ms3 × P90344)BC2F5 | bmr6-ref | (bmr18ms3 × P90344)BC2F5 | bmr12-18 | (bmr19ms3 × P90344)BC3F3 | bmr19-ref | ||
| P90812 | Bmr2, Bmr6, Bmr12, Bmr19 | (bmr2ms3 × P90812)F4 | bmr2-ref | (bmr3ms3 × P90812)BC2F5 | bmr6-3 | (bmr12ms3 × P90812)BC2F5 | bmr12-ref | (bmr19ms3 × P90812)BC2F5 | bmr19-ref |
| Tx623B | Bmr2, Bmr6, Bmr12, Bmr19 | (bmr2ms3 × Tx623B)BC3F5 | bmr2-ref | (bmr6ms3 × Tx623B)BC3F5 | bmr6-ref | (BTx623ms3 × bmr18)BC2F4 | bmr12-18 | (BTx623ms3 × bmr19)BC2F4 | bmr19-ref |
| P9401 | Bmr2, Bmr6, Bmr12, Bmr19 | (bmr5ms3 × P9401)BC4F5 | bmr2-5 | (bmr3ms3 × P9401)BC4F5 | bmr6-3 | (bmr7ms3 × P9401)BC4F5 | bmr12-7 | (P9401ms3 × bmr19)BC3F5 | bmr19-ref |
| Line Name | NPGS Accession Number | SRA Accession Number | Mutation Position on Chr01 * | Base Pair Change | Amino Acid Change Chemical Characteristic Change | Leaf Midrib Phenotype | Mutant Allele Name |
|---|---|---|---|---|---|---|---|
| 0623 | PI 677943 | SRR2759749 | 79,894,180 | C → T | A221V hydrophobic → hydrophobic | normal | |
| 2354 | PI 678119 | SRR2759203 | 79,894,457 | G → A | E236K negative → positive | brown | bmr19-2 |
| 2465 | PI 678132 | SRR2759494 | 79,895,561 | G → A | S378N polar → polar | normal | |
| 0662 | PI 677955 | SRR2759755 | 79,894,460 | G → A | V237I nonpolar → nonpolar | normal | |
| bmr19 | 79,894,470 | G → A | G240E nonpolar → negative | brown | bmr19-ref |
| Line | Relevant Alleles in Homozygosity | Neutral Detergent Fiber (Cellulose, Hemicellulose and Lignin) | Acid Detergent Fiber (Cellulose and Lignin) | Acid Detergent Lignin (Lignin) | |||
|---|---|---|---|---|---|---|---|
| Mean * | Std Dev | Mean | Std Dev | Mean | Std Dev | ||
| BTx623 | Bmr2, Bmr6, Bmr12, Bmr19 | 63.1 a | 5.7 | 37.5 a | 4.0 | 5.0 a | 0.6 |
| (bmr2ms3 × Tx623B)BC3F5 | bmr2-ref | 58.9 ab | 6.0 | 34.1 ab | 3.9 | 2.6 c | 0.4 |
| (bmr6ms3 × Tx623B)BC3F5 | bmr6-ref | 55.8 b | 8.1 | 31.8 b | 5.4 | 2.5 c | 0.3 |
| (BTx623ms3 × bmr18)BC2F4 | bmr12-18 | 57.6 b | 4.2 | 32.6 b | 2.7 | 2.5 c | 0.3 |
| (BTx623ms3 × bmr19)BC2F4 | bmr19-ref | 59.7 ab | 5.6 | 33.5 ab | 4.3 | 3.6 b | 0.2 |
| BTx623 EMS line 2354 | bmr19-2 | 62.2 ab | 3.5 | 34.8 ab | 2.5 | 3.7 b | 0.9 |
| Genotype * | Neutral Detergent Fiber (Cellulose, Hemicellulose and Lignin) | Acid Detergent Fiber (Cellulose and Lignin) | Acid Detergent Lignin (Lignin) | |||
|---|---|---|---|---|---|---|
| Mean † | Std Dev | Mean | Std Dev | Mean | Std Dev | |
| Bmr19 | 55.8 b | 1.7 | 29.1 b | 1.0 | 4.6 a | 0.2 |
| bmr19-ref | 56.1 b | 2.2 | 28.2 b | 1.2 | 3.9 c | 0.2 |
| bmr19-2 | 61.9 a | 1.8 | 31.8 a | 1.5 | 4.3 b | 0.2 |
| Genotye * | H (µg/g Dry Weight) | G (µg/g Dry Weight) | S (µg/g Dry Weight) | G/S | ||||
|---|---|---|---|---|---|---|---|---|
| Mean † | Std Dev | Mean | Std Dev | Mean | Std Dev | Mean | Std Dev | |
| Bmr19 | 6607.1 a | 852.3 | 169,109.5 a | 25,095.0 | 104,810.2 a | 21,144.7 | 1.6 a | 0.1 |
| bmr19-ref | 7629.6 b | 1134.5 | 126,173.5 c | 17,264.5 | 89,806.9 b | 16,331.9 | 1.4 b | 0.1 |
| bmr19-2 | 7239.5 ab | 951.1 | 141,872.7 b | 15,806.5 | 97,702.2 ab | 12,935.2 | 1.5 b | 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adeyanju, A.O.; Sattler, S.E.; Rich, P.J.; Rivera-Burgos, L.A.; Xu, X.; Ejeta, G. Sorghum Brown Midrib19 (Bmr19) Gene Links Lignin Biosynthesis to Folate Metabolism. Genes 2021, 12, 660. https://doi.org/10.3390/genes12050660
Adeyanju AO, Sattler SE, Rich PJ, Rivera-Burgos LA, Xu X, Ejeta G. Sorghum Brown Midrib19 (Bmr19) Gene Links Lignin Biosynthesis to Folate Metabolism. Genes. 2021; 12(5):660. https://doi.org/10.3390/genes12050660
Chicago/Turabian StyleAdeyanju, Adedayo O., Scott E. Sattler, Patrick J. Rich, Luis A. Rivera-Burgos, Xiaochen Xu, and Gebisa Ejeta. 2021. "Sorghum Brown Midrib19 (Bmr19) Gene Links Lignin Biosynthesis to Folate Metabolism" Genes 12, no. 5: 660. https://doi.org/10.3390/genes12050660
APA StyleAdeyanju, A. O., Sattler, S. E., Rich, P. J., Rivera-Burgos, L. A., Xu, X., & Ejeta, G. (2021). Sorghum Brown Midrib19 (Bmr19) Gene Links Lignin Biosynthesis to Folate Metabolism. Genes, 12(5), 660. https://doi.org/10.3390/genes12050660