
Identification, Phylogeny, Divergence, Structure, and Expression Analysis of A20/AN1 Zinc Finger Domain Containing Stress-Associated Proteins (SAPs) Genes in Jatropha curcas L.




Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification, Sequence Alignment, and Phylogenetic Analysis of SAP Genes in Jatropha Curcas
2.2. JcSAP Protein Physicochemical Characteristics
2.3. Gene Duplication Events, Homology and Synteny Analysis of JcSAP Genes
2.4. Conserved Domains, Motifs, and Gene Structure Organization of JcSAP Genes
2.5. Divergence Analysis
2.6. Protein Structure Analysis of JcSAP Genes
2.7. Cis-Elements Analysis and Predicted miRNA Target Sites
2.8. Gene Expression Profiling of JcSAP Genes
3. Results
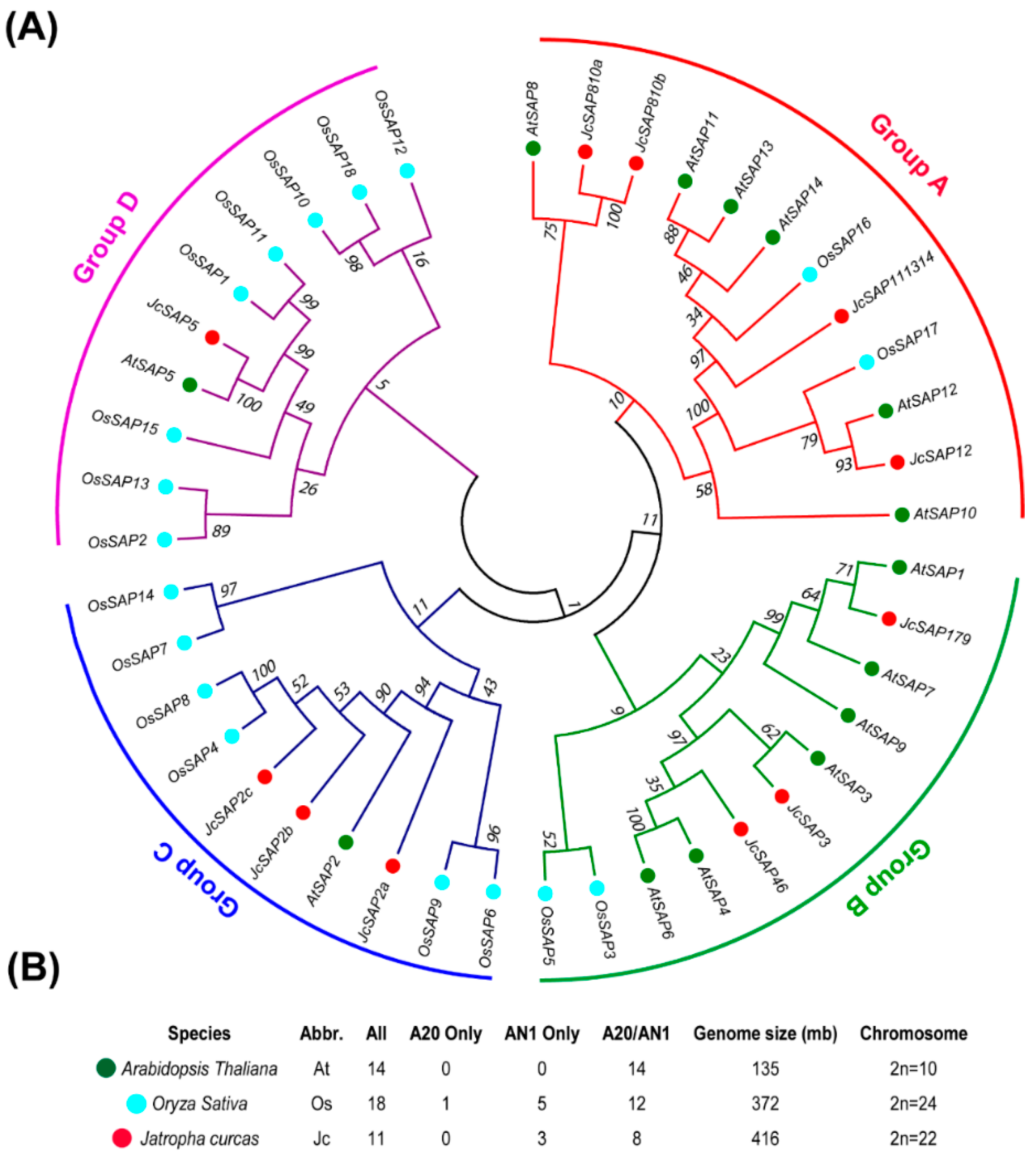
3.1. Identification and Phylogenetic Analysis of JcSAP Genes
3.2. Physicochemical Characteristics of JcSAP Genes
3.3. Synteny Analysis of JcSAP Genes
3.4. Determination of Non-Synonymous (Ka) and Synonymous (Ks) Substitution Rate
3.5. Gene Structure, Domain and Motif Analysis of JcSAP Genes
3.6. Protein Structure Analysis of JcSAP Genes
3.7. Promoter Region Analysis and Prediction of miRNA Target Sites of JcSAP Genes
3.8. Gene Expression Profiling of JcSAP Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wan, F.; Xu, Y.; Wang, S.; Gao, J.; Lu, D.; Zhou, C.; Liao, Y.; Ma, Y.; Zheng, Y. Identification and Expression Analysis of Zinc Finger A20/AN1 Stress-Associated Genes SmSAP Responding to Abiotic Stress in Eggplant. Horticulturae 2022, 8, 108. [Google Scholar] [CrossRef]
- Solanke, A.U.; Sharma, M.K.; Tyagi, A.K.; Sharma, A.K. Characterization and Phylogenetic Analysis of Environmental Stress-Responsive SAP Gene Family Encoding A20/AN1 Zinc Finger Proteins in Tomato. Mol. Genet. Genom. 2009, 282, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional Regulatory Networks in Cellular Responses and Tolerance to Dehydration and Cold Stresses. Annu. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef] [PubMed]
- Chinnusamy, V.; Zhu, J.; Zhu, J.-K. Cold Stress Regulation of Gene Expression in Plants. Trends Plant Sci. 2007, 12, 444–451. [Google Scholar] [CrossRef]
- Vij, S.; Tyagi, A.K. Emerging Trends in the Functional Genomics of the Abiotic Stress Response in Crop Plants. Plant Biotechnol. J. 2007, 5, 361–380. [Google Scholar] [CrossRef]
- Bhatnagar-Mathur, P.; Vadez, V.; Sharma, K.K. Transgenic Approaches for Abiotic Stress Tolerance in Plants: Retrospect and Prospects. Plant Cell Rep. 2008, 27, 411–424. [Google Scholar] [CrossRef]
- Solanke, A.U.; Sharma, A.K. Signal Transduction during Cold Stress in Plants. Physiol. Mol. Biol. Plants 2008, 14, 69–79. [Google Scholar] [CrossRef]
- Davletova, S.; Schlauch, K.; Coutu, J.; Mittler, R. The Zinc-Finger Protein Zat12 Plays a Central Role in Reactive Oxygen and Abiotic Stress Signaling in Arabidopsis. Plant Physiol. 2005, 139, 847–856. [Google Scholar] [CrossRef]
- Mukhopadhyay, A.; Vij, S.; Tyagi, A.K. Overexpression of a Zinc-Finger Protein Gene from Rice Confers Tolerance to Cold, Dehydration, and Salt Stress in Transgenic Tobacco. Proc. Natl. Acad. Sci. USA 2004, 101, 6309–6314. [Google Scholar] [CrossRef]
- Ciftci-Yilmaz, S.; Mittler, R. The Zinc Finger Network of Plants. Cell. Mol. Life Sci. 2008, 65, 1150–1160. [Google Scholar] [CrossRef]
- Xu, D.-Q.; Huang, J.; Guo, S.-Q.; Yang, X.; Bao, Y.-M.; Tang, H.-J.; Zhang, H.-S. Overexpression of a TFIIIA-Type Zinc Finger Protein Gene ZFP252 Enhances Drought and Salt Tolerance in Rice (Oryza Sativa L.). FEBS Lett. 2008, 582, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-Z.; Zheng, W.-J.; Cao, X.-Y.; Cui, X.-Y.; Zhao, S.-P.; Yu, T.-F.; Chen, J.; Zhou, Y.-B.; Chen, M.; Chai, S.-C. Genomic Analysis of Stress Associated Proteins in Soybean and the Role of GmSAP16 in Abiotic Stress Responses in Arabidopsis and Soybean. Front. Plant Sci. 2019, 10, 1453. [Google Scholar] [CrossRef]
- Baidyussen, A.; Aldammas, M.; Kurishbayev, A.; Myrzabaeva, M.; Zhubatkanov, A.; Sereda, G.; Porkhun, R.; Sereda, S.; Jatayev, S.; Langridge, P. Identification, Gene Expression and Genetic Polymorphism of Zinc Finger A20/AN1 Stress-Associated Genes, HvSAP, in Salt Stressed Barley from Kazakhstan. BMC Plant Biol. 2020, 20, 156. [Google Scholar] [CrossRef]
- He, X.; Xie, S.; Xie, P.; Yao, M.; Liu, W.; Qin, L.; Liu, Z.; Zheng, M.; Liu, H.; Guan, M. Genome-Wide Identification of Stress-Associated Proteins (SAP) with A20/AN1 Zinc Finger Domains Associated with Abiotic Stresses Responses in Brassica Napus. Environ. Exp. Bot. 2019, 165, 108–119. [Google Scholar] [CrossRef]
- Giri, J.; Dansana, P.K.; Kothari, K.S.; Sharma, G.; Vij, S.; Tyagi, A.K. SAPs as Novel Regulators of Abiotic Stress Response in Plants. Bioessays 2013, 35, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Long, L.; Tian, X.; Jin, J.; Liu, H.; Zhang, H.; Xu, F.; Song, C. Genome-Wide Identification and Expression Analysis of Stress-Associated Proteins (SAPs) Containing A20/AN1 Zinc Finger in Cotton. Mol. Genet. Genom. 2016, 291, 2199–2213. [Google Scholar] [CrossRef]
- Lai, W.; Zhou, Y.; Pan, R.; Liao, L.; He, J.; Liu, H.; Yang, Y.; Liu, S. Identification and Expression Analysis of Stress-Associated Proteins (SAPs) Containing A20/AN1 Zinc Finger in Cucumber. Plants 2020, 9, 400. [Google Scholar] [CrossRef] [PubMed]
- Fatima, S.; Zafar, Z.; Gul, A.; Bhatti, M.F. Genome-Wide Identification of Stress-Associated Proteins (SAPs) Encoding A20/AN1 Zinc Finger in Almond (Prunus Dulcis) and Their Differential Expression during Fruit Development. Plants 2021, 11, 117. [Google Scholar] [CrossRef]
- Fairless, D. The Little Shrub That Could–Maybe: India, like Many Countries, Has High Hopes for Jatropha as a Biofuel Source, but Little Is Known about How to Make It a Successful Crop. Daemon Fairless Digs for the Roots of a New Enthusiasm. Nature 2007, 449, 652–656. [Google Scholar] [CrossRef]
- Bhasanutra, R.; Sutiponpeibun, S. Jatropha Curcas Oil as a Substitute for Diesel Engine Oil. Int. Energy J. 2017, 4, 56–70. [Google Scholar]
- Openshaw, K. A Review of Jatropha Curcas: An Oil Plant of Unfulfilled Promise. Biomass Bioenergy 2000, 19, 1–15. [Google Scholar] [CrossRef]
- Dong, Y.; Lu, J.; Liu, J.; Jalal, A.; Wang, C. Genome-Wide Identification and Functional Analysis of JmjC Domain-Containing Genes in Flower Development of Rosa Chinensis. Plant Mol. Biol. 2020, 102, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, S.; Sun, J.; Sun, J.; Wang, H.; Cao, X.; Lu, J.; Jalal, A.; Wang, C. Genome-Wide Analysis Reveals Widespread Roles for RcREM Genes in Floral Organ Development in Rosa Chinensis. Genomics 2021, 113, 3881–3894. [Google Scholar] [CrossRef] [PubMed]
- Jalal, A.; Sun, J.; Chen, Y.; Fan, C.; Liu, J.; Wang, C. Evolutionary Analysis and Functional Identification of Clock-Associated PSEUDO-RESPONSE REGULATOR (PRRs) Genes in the Flowering Regulation of Roses. Int. J. Mol. Sci. 2022, 23, 7335. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ren, M.; Chen, H.; Wu, S.; Yan, H.; Jalal, A.; Wang, C. Evolution of SHORT VEGETATIVE PHASE (SVP) Genes in Rosaceae: Implications of Lineage-Specific Gene Duplication Events and Function Diversifications with Respect to Their Roles in Processes Other than Bud Dormancy. Plant Genome 2020, 13, e20053. [Google Scholar] [CrossRef]
- Ayaz, A.; Saqib, S.; Huang, H.; Zaman, W.; Lü, S.; Zhao, H. Genome-Wide Comparative Analysis of Long-Chain Acyl-CoA Synthetases (LACSs) Gene Family: A Focus on Identification, Evolution and Expression Profiling Related to Lipid Synthesis. Plant Physiol. Biochem. 2021, 161, 1–11. [Google Scholar] [CrossRef]
- Errum, A.; Rehman, N.; Khan, M.R.; Ali, G.M. Genome-Wide Characterization and Expression Analysis of Pseudo-Response Regulator Gene Family in Wheat. Mol. Biol. Rep. 2021, 48, 2411–2427. [Google Scholar] [CrossRef]
- Gaut, B.S.; Morton, B.R.; McCaig, B.C.; Clegg, M.T. Substitution Rate Comparisons between Grasses and Palms: Synonymous Rate Differences at the Nuclear Gene Adh Parallel Rate Differences at the Plastid Gene RbcL. Proc. Natl. Acad. Sci. USA 1996, 93, 10274–10279. [Google Scholar] [CrossRef]
- Ge, M.; Zhong, R.; Sadeghnezhad, E.; Hakeem, A.; Xiao, X.; Wang, P.; Fang, J. Genome-Wide Identification and Expression Analysis of Magnesium Transporter Gene Family in Grape (Vitis Vinifera). BMC Plant Biol. 2022, 22, 217. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, X.; Bai, H.; Liu, C. Genome-Wide Identification, Classification and Expression Analysis of the JmjC Domain-Containing Histone Demethylase Gene Family in Jatropha Curcas L. Sci. Rep. 2022, 12, 6543. [Google Scholar] [CrossRef]
- Ali, Q.; Ayaz, M.; Mu, G.; Hussain, A.; Yuanyuan, Q.; Yu, C.; Xu, Y.; Manghwar, H.; Gu, Q.; Wu, H. Revealing Plant Growth-Promoting Mechanisms of Bacillus Strains in Elevating Rice Growth and Its Interaction with Salt Stress. Front. Plant Sci. 2022, 13, 3190. [Google Scholar] [CrossRef] [PubMed]
- Sapeta, H.; Lourenço, T.; Lorenz, S.; Grumaz, C.; Kirstahler, P.; Barros, P.M.; Costa, J.M.; Sohn, K.; Oliveira, M.M. Transcriptomics and Physiological Analyses Reveal Co-Ordinated Alteration of Metabolic Pathways in Jatropha Curcas Drought Tolerance. J. Exp. Bot. 2016, 67, 845–860. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Wang, J.; Wang, W.; Hu, T.; Hu, H.; Bao, C. A High-Quality Chromosome-Level Genome Assembly Reveals Genetics for Important Traits in Eggplant. Hortic. Res. 2020, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- Rogozin, I.B.; Sverdlov, A.V.; Babenko, V.N.; Koonin, E.V. Analysis of Evolution of Exon-Intron Structure of Eukaryotic Genes. Brief. Bioinform. 2005, 6, 118–134. [Google Scholar] [CrossRef]
- Rose, A.B. Intron-Mediated Regulation of Gene Expression. Nucl. pre-mRNA Process. Plants 2008, 326, 277–290. [Google Scholar] [CrossRef]
- Dong, Q.; Duan, D.; Zhao, S.; Xu, B.; Luo, J.; Wang, Q.; Huang, D.; Liu, C.; Li, C.; Gong, X.; et al. Genome-Wide Analysis and Cloning of the Apple Stress-Associated Protein Gene Family Reveals MdSAP15, Which Confers Tolerance to Drought and Osmotic Stresses in Transgenic Arabidopsis. Int. J. Mol. Sci. 2018, 19, 2478. [Google Scholar] [CrossRef]
- Grzybowska, E.A. Human Intronless Genes: Functional Groups, Associated Diseases, Evolution, and MRNA Processing in Absence of Splicing. Biochem. Biophys. Res. Commun. 2012, 424, 1–6. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID (Uniprot) | Chr | AA | CDS | MW/kDa | PI | GRAVY | Formula | Predicted Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|
| JcSAP179 | A0A067JZZ9_JATCU | Un | 178 | 537 | 18834.23 | 8 | −0.453 | C809H1279N231O261S13 | Cytoplasmic |
| JcSAP2a | A0A067KTX6_JATCU | Un | 133 | 402 | 14686.65 | 9.1 | −0.612 | C630H990N188O194S12 | Cytoplasmic |
| JcSAP2b | A0A067JE11_JATCU | Un | 171 | 516 | 18507.08 | 8.5 | −0.442 | C785H1266N236O249S16 | Cytoplasmic |
| JcSAP2c | A0A067L4U9_JATCU | Un | 170 | 513 | 18087.44 | 8.8 | −0.304 | C761H1233N229O251S15 | Cytoplasmic |
| JcSAP3 | A0A067KRF9_JATCU | Un | 170 | 513 | 18661.18 | 8.8 | −0.492 | C793H1276N236O254S15 | Cytoplasmic |
| JcSAP46 | A0A067L6B6_JATCU | Un | 159 | 480 | 17559.01 | 8.6 | −0.521 | C754H1197N225O231S14 | Cytoplasmic |
| JcSAP5 | A0A067L4Q5_JATCU | Un | 185 | 558 | 20311.78 | 9.4 | −0.665 | C849H1395N271O283S12 | Cytoplasmic |
| JcSAP810a | A0A067JER8_JATCU | Un | 135 | 408 | 14879.04 | 9.3 | −0.49 | C643H1038N186O199S10 | Cytoplasmic |
| JcSAP810b | A0A067JQU7_JATCU | Un | 204 | 615 | 22711.95 | 9.3 | −0.775 | C972H1602N282O314S14 | Cytoplasmic |
| JcSAP111314 | A0A067K2L3_JATCU | Un | 288 | 867 | 31998.28 | 8.6 | −0.665 | C1367H2182N416O426S23 | Extracellular |
| JcSAP12 | A0A067LAI2_JATCU | Un | 197 | 594 | 21773.79 | 9 | −0.639 | C924H1487N285O288S18 | Extracellular |
| H (%) | T (%) | E (%) | RC (%) | |
|---|---|---|---|---|
| JcSAP11/13/14 | 22.22 | 1.04 | 11.11 | 65.62 |
| JcSAP12 | 13.2 | 2.54 | 13.71 | 70.56 |
| JcSAP1/7/9 | 21.35 | 2.81 | 11.8 | 64.04 |
| JcSAP3 | 27.65 | 3.53 | 11.76 | 57.06 |
| JcSAP2b | 25.15 | 4.68 | 10.53 | 59.65 |
| JcSAP4/6 | 30.19 | 4.4 | 12.58 | 52.83 |
| JcSAP2a | 26.32 | 4.51 | 13.53 | 55.64 |
| JcSAP5 | 37.84 | 2.7 | 10.27 | 49.19 |
| JcSAP8/10a | 14.07 | 3.7 | 13.33 | 68.89 |
| JcSAP2c | 28.82 | 4.12 | 11.76 | 55.29 |
| JcSAP8/10b | 21.08 | 2.94 | 13.24 | 62.75 |
| Paralogous Genes | Ka | Ks | Ka/Ks | T (MYA) | |
|---|---|---|---|---|---|
| JcSAP111314 | JcSAP12 | 0.2614 | 0.5275 | 0.495545 | 40.2 |
| JcSAP810b | JcSAP810a | 0.04609 | 0.11945 | 0.385851 | 9.1 |
| JcSAP2b | JcSAP2c | 0.12225 | 0.33925 | 0.360354 | 25.8 |
| JcSAP4/6 | JcSAP3 | 0.1221 | 0.4072 | 0.299853 | 31.0 |
| JcSAP5 | JcSAP179 | 0.26015 | 0.46745 | 0.55653 | 35.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jalal, A.; Ali, Q.; Manghwar, H.; Zhu, D. Identification, Phylogeny, Divergence, Structure, and Expression Analysis of A20/AN1 Zinc Finger Domain Containing Stress-Associated Proteins (SAPs) Genes in Jatropha curcas L. Genes 2022, 13, 1766. https://doi.org/10.3390/genes13101766
Jalal A, Ali Q, Manghwar H, Zhu D. Identification, Phylogeny, Divergence, Structure, and Expression Analysis of A20/AN1 Zinc Finger Domain Containing Stress-Associated Proteins (SAPs) Genes in Jatropha curcas L. Genes. 2022; 13(10):1766. https://doi.org/10.3390/genes13101766
Chicago/Turabian StyleJalal, Abdul, Qurban Ali, Hakim Manghwar, and Daochen Zhu. 2022. "Identification, Phylogeny, Divergence, Structure, and Expression Analysis of A20/AN1 Zinc Finger Domain Containing Stress-Associated Proteins (SAPs) Genes in Jatropha curcas L." Genes 13, no. 10: 1766. https://doi.org/10.3390/genes13101766
APA StyleJalal, A., Ali, Q., Manghwar, H., & Zhu, D. (2022). Identification, Phylogeny, Divergence, Structure, and Expression Analysis of A20/AN1 Zinc Finger Domain Containing Stress-Associated Proteins (SAPs) Genes in Jatropha curcas L. Genes, 13(10), 1766. https://doi.org/10.3390/genes13101766