

The Roles of SNHG Family in Osteoblast Differentiation
Department of Orthodontics, Peking University School and Hospital of Stomatology, National Center of Stomatology, National Clinical Research Center for Oral Diseases, National Engineering Laboratory for Digital and Material Technology of Stomatology, No. 22, Zhongguancun South Avenue, Haidian District, Beijing 100081, China
*
Author to whom correspondence should be addressed.
Genes 2022, 13(12), 2268; https://doi.org/10.3390/genes13122268
Submission received: 10 October 2022
/
Revised: 17 November 2022
/
Accepted: 28 November 2022
/
Published: 2 December 2022
(This article belongs to the Special Issue Genetics Studies of Bone Disease)
Abstract
:Small nucleolar RNA host genes (SNHGs), members of long-chain noncoding RNAs (lncRNAs), have received increasing attention regarding their roles in multiple bone diseases. Studies have revealed that SNHGs display unique expression profile during osteoblast differentiation and that they could act as promising biomarkers of certain bone diseases, such as osteoporosis. Osteogenesis of mesenchymal stem cells (MSCs) is an important part of bone repair and reconstruction. Moreover, studies confirmed that the SNHG family participate in the regulation of osteogenic differentiation of MSCs in part by regulating important pathways of osteogenesis, such as Wnt/β-catenin signaling. Based on these observations, clarifying the SNHG family’s roles in osteogenesis (especially in MSCs) and their related mechanisms would provide novel ideas for possible applications of lncRNAs in the diagnosis and treatment of bone diseases. After searching, screening, browsing and intensive reading, we uncovered more than 30 papers related to the SNHG family and osteoblast differentiation that were published in recent years. Here, our review aims to summarize these findings in order to provide a theoretical basis for further research.
1. Introduction
Disruption of bone remodeling cycles is the foundation of many bone diseases, including osteoporosis [1]. In the process of bone remodeling, mesenchymal stem cells (MSCs) differentiate into osteoblasts, which then mature into osteocytes. This ability enables MSCs to participate in bone reconstruction and possess greater bone repair application potential than other cell types [2]. Small nucleolar RNAs (snoRNAs), members of noncoding RNAs, mainly exist in nucleolus and consist of about 60-300 nucleotides [3]. Our review focuses on small nucleolar RNA host genes (SNHGs), which are host genes of snoRNAs. In humans, more than half of snoRNAs are transcribed from host genes. Among these host genes, SNHGs account for about four-fifths [4]. Currently, the SNHG family has been reported to have more than 20 members [5].
SNHGs’ aberrant expression is reflected in bone disorders. It was observed that in a postmenopausal osteoporotic mouse model (OVX mice), compared with sham mice, SNHG1 expression in serum and femoral tissue was obviously increased [6]. In serum of osteoporosis patients, SNHG14 expression was greatly elevated [7]. In blood mononuclear cells (MNCs) from osteoporosis patients, differentiation antagonizing non-protein coding RNA (DANCR) was upregulated [8]. Bone marrow-derived stem cells (BMSCs) from osteoporosis patients were derived, and researchers observed a relatively high expression of DANCR [9]. Centofanti et al. observed aberrantly expressed long-chain noncoding RNAs (lncRNAs) in osteoblast primary cells from an osteoporosis group and control group. Among them, growth arrest specific 5 (GAS5) was one of the six lncRNAs with the most significant downregulation in the osteoporosis group [10].
The above findings suggest that the SNHG family may play a role in bone repair. In fact, studies regarding SNHG members and osteogenesis have increased significantly in recent years. Among SNHG family members, SNHG1, SNHG2 (alias GAS5), SNHG5, SNHG7, SNHG14 and SNHG13 (alias DANCR) have been demonstrated to be involved in osteogenesis, and most of these studies focused on the relationship between SNHG and MSC osteoblast differentiation. Due to the complexity of their biological functions and related mechanisms, the SNHG family’s roles in osteogenesis have not been clearly expounded. Therefore, we try to review these functions and mechanisms here for a better summarization.
2. SNHGs Display Aberrant Expression in Osteoblast Differentiation
Numerous studies have indicated that SNHGs are differentially expressed during osteoblast differentiation [6,11,12,13]. During osteogenesis induction of bone marrow mesenchymal stem cells (BMSCs), with the notable enhancement of alkaline phosphatase (ALP) activity, SNHG1 was markedly downregulated [6]. Another study also provided evidence that SNHG1 was downregulated in a time-dependent manner during osteogenic induction of BMSCs [13]. The above results unanimously indicate that SNHG1 may be involved in osteogenic differentiation of BMSCs. Other SNHG family members also participate in osteoblast differentiation. It was observed that expression of SNHG5 was upregulated following prolongation of BMSC osteogenic induction [11]. SNHG7 expression was significantly reduced during osteogenesis of TNF-α-treated human dental pulp stem cells (hDPSCs). Moreover, the amplitude of SNHG7 expression variation was related to the concentration of TNF-α in the experimental treatment [12].
More studies have focused on DANCR (alias SNHG13). Based on previous results showing that human amnion-derived mesenchymal stem cells (HAMSCs) may have the ability to boost osteoblast differentiation of BMSCs [14], researchers demonstrated that when BMSCs were co-cultured with HAMSCs, expression of DANCR was inhibited. Furthermore, DANCR’s downregulation may act as an intermediate process of HAMSCs’ promoting effect on BMSC osteogenesis [15]. In previous studies, which indicated that DANCR might influence the pathological process of osteoporosis [8], researchers assessed the expression of DANCR during osteoblast induction of BMSCs, and results indicated that the expression of DANCR gradually decreased [16].
Our team has performed a series of studies on SNHG members [17,18]. We found that GAS5 displayed a gradual upregulation in hDPSCs during 14-day osteoblast induction [17]. Moreover, in our research on another SNHG family member, SNHG8, we found that SNHG8 may be involved in periodontal ligament stem cells (PDLSCs) osteogenic differentiation. Osteogenesis of PDLSCs was induced for two days, and we found that the induction not only increased the expression of osteogenic markers but also promoted expression of SNHG8 (Figure 1).
GAS5 has also received attention from other researchers. It was reported that GAS5 expression exhibited an upward trend during osteogenic differentiation of BMSCs [19]. Furthermore, studies have observed that GAS5 had 15 transcript variants. Although some variants’ expression was promoted during BMSC osteogenesis, the GAS5 transcript variant 2 displayed an opposite trend [20].
To summarize, the studies cited above indicated that SHNGs exhibited different expression levels during osteoblast differentiation. During BMSC osteoblast differentiation, SNHG1 and DANCR (alias SNHG13) tended to be downregulated, while SNHG5 exhibited an opposite trend. During DPSC osteoblast differentiation, SNHG7 tended to decrease. GAS5 (alias SNHG2) presented a more complicated expression trend during osteoblast differentiation. More details are shown in tabular form in Table 1. These results signify that SNHGs might play a role in osteoblast differentiation.
3. The Mechanism of SNHGs Regulating Osteoblast Differentiation
3.1. SNHG1
SNHG1, small nucleolar RNA host gene 1, is located on chromosome 11q12.3 in humans and includes 11 exons [21,22]. Researchers observed that SNHG1 served as molecular sponge for miR-181c-5p and decrease the level of miR-181c-5p [6]. Previous research reported that miR-181c-5p might act as biomarker of osteoporosis [23]. By targeting miR-181c-5p, silencing of SNHG1 enhanced osteoblast differentiation of BMSCs, while overexpression of SNHG1 promoted osteoclast formation of BMSCs. The possible downstream molecular target of miR-181c-5p was SFRP1, which regulates the Wnt3a signaling pathway. This research discovered that SNHG1 overexpression promoted SFRP1 expression but inhibited Wnt3a, while miR-181c-5p overexpression significantly reversed these changes. Altogether, SNHG1 modulated the SFRP1/Wnt3a pathway via directly binding to miR-181c-5p and finally produced inhibiting effects on BMSC osteogenic differentiation [6]. Jiang et al. [24] also studied SNHG1’s role in BMSC osteogenesis. They observed that SNHG1 inhibited p38 activation via Nedd4, which is a member of the E3 family (key enzymes of ubiquitination) [25,26,27]. MAPK signaling pathways, including p38, play important roles in osteoblast differentiation. The inactivity of p38 could induce disturbance of osteoblast differentiation [28,29]. Thus, SNHG1 decreased p38 activity via ubiquitination, which was mediated by Nedd4, and then impeded osteoblast differentiation of BMSCs [24]. Another study in relation to BMSC osteoblast differentiation focused on Wnt/β-catenin signaling, which impeded osteoclast formation [30,31]. It was confirmed that Dickkopf 1 (DKK1) was a Wnt signaling inhibitor [32]. Via bioinformatics analysis, they found that both SNHG1 and DKK1 interacted with miR-101. Later results indicated that SNHG1 could suppress BMSC osteoblast differentiation by serving as competitive endogenous RNA (ceRNA) for miR-101. In this process, DKK1 was the key downstream molecular target. In short, SNHG1 modulated Wnt/β-catenin signal pathway though the miR-101/DKK1 axis and finally impeded BMSC osteoblast differentiation [13].
MC3T3-E1, a pre-osteoblast cell line, is commonly used for studies regarding osteoblast induction. It was reported that during osteoblast induction of MC3T3-E1 cells, SNHG1 played a negative role. In this process, SNHG1 acted as a sponge for miR-181a-5p. Moreover, by means of online bioinformatics prediction, PTEN was recognized as the target of miR-181a-5p, and PTEN might participate in this modulatory axis. Overall, this study revealed that the SNHG1/miR-181a-5p/PTEN axis had an inhibitory effect on osteogenesis of MC3T3-E1 cells [33].
SNHG1’s role in osteoblast differentiation of MSCs, especially BMSCs, has been summarized thus far. In general, SNHG1 exerts a negative effect on osteogenic differentiation. This result may help to better understand SNHG1-related mechanisms of bone diseases.
3.2. GAS5 (Alias SNHG2)
GAS5, growth arrest specific 5 (alias SNHG2), was overexpressed in growth arrest cells, hence the name growth arrest specific 5 [34]. Our team observed that silencing GAS5 impeded osteogenic differentiation of PDLSCs, and the regulatory effect of GAS5 was mediated through the p38/JNK pathway, which was an important pathway related to bone formation [17]. Wang et al. [19] showed that in BMSCs derived from ovariectomized (OVX) animal models, expression of GAS5 was promoted, which indicated that GAS5 might be involved in BMSC osteogenesis. Later experiments indicated that GAS5 acted as a sponge for miR-135a-5p and decrease its level. The downstream target of miR-135a-5p was FOXO1, which could significantly promote osteogenic differentiation [35]. In BMSCs derived from OVX-induced osteoporotic mice, GAS5 functioned as sponge for miR-135a-5p to positively modulate FOXO1 expression and finally increase osteogenesis [19]. Another study also investigated GAS5’s role in BMSC osteogenesis. Researchers found that GAS5 levels were decreased in BMSCs and bone tissues of osteoporosis patients. Furthermore, GAS5 was positively related to osteogenesis differentiation. Later studies showed that GAS5 functioned via specifically combining with UPF1 protein. The result of this combination was the acceleration of SMAD7’s decay, while SMAD7 was an inhibitor of SMAD1/5/8 [36]. In short, GAS5 increased osteogenesis differentiation via the UPF1/smad7 axis and then increased bone repair activity [37] (Figure 2).
LncRNAs can generate different transcript variants. Variants from the same lncRNA might display different expression profiles in diseases [38,39]. Song et al. [20] reported 15 transcript variants of GAS5. They focused on GAS5 transcript variant 2 and found that it inhibited osteoblast differentiation of hBMSCs. This result was contrary to the findings of previous researches regarding GAS5’s roles in osteoblast differentiation of BMSCs. They further studied the mechanism underlying their findings and discovered that GAS5 (transcript variant 2) targeted miR-382-3p, which could modulate TAF1 expression. TAF1 inhibited osteogenesis. Moreover, TAF1 promoted GAS5 expression, indicating that they formed a positive GAS5/miR-382-3p/TAF1 feedback loop in hBMSCs [20].
In summary, in osteogenesis of PDLSCs and BMSCs, GAS5 usually plays a positive role, while GAS5 (transcript variant 2) does the opposite. More studies are necessary for a deeper understanding of GAS5 variants’ regulatory effects in osteoblast differentiation.
3.3. SNHG5
SNHG5, small nucleolar RNA host gene 5, is 524 base pairs (bp) in length. In humans, it is located on chromosome 6q14.3 [18,40]. It was observed that knockdown of SNHG5 impeded hBMSCs osteoblast differentiation. The mechanism underlying this phenomenon was related to miR-582-5p. SNHG5 served as sponge for miR-582-5p. As a result, SNHG5 modulated the expression of RUNX3. RUNX3 belongs to the RUNX family and has been confirmed to function as an inducer of osteogenic differentiation. Mechanically, SNHG5 served as sponge for miR-582-5p and then modulated RUNX3 expression to promote hBMSC osteogenic differentiation. Moreover, RUNX3 also promoted activity of SNHG5 at the level of transcription. In this way, a positive feedback loop of SNHG5/miR-582-5p/RUNX3 was formed [11].
Our team also studied the positive role of SNHG5 in the osteogenesis and identified a regulatory axis that modulates hBMSC osteogenic differentiation: YY1/SNHG5/miR-212-3P/GDF5/Smad. Through this axis, promoting SNHG5 expression stimulated hBMSC osteogenic differentiation, which might provide a potential target for bone repair [18]. It is worth mentioning that in another study on periodontitis, our team found that local si-SNHG5 injection into mouse periodontitis model resulted in severe bone loss [41]. The positive role of SNHG5 in bone formation was further demonstrated by this finding.
3.4. SNHG7
SNHG7, small nucleolar RNA host gene 7, is 2176 bp in length. In humans, SNHG7 is located in the chromosome 9q34.3 [42,43]. It was reported that in the pre-osteoblast cell line, MC3T3-E1, SNHG7 functioned as sponge for miR-9, while miR-9 targeted the 3’UTR of TGFBR2. Further study revealed that knockdown of SNHG7 inhibited the TGF-β signaling pathway. TGF-β has been confirmed to act as significant regulator of osteoblast and osteoclast differentiation [44]. These results showed that knockdown of SNHG7 resulted in lower osteogenic activation via the miR-9/TGF-β axis [45]. Additionally, in TNF-α-treated hDPSCs, research reported that SNHG7 exerted a positive effect on osteoblast differentiation [12]. In this research, hDPSCs were treated with 50 ng/mL of TNF-α to simulate an inflammatory microenvironment, and these cells then underwent osteogenic induction. Researchers observed that in hDPSCs, promoting expression of SNHG7 reversed the inhibiting influence of TNF-α on osteoblast/dentinogenic differentiation. Additionally, miR-6512-3p participated in this process via interacting with SNHG7. In short, SNHG7 promoted osteoblast differentiation of hDPSCs treated with TNF-α via targeting miR-6512-3p [12]. Another study also showed that silencing of SNHG7 impeded the odontogenic and osteogenic activity of hDPSCs [46]. Researchers identified aberrantly expressed mRNAs, lncRNAs and microRNAs during osteo/odontogenic induction of DPSCs. Among them, SNHG7 was one of the most upregulated. To verify SNHG7’s role in osteo/odontogenic induction of DPSCs, SNHG7 silencing was conducted, and as a result, the expression of mineralization and osteogenic marker genes in DPSCs was inhibited [46]. These results are consistent with the positive role of SNHG7 in osteogenesis shown in previous studies.
3.5. DANCR (Alias SNHG13)
DANCR, differentiation antagonizing non-protein coding RNA, is found on chromosome 4q12 [47]. As a member of lncRNA family, DANCR participates in numerous diseases and pathological process, such as tumors and bone diseases [48,49]. There have been an increasing number of studies focusing on the relationship between DANCR and osteoblast differentiation regulation. For example, Tang et al. [50] showed that DANCR participated in osteolysis after total hip arthroplasty, and in a subsequent experiment, they found that DANCR could decrease MSCs’ osteogenesis activity via modulating FOXO1, which is a transcription factor (Figure 3).
Additional studies have focused on the relationship between DANCR and BMSC differentiation. Previous studies have confirmed that human amnion-derived mesenchymal stem cells (HAMSCs) had the ability to stimulate osteoblast differentiation of hBMSCs [51]. To explore the mechanism, researchers focused on the differentially expressed lncRNAs in hBMSCs co-cultured with HAMSCs. Among these differentially expressed genes, DANCR attracted attention of researchers. They observed that promoting DANCR expression impeded osteoblast differentiation of hBMSCs in the co-culture system [15]. Another study also showed that DANCR exerted inhibitory effects on hBMSC osteoblast differentiation. It was reported that DANCR modulated hBMSC osteoblast differentiation by absorbing miR-1301-3p. Moreover, they confirmed that PROX1, a transcription factor, was a downstream target of miR-1301-3p. In summary, this study showed that DANCR inhibited hBMSC osteogenesis via the DANCR/miR-1301-3p/PROX1 axis [16]. Additionally, it was noticed that in BMSCs derived from osteoporosis patients, DANCR expression was decreased during osteoblast induction. This research first verified the interaction between DANCR, miR-320a and CTNNB1 during osteogenesis. It was demonstrated that DANCR and miR-320a did not interact with each other in terms of expression. CTNNB1, the gene that encodes β-catenin, was directly modulated by miR-320a. In short, this study verified a new network composed of DANCR, miR-320a and CTNNB1 through the Wnt/β-catenin pathway during hBMSC osteoblast differentiation [9].
In MC3T3-E1, a pre-osteoblast cell line, DANCR also attracted researchers’ attention. After silencing DANCR in MC3T3-E1 cells, researchers observed an increasing trend of osteoblast differentiation. Further study revealed that the Wnt/β-catenin pathway also participated in this effect, which is consistent with abovementioned results [52].
Additionally, effects of nutrients on osteogenesis also related to DANCR. Yang et al. [47] reported that sesamin displayed a dual function in both osteogenesis promotion and osteoclastogenesis inhibition. Sesamin increased osteogenesis by activating the Wnt/β-catenin pathway and decreased osteoclastogenesis via deactivating the NF-κB signaling pathway. Previous studies confirmed that deactivating the NF-κB signaling pathway inhibited osteoclast formation [53]. Further studies revealed that sesamin’s promoting effects on osteoblast were attributable to downregulation of DANCR expression. In other words, DANCR exhibited negative effects on osteoblasts, which was consistent with the findings of previous studies [9,16,54].
3.6. SNHG14
SNHG14, small nucleolar RNA host gene 14, has been scarcely reported in relation to its effects on the differentiation of MSCs. In humans, SNHG14 is located on chromosome 15q11.2 [55]. Researchers noticed that SNHG14 may show disordered expression during human mesenchymal stem cells (hMSCs) differentiation. Further studies showed that silencing of SNHG14 inhibited osteogenesis of hMSCs via the miR-2861/AKT2 axis. More specifically, the promoting effect of SNHG14 on hMSC osteogenesis could be reversed by miR-2861, and the downstream target of miR-2861 was AKT2 [7]. Consequently, promoting SNHG14 expression would ultimately stimulate hMSC osteoblast differentiation.
4. Prospect
In recent years, SNHGs are gaining increased attention in the epigenetics of bone remodeling. Researchers in related fields focus on relationships between the SNHG family and osteogenesis, and the purpose of this review was to summarize findings of this research field (Table 2).
Our team has found that the SNHG family not only plays a role in osteogenesis but also participates in the progression of inflammation. For example, we found that SNHG5 mediated periodontal inflammation via the NF-κB signaling pathway [41]. Another SNHG family member, SNHG8, also affected the NF-κB signaling pathway and inflammatory processes [53]. We observed that in a hypoxic inflammatory microenvironment, decreased SNHG8 expression activated the downstream NF-κB signaling pathway. This indicated that SNHG8 participated in periodontal tissue reconstruction, which could be related to the bone remodeling discussed in this review. It can be considered that the SNHG family not only affects bone reconstruction through osteogenesis but also through its effects on inflammation. This conclusion provides novel insights for exploring more functions of the SNHG family.
Understanding of the SNHG family’s regulatory network in osteoblast differentiation remains to be improved. Most of studies mentioned above were based on ceRNA (competing endogenous RNA) mechanisms. SNHGs could act as ceRNAs and competitively bind to specific microRNAs. Downstream, microRNA mediates the silencing of specific genes [56]. However, in several studies mentioned in this review, SNHGs do not function through ceRNA mechanisms but directly bind to proteins (Figure 2 and Figure 3). This suggests that the ceRNA mechanism involved in SNHG function may have a parallel protein mechanism. Moreover, there may be an intersection between these two kinds of mechanisms. In general, SNHGs’ roles in osteogenesis tend to be diverse, and more work needs to be conducted to establish definitive mechanisms through which SNHGs modulate osteoblast differentiation.
5. Conclusions
In this review, we first summarized SNHGs that reportedly participate in osteogenic differentiation, especially in MSCs. To summarize, SNHG1 and SNHG 13 (alias DANCR) have negative effects on osteogenesis, while SNHG5, SNHG7 and SNHG14 can promote osteogenesis. Except the inhibitory effect of GAS5 transcript variant 2 on osteogenesis, most studies of GAS5 (alias SNHG2) show that it can promote osteogenesis.
In addition, we discussed their molecular regulatory mechanisms, which can be divided into two types: lncRNA-miRNA-mRNA crosstalk and direct interaction with protein. In the future, researchers may reveal more molecules which are involved in the SNHG family’s regulatory network and further verify the SNHG family’s effects on osteoblast differentiation.
Regarding the treatment of bone diseases, for the needs of bone regeneration, MSCs have always been the focus of researchers, and MSC implantation has shown its clinical value in the therapy of osteoporosis [57]. Moreover, adding specific growth factors could improve the treatment effect [58]. Most of studies on SNHGs and osteogenesis were carried out in MSCs, and the effects of SNHGs on MSC osteogenesis could have potential to be applied in clinic. For example, based on the existing MSC implantation therapy, it could be considered to use SNHGs’ effects at the same time to increase the therapeutic effect. We expect that the above findings regarding the SNHG family and osteogenesis will eventually guide researchers to find new treatments for bone diseases.
Author Contributions
Writing—original draft preparation, A.-Q.T.; writing—review and editing, Y.-F.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors have no conflicts of interest to declare.
References
- Shibli, J.A.; Nagay, B.E.; Suárez, L.J.; Urdániga Hung, C.; Bertolini, M.; Barão, V.A.R.; Souza, J.G.S. Bone Tissue Engineering Using Osteogenic Cells: From the Bench to the Clinical Application. Tissue Eng. Part C Methods 2022, 28, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, G.; Moncrieff, L.; Dompe, C.; Janowicz, K.; Sibiak, R.; Bryja, A.; Jankowski, M.; Mozdziak, P.; Bukowska, D.; Antosik, P.; et al. Bone Regeneration, Reconstruction and Use of Osteogenic Cells; from Basic Knowledge, Animal Models to Clinical Trials. J. Clin. Med. 2020, 9, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, P.; Ghosh, S.; Wang, B.; Heyns, M.; Graham, K.; Mackey, J.R.; Kovalchuk, O.; Damaraju, S. Profiling of Small Nucleolar RNAs by Next Generation Sequencing: Potential New Players for Breast Cancer Prognosis. PLoS ONE 2016, 11, e0162622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, J.; Li, Y.; Liu, C.; Xiang, Y.; Li, C.; Ye, Y.; Zhang, Z.; Hawke, D.H.; Park, P.K.; Diao, L.; et al. A Pan-Cancer Analysis of the Expression and Clinical Relevance of Small Nucleolar RNAs in Human Cancer. Cell Rep. 2017, 21, 1968–1981. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y.; Sun, W.; Wang, Z.; Dong, W.; He, L.; Zhang, T.; Zhang, H. Long Non-Coding Small Nucleolar RNA Host Genes (SNHGs) in Endocrine-Related Cancers. Onco. Targets Ther. 2020, 13, 7699–7717. [Google Scholar] [CrossRef]
- Yu, X.; Rong, P.-Z.; Song, M.-S.; Shi, Z.-W.; Feng, G.; Chen, X.-J.; Shi, L.; Wang, C.-H.; Pang, Q.-J. LncRNA SNHG1 Induced by SP1 Regulates Bone Remodeling and Angiogenesis via Sponging MiR-181c-5p and Modulating SFRP1/Wnt Signaling Pathway. Mol. Med. 2021, 27, 141. [Google Scholar] [CrossRef]
- Du, M.; Wu, B.; Fan, S.; Liu, Y.; Ma, X.; Fu, X. SNHG14 Induces Osteogenic Differentiation of Human Stromal (Mesenchymal) Stem Cells in Vitro by Downregulating MiR-2861. BMC Musculoskelet. Disord. 2020, 21, 525. [Google Scholar] [CrossRef]
- Tong, X.; Gu, P.; Xu, S.; Lin, X. Long Non-Coding RNA-DANCR in Human Circulating Monocytes: A Potential Biomarker Associated with Postmenopausal Osteoporosis. Biosci. Biotechnol. Biochem. 2015, 79, 732–737. [Google Scholar] [CrossRef]
- Wang, C.-G.; Hu, Y.-H.; Su, S.-L.; Zhong, D. LncRNA DANCR and MiR-320a Suppressed Osteogenic Differentiation in Osteoporosis by Directly Inhibiting the Wnt/β-Catenin Signaling Pathway. Exp. Mol. Med. 2020, 52, 1310–1325. [Google Scholar] [CrossRef]
- Centofanti, F.; Santoro, M.; Marini, M.; Visconti, V.V.; Rinaldi, A.M.; Celi, M.; D’Arcangelo, G.; Novelli, G.; Orlandi, A.; Tancredi, V.; et al. Identification of Aberrantly-Expressed Long Non-Coding RNAs in Osteoblastic Cells from Osteoporotic Patients. Biomedicines 2020, 8, 65. [Google Scholar] [CrossRef]
- Zheng, J.; Guo, H.; Qin, Y.; Liu, Z.; Ding, Z.; Zhang, L.; Wang, W. SNHG5/MiR-582-5p/RUNX3 Feedback Loop Regulates Osteogenic Differentiation and Apoptosis of Bone Marrow Mesenchymal Stem Cells. J. Cell. Physiol. 2020. Published online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Chen, W. SNHG7 Promotes the Osteo/Dentinogenic Differentiation Ability of Human Dental Pulp Stem Cells by Interacting with Hsa-MiR-6512–3p in an Inflammatory Microenvironment. Biochem. Biophys. Res. Commun. 2021, 581, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Fu, H.; Xu, Z.; Fan, W.; Liu, F.; Chen, B. LncRNA SNHG1 Attenuates Osteogenic Differentiation via the MiR-101/DKK1 Axis in Bone Marrow Mesenchymal Stem Cells. Mol. Med. Rep. 2020, 22, 3715–3722. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jiang, F.; Liang, Y.; Shen, M.; Chen, N. Human Amnion-Derived Mesenchymal Stem Cells Promote Osteogenic Differentiation in Human Bone Marrow Mesenchymal Stem Cells by Influencing the ERK1/2 Signaling Pathway. Stem Cells Int. 2016, 2016, 4851081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Miao, J.; Meng, X.; Chen, N.; Wang, Y. Expression of Long Non-Coding RNAs in Human Bone Marrow Mesenchymal Stem Cells Co-Cultured with Human Amnion-Derived Mesenchymal Stem Cells. Mol. Med. Rep. 2017, 16, 6683–6689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, W.; Di, S.; Xing, S.; Sun, Z.; Shen, Z.; Dou, X.; He, S.; Tang, H.; Min, J. Long Non-Coding RNA DANCR Modulates Osteogenic Differentiation by Regulating the MiR-1301-3p/PROX1 Axis. Mol. Cell Biochem. 2021, 476, 2503–2512. [Google Scholar] [CrossRef]
- Yang, Q.; Han, Y.; Liu, P.; Huang, Y.; Li, X.; Jia, L.; Zheng, Y.; Li, W. Long Noncoding RNA GAS5 Promotes Osteogenic Differentiation of Human Periodontal Ligament Stem Cells by Regulating GDF5 and P38/JNK Signaling Pathway. Front. Pharmacol. 2020, 11, 701. [Google Scholar] [CrossRef]
- Han, Y.; Yang, Q.; Huang, Y.; Jia, L.; Zheng, Y.; Li, W. Long Non-Coding RNA SNHG5 Promotes the Osteogenic Differentiation of Bone Marrow Mesenchymal Stem Cells via the MiR-212-3p/GDF5/SMAD Pathway. Stem. Cell Res. Ther. 2022, 13, 130. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, D.; Zhu, Y.; Dong, Y.; Liu, Y. Long Non-Coding RNA GAS5 Promotes Osteogenic Differentiation of Bone Marrow Mesenchymal Stem Cells by Regulating the MiR-135a-5p/FOXO1 Pathway. Mol. Cell. Endocrinol. 2019, 496, 110534. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, H.; Song, Z.; Yang, Y.; Zhang, S.; Wang, W.; Zhang, S. Long Noncoding RNA GAS5 Inhibits Osteogenic Differentiation through MicroRNA 382-3p/TAF1 Signaling. Mol. Cell Biol. 2022, 42, e00541-20. [Google Scholar] [CrossRef]
- Tian, M.; Gong, W.; Guo, J. Long Non-Coding RNA SNHG1 Indicates Poor Prognosis and Facilitates Disease Progression in Acute Myeloid Leukemia. Biol. Open 2019, 8, bio046417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long Non-Coding RNA SNHG1 Activates Glycolysis to Promote Hepatocellular Cancer Progression through the miR-326/PKM2 Axis. Available online: https://onlinelibrary.wiley.com/doi/epdf/10.1002/jgm.3440 (accessed on 31 August 2022).
- Ma, J.; Lin, X.; Chen, C.; Li, S.; Zhang, S.; Chen, Z.; Li, D.; Zhao, F.; Yang, C.; Yin, C.; et al. Circulating MiR-181c-5p and MiR-497-5p Are Potential Biomarkers for Prognosis and Diagnosis of Osteoporosis. J. Clin. Endocrinol. Metab. 2020, 105, 1445–1460. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wu, W.; Jiao, G.; Chen, Y.; Liu, H. LncRNA SNHG1 Modulates P38 MAPK Pathway through Nedd4 and Thus Inhibits Osteogenic Differentiation of Bone Marrow Mesenchymal Stem Cells. Life Sci. 2019, 228, 208–214. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, S.K.; Xiao, K.; Venkataramanan, V.; Snyder, P.M.; Freedman, N.J.; Weissman, A.M. Nedd4 Mediates Agonist-Dependent Ubiquitination, Lysosomal Targeting, and Degradation of the Β2-Adrenergic Receptor. J. Biol. Chem. 2008, 283, 22166–22176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, A.; Hou, Q.; Jarzylo, L.; Amato, S.; Gilbert, J.; Shang, F.; Man, H.-Y. Nedd4-Mediated AMPA Receptor Ubiquitination Regulates Receptor Turnover and Trafficking. J. Neurochem. 2011, 119, 27–39. [Google Scholar] [CrossRef]
- Grimsey, N.J.; Narala, R.; Rada, C.C.; Mehta, S.; Stephens, B.S.; Kufareva, I.; Lapek, J.; Gonzalez, D.J.; Handel, T.M.; Zhang, J.; et al. A Tyrosine Switch on NEDD4-2 E3 Ligase Transmits GPCR Inflammatory Signaling. Cell Rep. 2018, 24, 3312–3323.e5. [Google Scholar] [CrossRef] [Green Version]
- Robaszkiewicz, A.; Valkó, Z.; Kovács, K.; Hegedűs, C.; Bakondi, E.; Bai, P.; Virág, L. The Role of P38 Signaling and Poly(ADP-Ribosyl)Ation-Induced Metabolic Collapse in the Osteogenic Differentiation-Coupled Cell Death Pathway. Free Radic. Biol. Med. 2014, 76, 69–79. [Google Scholar] [CrossRef] [Green Version]
- Ba, P.; Duan, X.; Fu, G.; Lv, S.; Yang, P.; Sun, Q. Differential Effects of P38 and Erk1/2 on the Chondrogenic and Osteogenic Differentiation of Dental Pulp Stem Cells. Mol. Med. Rep. 2017, 16, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Yavropoulou, M.; Yovos, J. The Role of the Wnt Signaling Pathway in Osteoblast Commitment and Differentiation. Hormones 2007, 6, 279–294. [Google Scholar] [CrossRef]
- Albers, J.; Keller, J.; Baranowsky, A.; Beil, F.T.; Catala-Lehnen, P.; Schulze, J.; Amling, M.; Schinke, T. Canonical Wnt Signaling Inhibits Osteoclastogenesis Independent of Osteoprotegerin. J. Cell Biol. 2013, 200, 537–549. [Google Scholar] [CrossRef]
- Li, Y.; Lu, W.; King, T.D.; Liu, C.-C.; Bijur, G.N.; Bu, G. Dkk1 Stabilizes Wnt Co-Receptor LRP6: Implication for Wnt Ligand-Induced LRP6 Down-Regulation. PLoS ONE 2010, 5, e11014. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhang, J.; Han, X.; Wang, G. LncRNA SNHG1 Delayed Fracture Healing via Modulating MiR-181a-5p/PTEN Axis. J. Investig. Surg. 2022, 35, 1304–1312. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.; King, R.M.; Philipson, L. Genes Specifically Expressed at Growth Arrest of Mammalian Cells. Cell 1988, 54, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Park, S.J.; Jung, S.-H.; Kim, E.J.; Jogeswar, G.; Ajita, J.; Rhee, Y.; Kim, C.-H.; Lim, S.-K. MiR-182 Is a Negative Regulator of Osteoblast Proliferation, Differentiation, and Skeletogenesis through Targeting FoxO1. J. Bone Miner. Res. 2012, 27, 1669–1679. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, K.; Miyazono, K. Regulation of TGF-β Family Signaling by Inhibitory Smads. Cold Spring Harb. Perspect. Biol. 2017, 9, a022095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Xie, Z.; Li, J.; Lin, J.; Zheng, G.; Liu, W.; Tang, S.; Cen, S.; Ye, G.; Li, Z.; et al. GAS5 Protects against Osteoporosis by Targeting UPF1/SMAD7 Axis in Osteoblast Differentiation. eLife 2020, 9, e59079. [Google Scholar] [CrossRef]
- Cho, H.; Li, Y.; Archacki, S.; Wang, F.; Yu, G.; Chakrabarti, S.; Guo, Y.; Chen, Q.; Wang, Q.K. Splice Variants of LncRNA RNA ANRIL Exert Opposing Effects on Endothelial Cell Activities Associated with Coronary Artery Disease. RNA Biol. 2020, 17, 1391–1401. [Google Scholar] [CrossRef]
- Wu, S.; Nitschke, K.; Worst, T.S.; Fierek, A.; Weis, C.-A.; Eckstein, M.; Porubsky, S.; Kriegmair, M.; Erben, P. Long Noncoding RNA MIR31HG and Its Splice Variants Regulate Proliferation and Migration: Prognostic Implications for Muscle Invasive Bladder Cancer. J. Exp. Clin. Cancer Res. 2020, 39, 288. [Google Scholar] [CrossRef]
- Gao, J.; Zeng, K.; Liu, Y.; Gao, L.; Liu, L. LncRNA SNHG5 Promotes Growth and Invasion in Melanoma by Regulating the MiR-26a-5p/TRPC3 Pathway. Onco. Targets Ther. 2018, 12, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Huang, Y.; Yang, Q.; Jia, L.; Zheng, Y.; Li, W. Long Non-Coding RNA SNHG5 Mediates Periodontal Inflammation through the NF-ΚB Signalling Pathway. J. Clin. Periodontol. 2022, 49, 1038–1051. [Google Scholar] [CrossRef]
- Ota, T.; Suzuki, Y.; Nishikawa, T.; Otsuki, T.; Sugiyama, T.; Irie, R.; Wakamatsu, A.; Hayashi, K.; Sato, H.; Nagai, K.; et al. Complete Sequencing and Characterization of 21,243 Full-Length Human CDNAs. Nat. Genet. 2004, 36, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Chen, J.; Li, C.; Yuan, Y.; Fang, S.; Liu, W.; Qian, Y.; Ma, J.; Chang, L.; Chen, F.; et al. Exosome-Mediated Transfer of SNHG7 Enhances Docetaxel Resistance in Lung Adenocarcinoma. Cancer Lett. 2022, 526, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Janssens, K.; ten Dijke, P.; Ralston, S.H.; Bergmann, C.; Van Hul, W. Transforming Growth Factor-Β1 Mutations in Camurati-Engelmann Disease Lead to Increased Signaling by Altering Either Activation or Secretion of the Mutant Protein. J. Biol. Chem. 2003, 278, 7718–7724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Liu, Z.; Shen, L.; Jiang, H. Long non-coding RNA SNHG7 promotes the fracture repair through negative modulation of miR-9. Am. J. Transl. Res. 2019, 11, 974–982. [Google Scholar] [PubMed]
- Differential Expression of lncRNA/miRNA/mRNA and their Related Functional Networks during the Osteogenic/Odontogenic Differentiation of Dental Pulp Stem Cells. Available online: https://onlinelibrary.wiley.com/doi/epdf/10.1002/jcp.29223 (accessed on 2 August 2022).
- Yang, Z.; Feng, L.; Wang, H.; Li, Y.; Lo, J.H.T.; Zhang, X.; Lu, X.; Wang, Y.; Lin, S.; Tortorella, M.D.; et al. DANCR Mediates the Rescuing Effects of Sesamin on Postmenopausal Osteoporosis Treatment via Orchestrating Osteogenesis and Osteoclastogenesis. Nutrients 2021, 13, 4455. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, M.; Liang, L.; Li, J.; Chen, Y.-X. Over-Expression of LncRNA DANCR Is Associated with Advanced Tumor Progression and Poor Prognosis in Patients with Colorectal Cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 11480–11484. [Google Scholar]
- Zhang, L.; Yang, C.; Chen, S.; Wang, G.; Shi, B.; Tao, X.; Zhou, L.; Zhao, J. Long Noncoding RNA DANCR Is a Positive Regulator of Proliferation and Chondrogenic Differentiation in Human Synovium-Derived Stem Cells. DNA Cell Biol. 2017, 36, 136–142. [Google Scholar] [CrossRef]
- Tang, Z.; Gong, Z.; Sun, X. LncRNA DANCR Involved Osteolysis after Total Hip Arthroplasty by Regulating FOXO1 Expression to Inhibit Osteoblast Differentiation. J. Biomed. Sci. 2018, 25, 4. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Yin, Y.; Jiang, F.; Chen, N. Human Amnion Mesenchymal Stem Cells Promote Proliferation and Osteogenic Differentiation in Human Bone Marrow Mesenchymal Stem Cells. J. Mol. Hist. 2015, 46, 13–20. [Google Scholar] [CrossRef]
- Jiang, S.Y.; Miao, Y.X.; Hirokazu, T.; Zhu, S.Z.; Lu, J.S. Effects of lncRNA DANCR on proliferation and differentiation of osteoblasts by regulating the Wnt/β-catenin pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5558–5566. [Google Scholar] [CrossRef]
- Wang, C.; Yang, Q.; Han, Y.; Liu, H.; Wang, Y.; Huang, Y.; Zheng, Y.; Li, W. A Reduced Level of the Long Non-Coding RNA SNHG8 Activates the NF-KappaB Pathway by Releasing Functional HIF-1alpha in a Hypoxic Inflammatory Microenvironment. Stem. Cell Res. Ther 2022, 13, 229. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, Y.; Zhao, Z.; Han, X.; Chen, Y. Knockdown of DANCR Reduces Osteoclastogenesis and Root Resorption Induced by Compression Force via Jagged1. Cell Cycle 2019, 18, 1759–1769. [Google Scholar] [CrossRef] [PubMed]
- Xue, Q.; Yang, L.; Wang, H.; Han, S. Silence of Long Noncoding RNA SNHG14 Alleviates Ischemia/Reperfusion-Induced Acute Kidney Injury by Regulating MiR-124-3p/MMP2 Axis. Biomed. Res. Int. 2021, 2021, 8884438. [Google Scholar] [CrossRef] [PubMed]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A CeRNA Hypothesis: The Rosetta Stone of a Hidden RNA Language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paspaliaris, V.; Kolios, G. Stem Cells in Osteoporosis: From Biology to New Therapeutic Approaches. Stem Cells Int. 2019, 2019, 1730978. [Google Scholar] [CrossRef] [Green Version]
- De Witte, T.-M.; Fratila-Apachitei, L.E.; Zadpoor, A.A.; Peppas, N.A. Bone Tissue Engineering via Growth Factor Delivery: From Scaffolds to Complex Matrices. Regen. Biomater. 2018, 5, 197–211. [Google Scholar] [CrossRef]
Figure 1.
The expression of SNHG8 increased in osteogenic medium. For the induction of osteoblast differentiation, PDLSCs were cultured in osteogenic medium (OM), which was composed of growth medium supplemented with b-glycerophosphate (10 mM), dexamethasone (100 nM) and vitamin C (200 mM). The relative mRNA expression of (A) alkaline phosphatase (ALP), (B) osteocalcin (OCN) and (C) small nucleolar RNA host gene 8 (SNHG8) was measured by qRT-PCR on the second day of osteogenic induction. Three repeated experiments were conducted. Data are presented as the mean ± SD, * p < 0.05, ** p < 0.01.
Figure 1.
The expression of SNHG8 increased in osteogenic medium. For the induction of osteoblast differentiation, PDLSCs were cultured in osteogenic medium (OM), which was composed of growth medium supplemented with b-glycerophosphate (10 mM), dexamethasone (100 nM) and vitamin C (200 mM). The relative mRNA expression of (A) alkaline phosphatase (ALP), (B) osteocalcin (OCN) and (C) small nucleolar RNA host gene 8 (SNHG8) was measured by qRT-PCR on the second day of osteogenic induction. Three repeated experiments were conducted. Data are presented as the mean ± SD, * p < 0.05, ** p < 0.01.
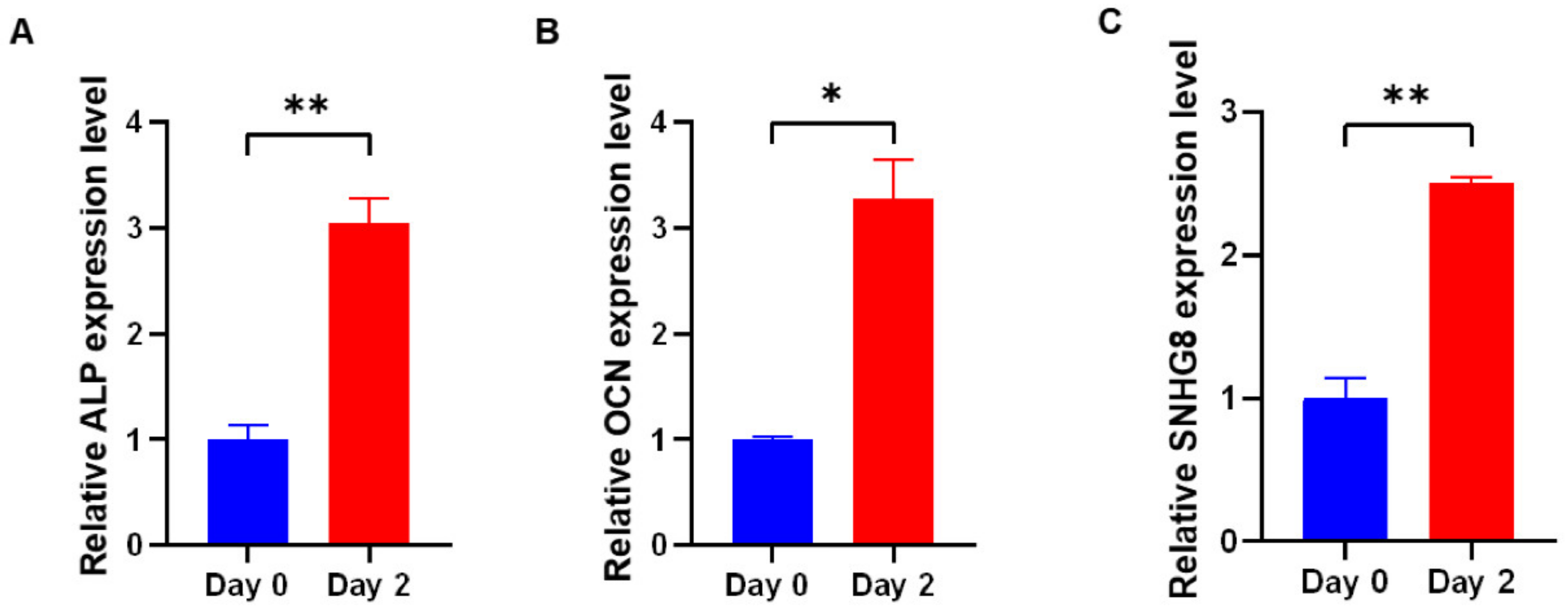
Figure 2.
Schematic illustration of lncRNA GAS5 regulating osteogenesis of BMSCs by directly binding to protein UPF1. GAS5 binds to the HD region of UPF1 specifically, which results in binding of UPF1 and SMAD7 (mRNA). After binding to UPF1, SMAD7 (mRNA) degrades. As an inhibitor of the SMAD 1/5/8 pathway, degradation of SMAD7 promotes activation of SMAD 1/5/8 pathway and ultimately promotes osteoblast differentiation of BMSCs.
Figure 2.
Schematic illustration of lncRNA GAS5 regulating osteogenesis of BMSCs by directly binding to protein UPF1. GAS5 binds to the HD region of UPF1 specifically, which results in binding of UPF1 and SMAD7 (mRNA). After binding to UPF1, SMAD7 (mRNA) degrades. As an inhibitor of the SMAD 1/5/8 pathway, degradation of SMAD7 promotes activation of SMAD 1/5/8 pathway and ultimately promotes osteoblast differentiation of BMSCs.

Figure 3.
Schematic illustration of lncRNA DANCR regulating osteoblast differentiation of MSC by directly binding to protein FOXO1. Through unknown mechanisms, DANCR inhibits FOXO1 expression. Additionally, DANCR directly binds to FOXO1 and promotes Skp2-mediated ubiquitination of FOXO1, which further reduces FOXO1 expression. FOXO1, a transcription factor, regulates proliferation, differentiation and apoptosis of osteoblasts. By decreasing FOXO1 expression, DANCR ultimately inhibits osteoblast differentiation of MSC. U.M., unknown mechanism.
Figure 3.
Schematic illustration of lncRNA DANCR regulating osteoblast differentiation of MSC by directly binding to protein FOXO1. Through unknown mechanisms, DANCR inhibits FOXO1 expression. Additionally, DANCR directly binds to FOXO1 and promotes Skp2-mediated ubiquitination of FOXO1, which further reduces FOXO1 expression. FOXO1, a transcription factor, regulates proliferation, differentiation and apoptosis of osteoblasts. By decreasing FOXO1 expression, DANCR ultimately inhibits osteoblast differentiation of MSC. U.M., unknown mechanism.


{kind=link}
{kind=link}
{kind=link}
Table 1.
SNHG expression trends during osteoblast differentiation.
| LncRNA | Cell | Expression Trend during Osteoblast Differentiation | References |
|---|---|---|---|
| SNHG1 | BMSC | Downregulation | [6,13] |
| SNHG2 (GAS5) | hDPSC/BMSC | Upregulation | [17,19] |
| GAS5 transcript variant 2 | hBMSC | Downregulation | [20] |
| SNHG5 | hBMSC | Upregulation | [11,18] |
| SNHG7 | hDPSC | Downregulation | [12] |
| SNHG13 (DANCR) | hBMSC (co-cultured with HAMSC) | Downregulation | [15] |
| SNHG13 (DANCR) | BMSC | Downregulation | [16] |
Table 2.
Key SNHGs regulate osteoblast differentiation.
| LncRNA | Cell | Regulatory Axis | LncRNA’s Effect on Osteogenesis | References |
|---|---|---|---|---|
| SNHG1 | BMSC | SNHG1/miR-181c-5p/SFRP1/Wnt3a | Inhibitory | [6] |
| SNHG1 | BMSC | SNHG1/Nedd4/p38 | Inhibitory | [24] |
| SNHG1 | BMSC | SNHG1/miR101/DKK1/Wnt/ β-catenin | Inhibitory | [13] |
| SNHG1 | MC3T3-E1 | SNHG1/miR-181a- 5p/PTEN | Inhibitory | [33] |
| SNHG2 (GAS5) | hPDLSC | SNHG2/p38/JNK | Stimulative | [17] |
| SNHG2 (GAS5) | BMSC | SNHG2/miR-135a-5p/FOXO1 | Stimulative | [19] |
| SNHG2 (GAS5) | BMSC | SNHG2/UPF1/Smad7 | Stimulative | [37] |
| GAS5 transcript variant 2 | BMSC | SNHG2/miR-382-3p/TAF1 | Inhibitory | [20] |
| SNHG5 | BMSC | SNHG5/miR-582-5p/RUNX3 | Stimulative | [11] |
| SNHG5 | BMSC | YY1/SNHG5/miR-212-3p/GDF5/Smad | Stimulative | [18] |
| SNHG7 | MC3T3-E1 | SNHG7/miR-9/TGF-β | Stimulative | [45] |
| SNHG7 | hDPSC | SNHG7/miR-6512-3p/TNF-α | Stimulative | [12] |
| SNHG13 (DANCR) | MSC | SNHG13/FOXO1 | Inhibitory | [50] |
| SNHG13 (DANCR) | BMSC | SNHG13/miR-1301-3p/PROX1 | Inhibitory | [16] |
| SNHG13 (DANCR) | BMSC | SNHG13/miR-320a/CTNNB1/Wnt/β-catenin | Inhibitory | [9] |
| SNHG13 (DANCR) | MC3T3-E1 | SNHG13/Wnt/β-catenin | Inhibitory | [52] |
| SNHG14 | hMSC | SNHG14/miR-2861/AKT2 | Stimulative | [7] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tan, A.-Q.; Zheng, Y.-F. The Roles of SNHG Family in Osteoblast Differentiation. Genes 2022, 13, 2268. https://doi.org/10.3390/genes13122268
AMA Style
Tan A-Q, Zheng Y-F. The Roles of SNHG Family in Osteoblast Differentiation. Genes. 2022; 13(12):2268. https://doi.org/10.3390/genes13122268
Chicago/Turabian StyleTan, An-Qi, and Yun-Fei Zheng. 2022. "The Roles of SNHG Family in Osteoblast Differentiation" Genes 13, no. 12: 2268. https://doi.org/10.3390/genes13122268
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.