
Characterization of Mediterranean Durum Wheat for Resistance to Pyrenophora tritici-repentis

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experimental Design and Inoculation
2.3. Climatic Conditions
2.4. Confirmation of the Pathogen
2.5. Tan Spot Evaluation
2.6. Statistical Analysis
3. Results
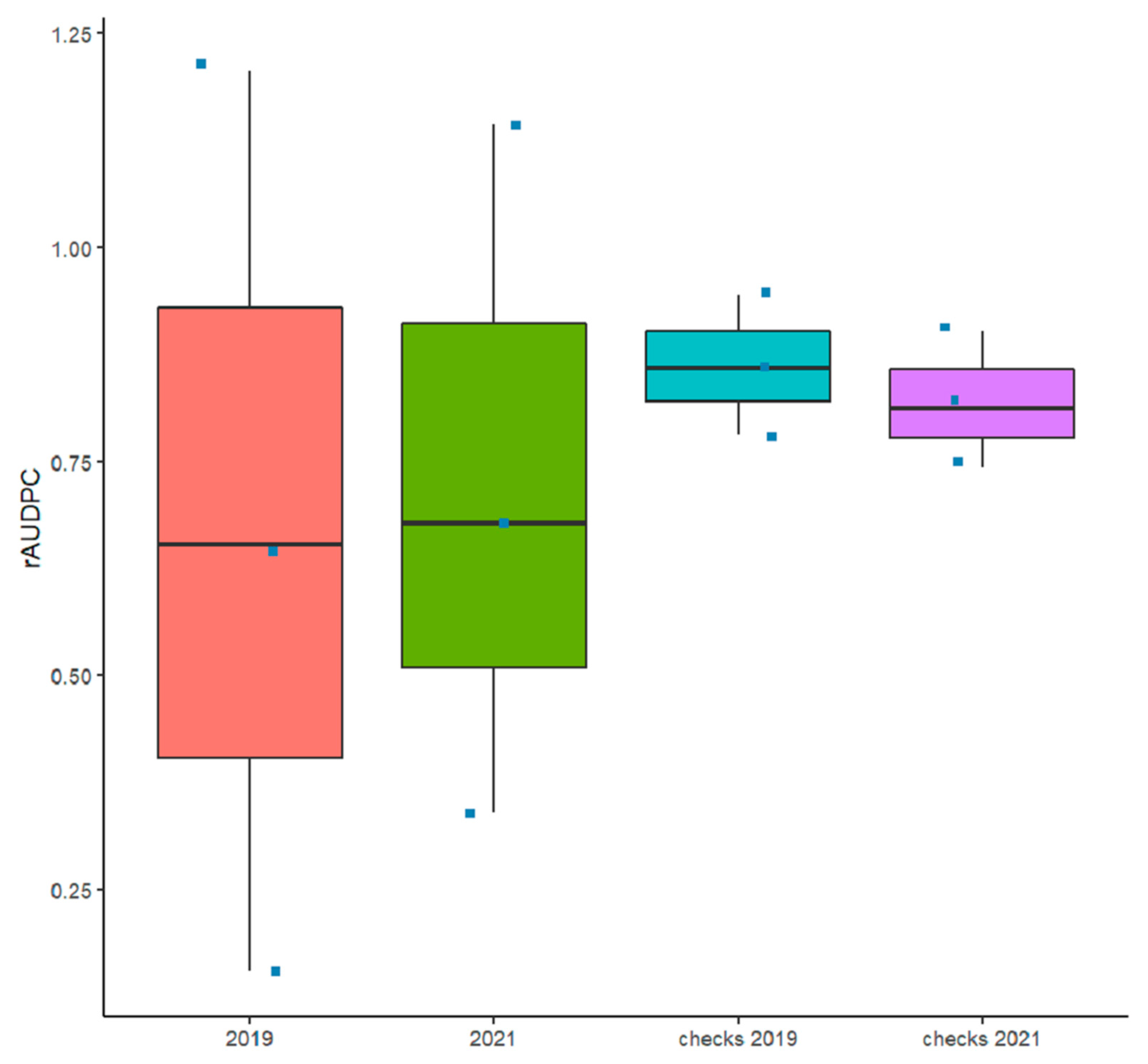
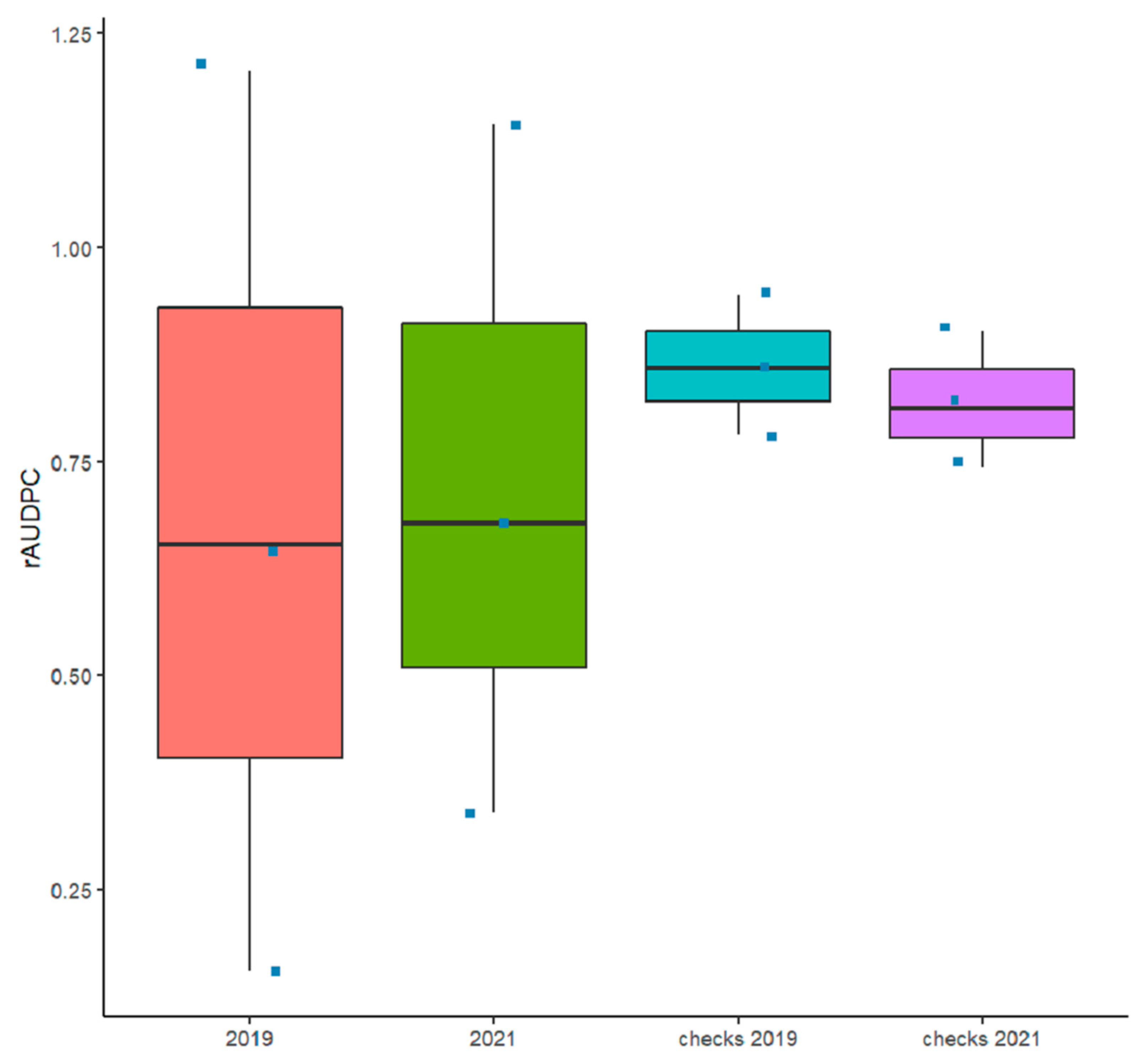
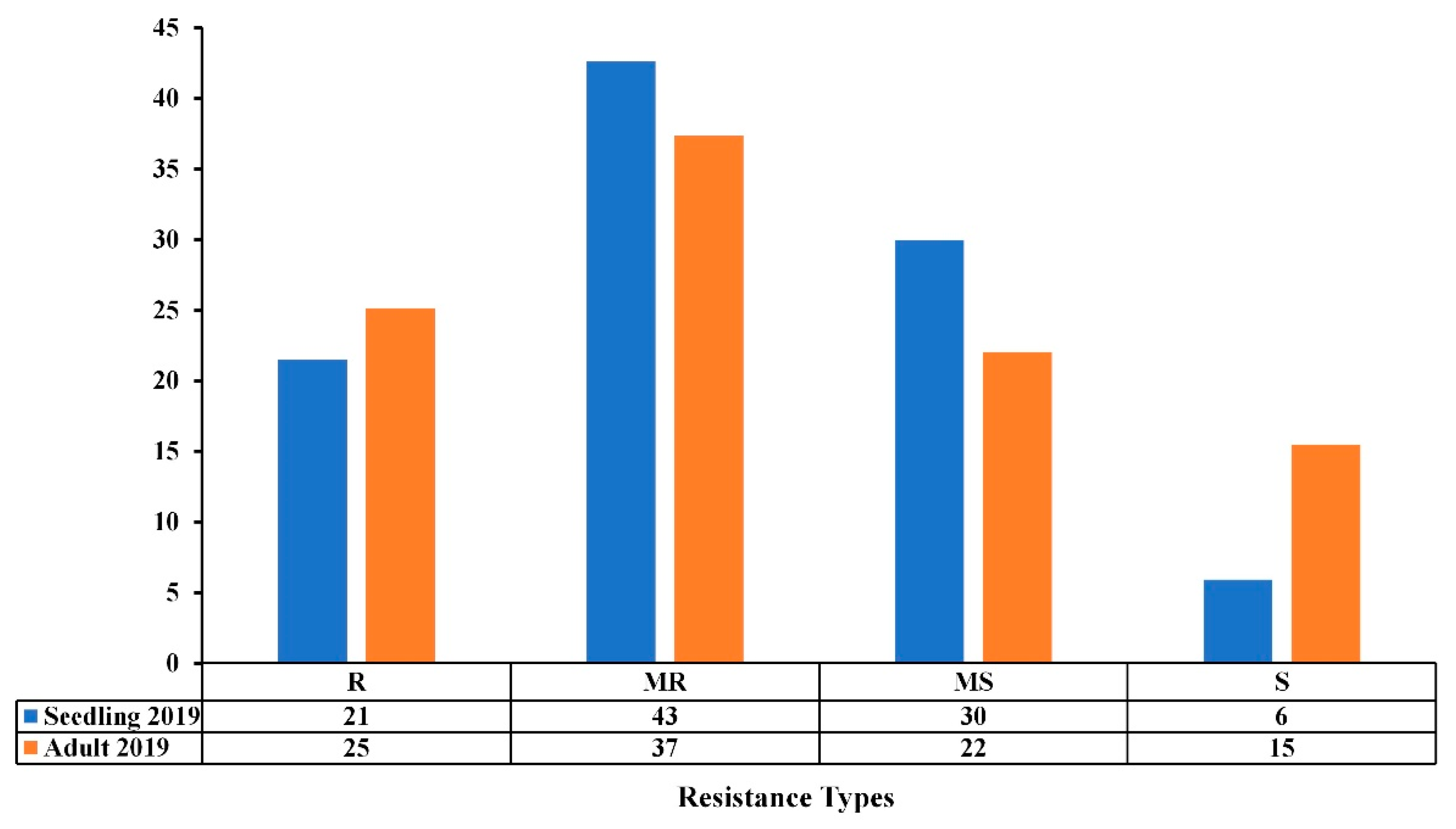
3.1. Reactions of Genotypes across the Two Trials
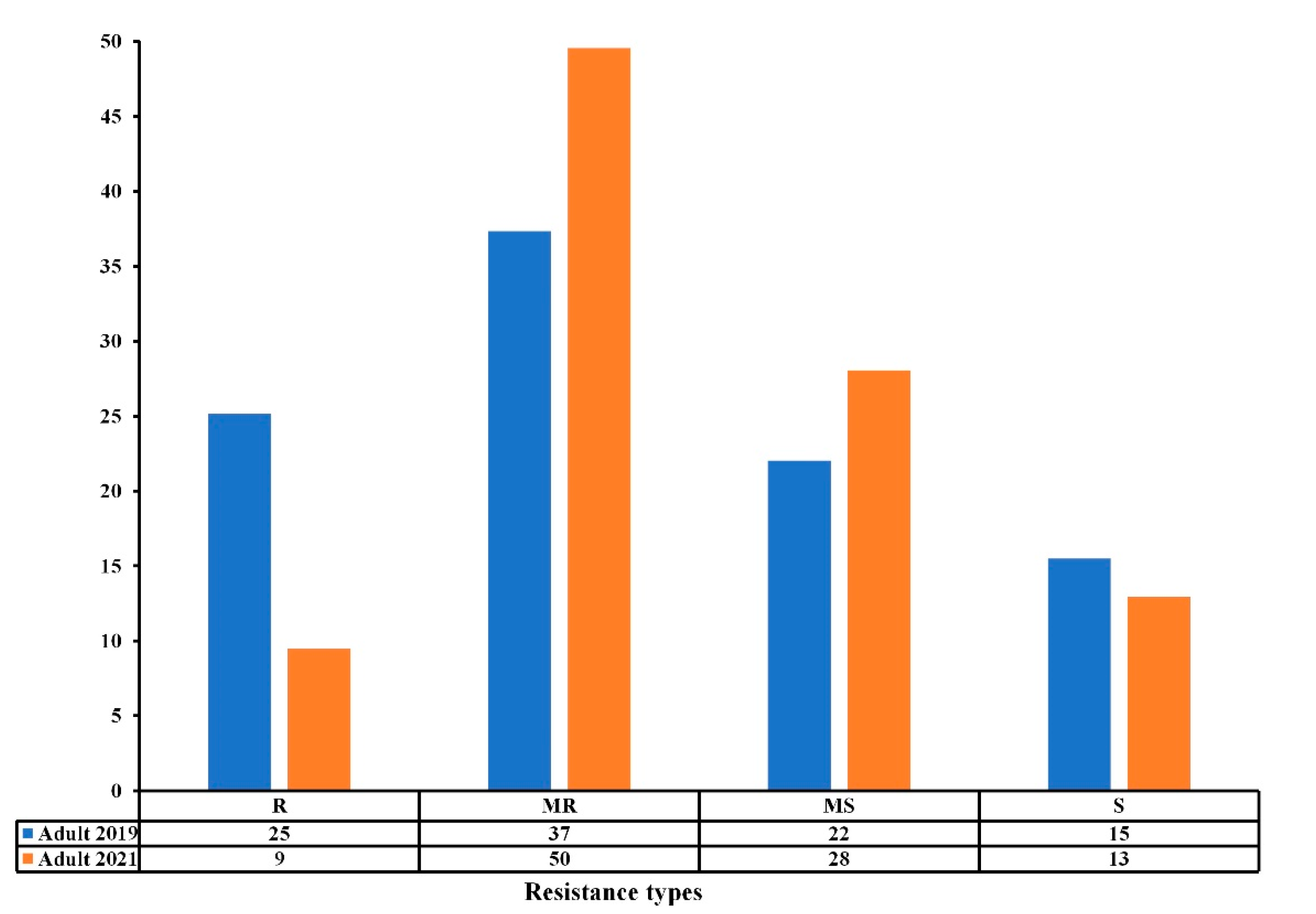
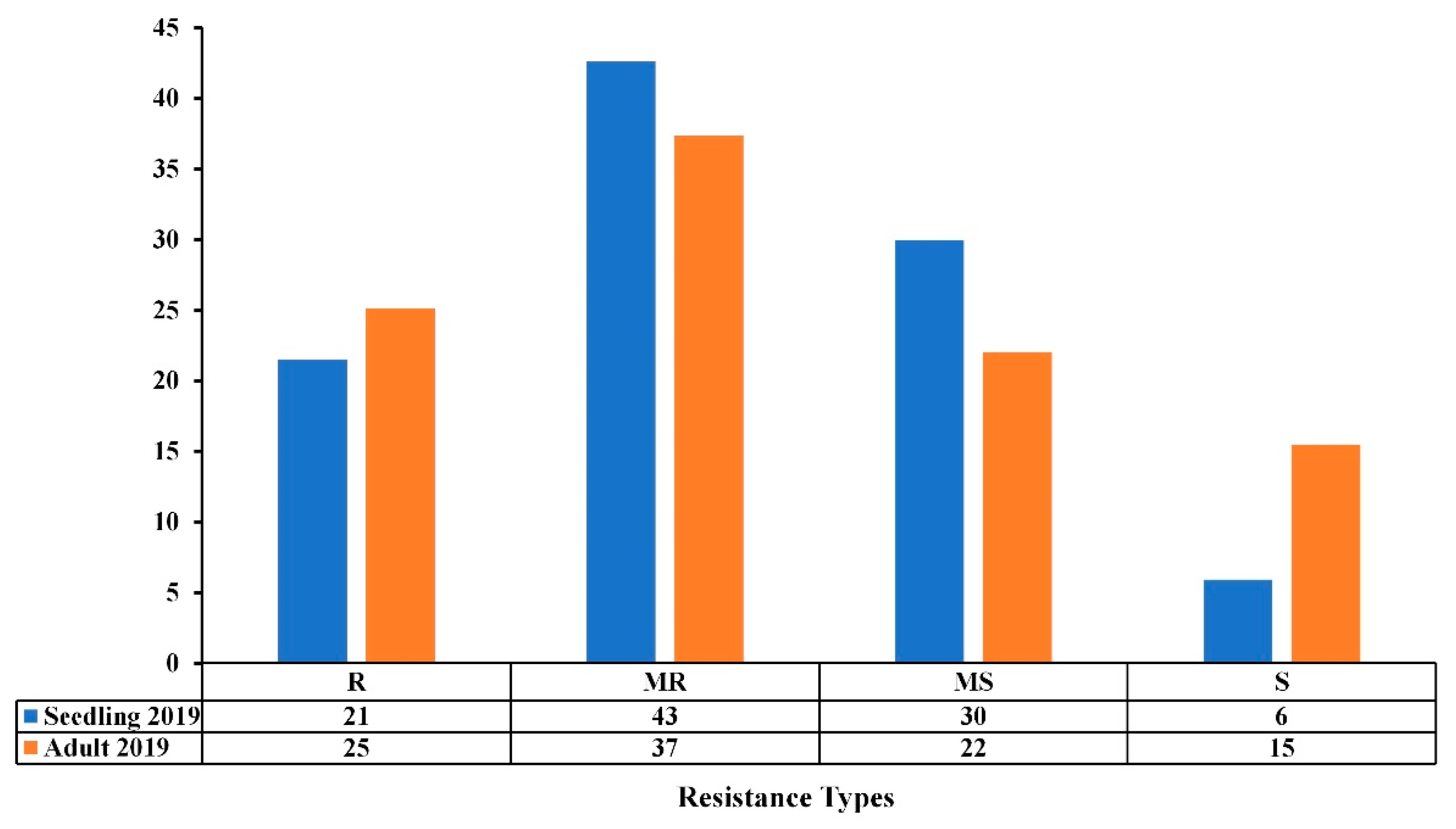
3.2. Comparison of Seedling and Adult Plant Resistance
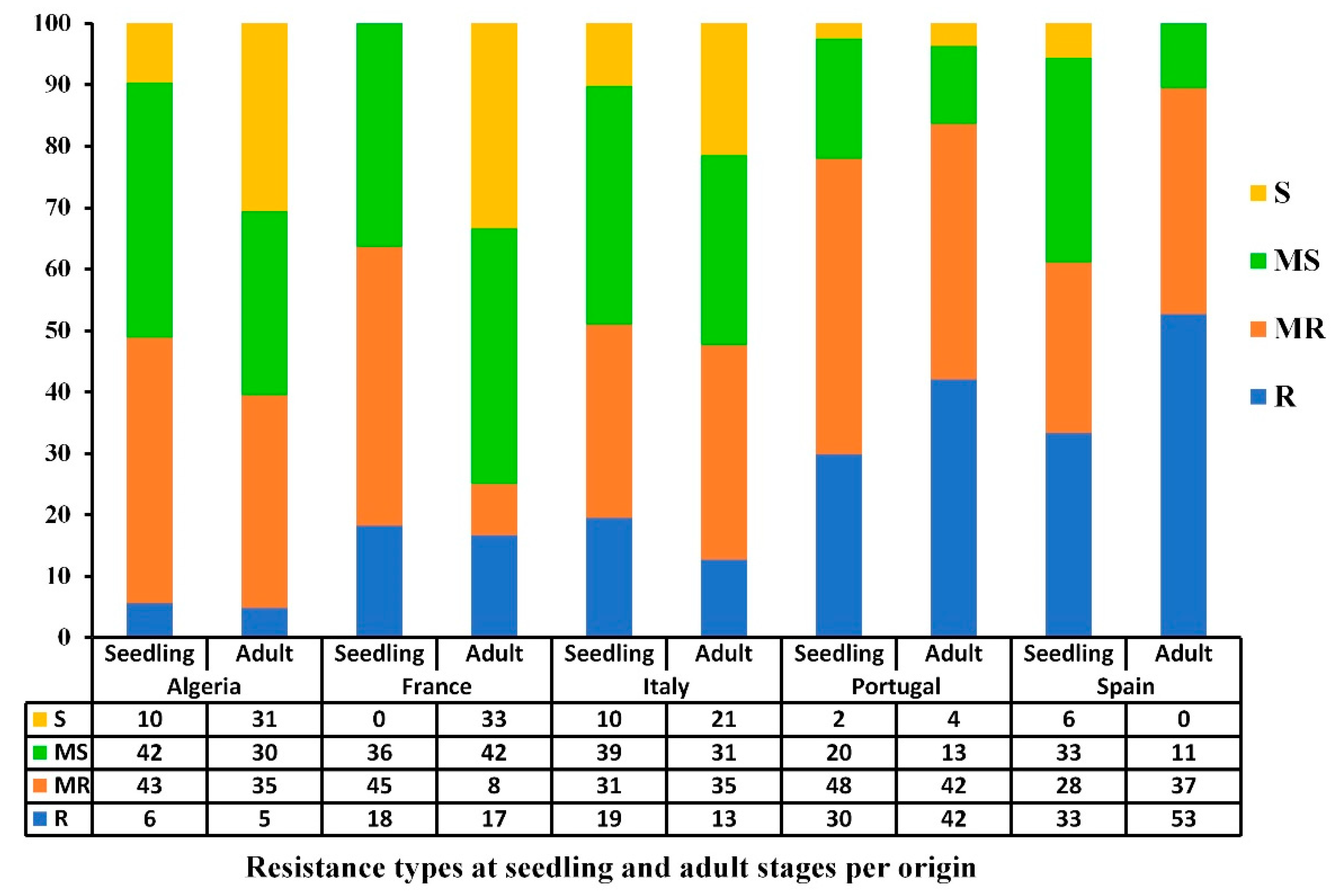
3.3. Geographical Distribution of the Resistant and Susceptible Accessions
3.4. Resistance Levels to Tan Spot with Regard to the Level of Improvement
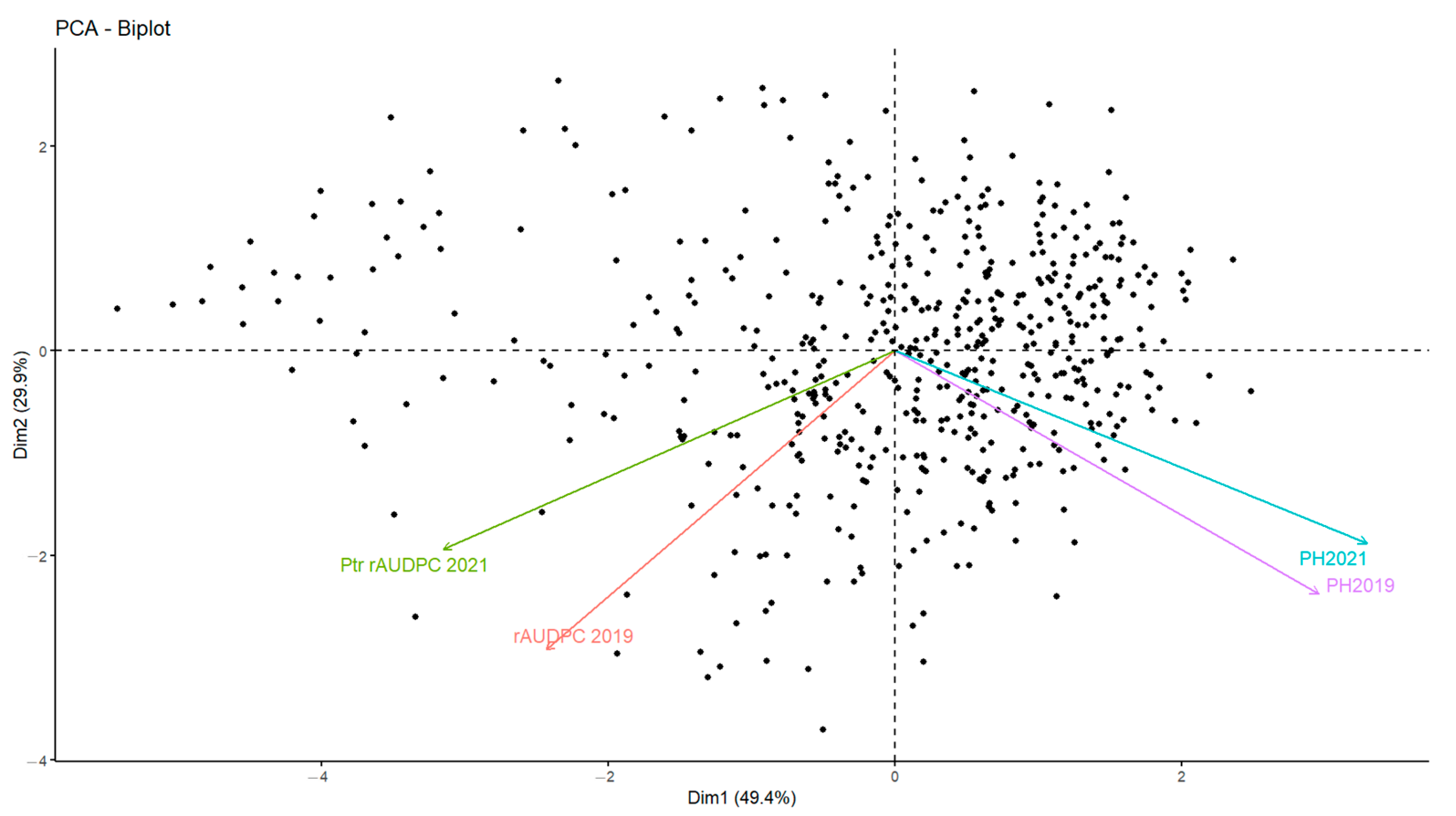
3.5. Association between Disease and Plant Height
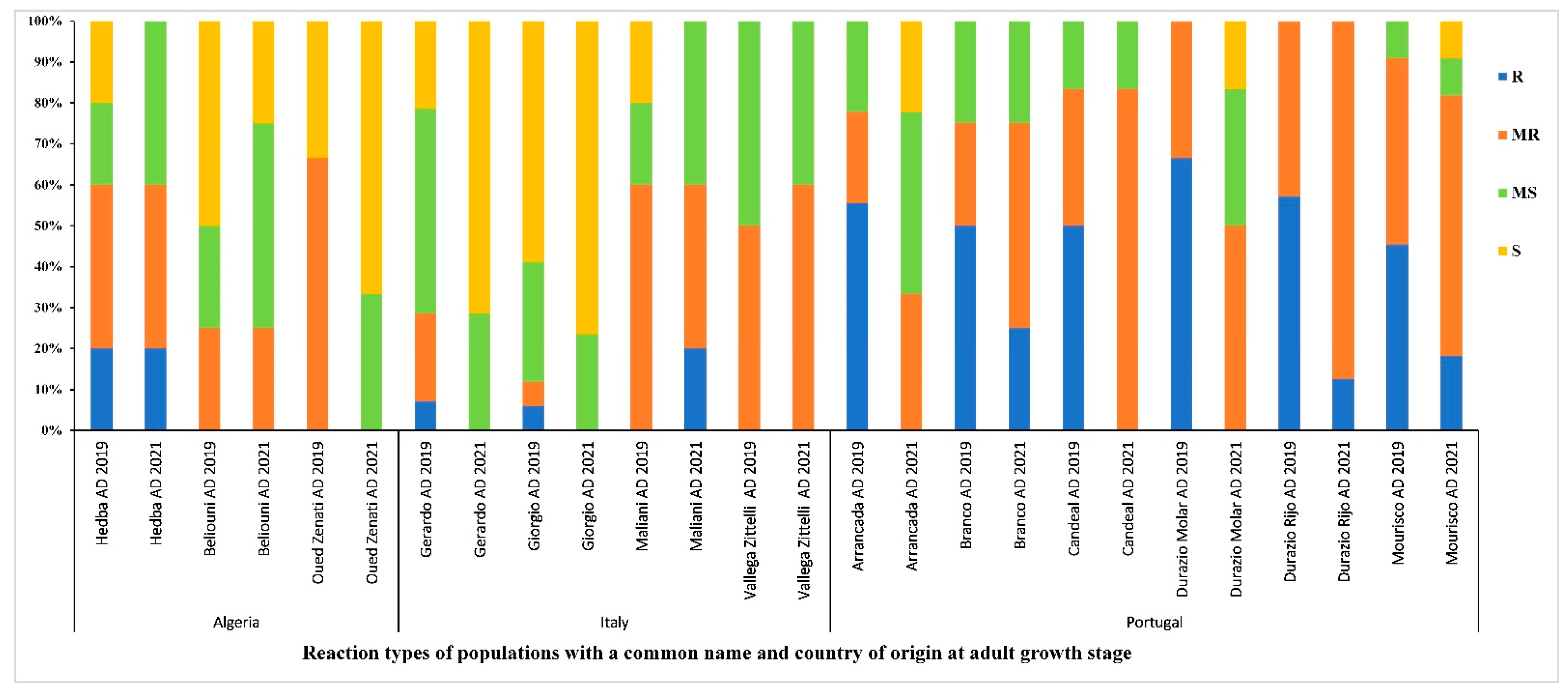
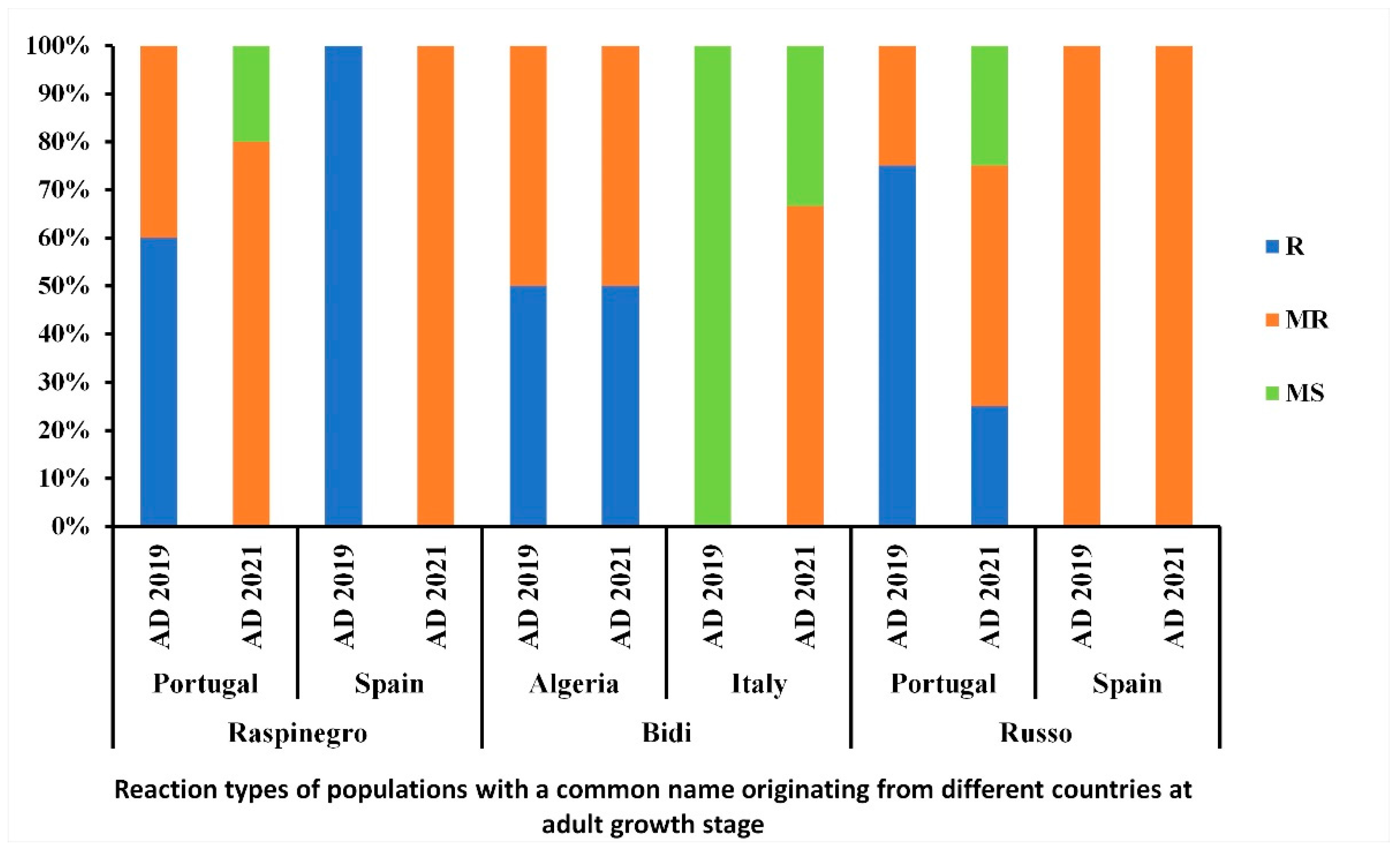
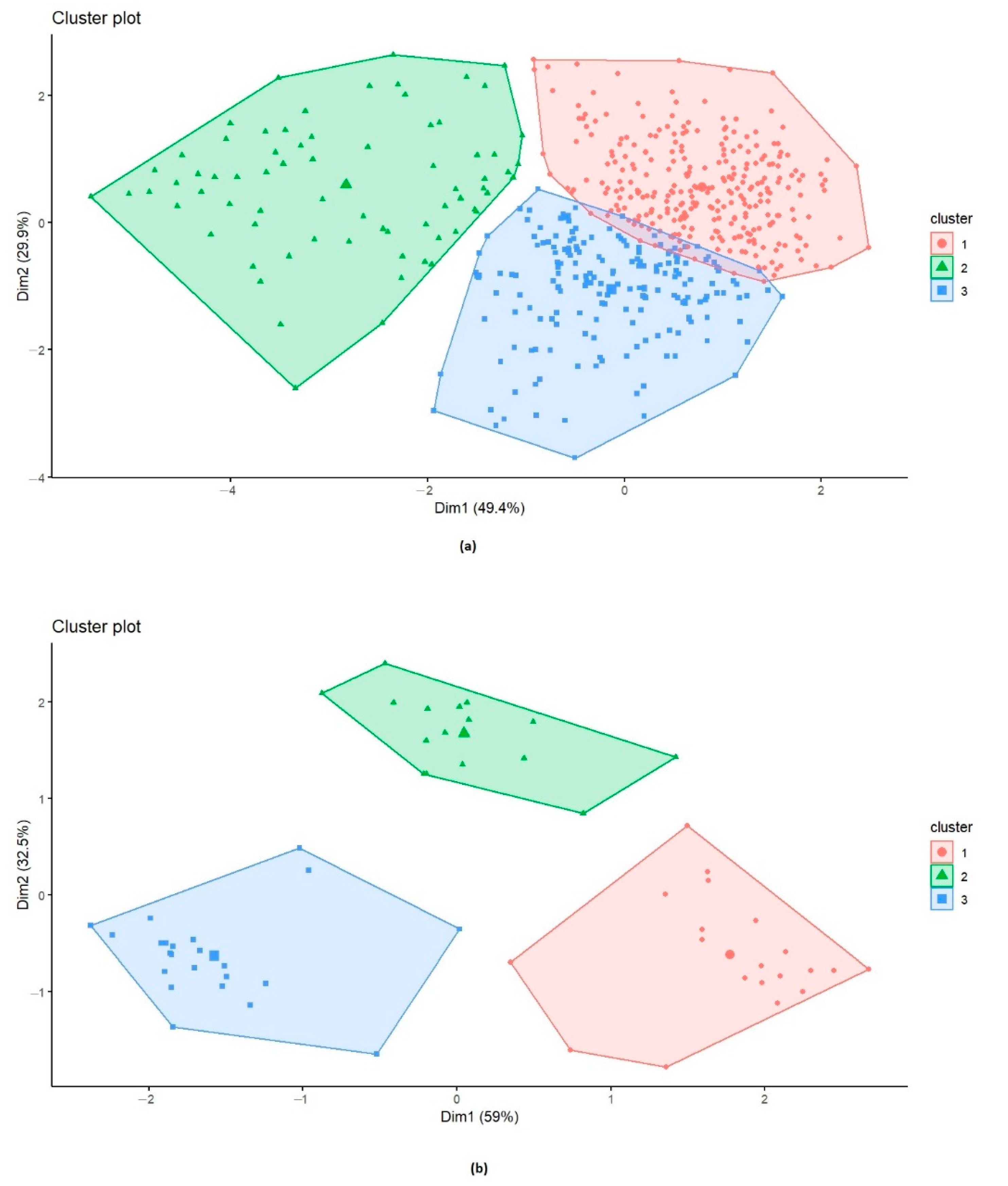
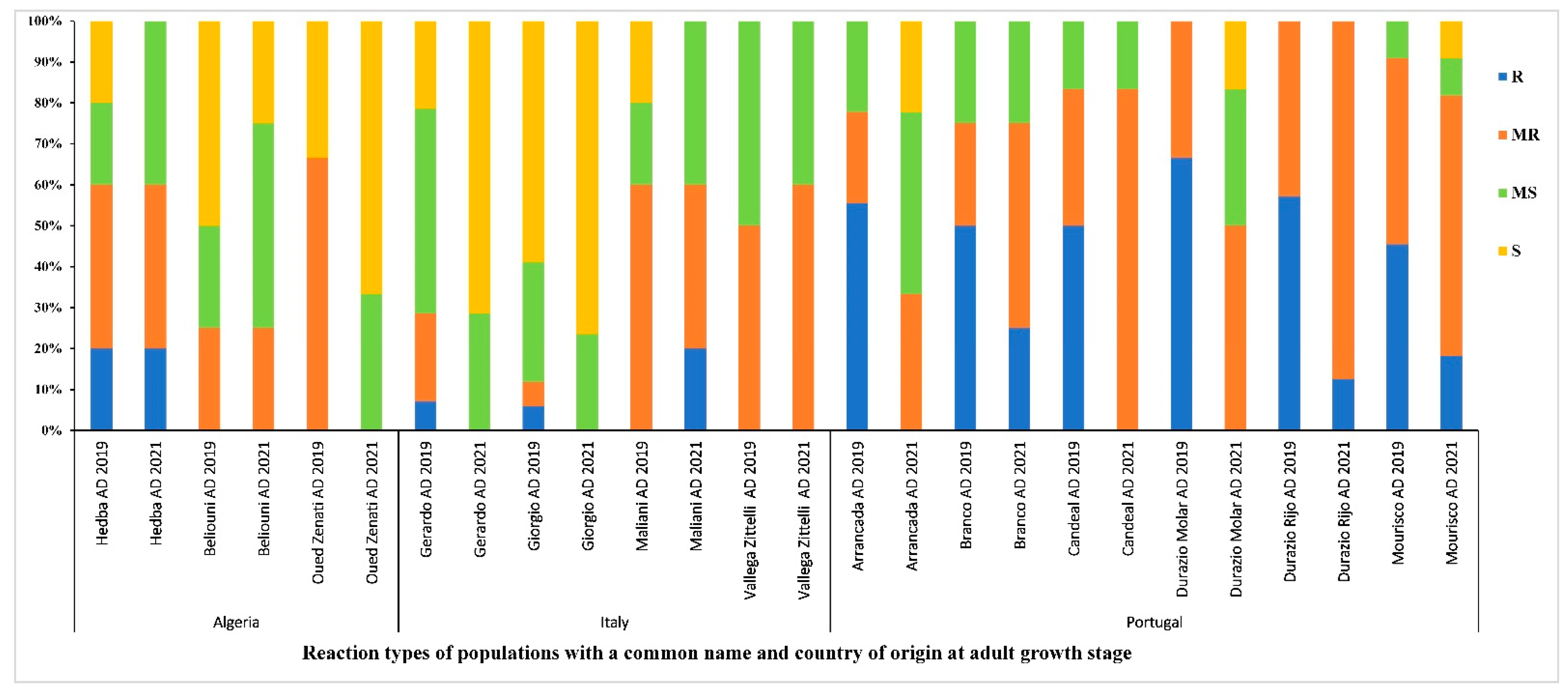
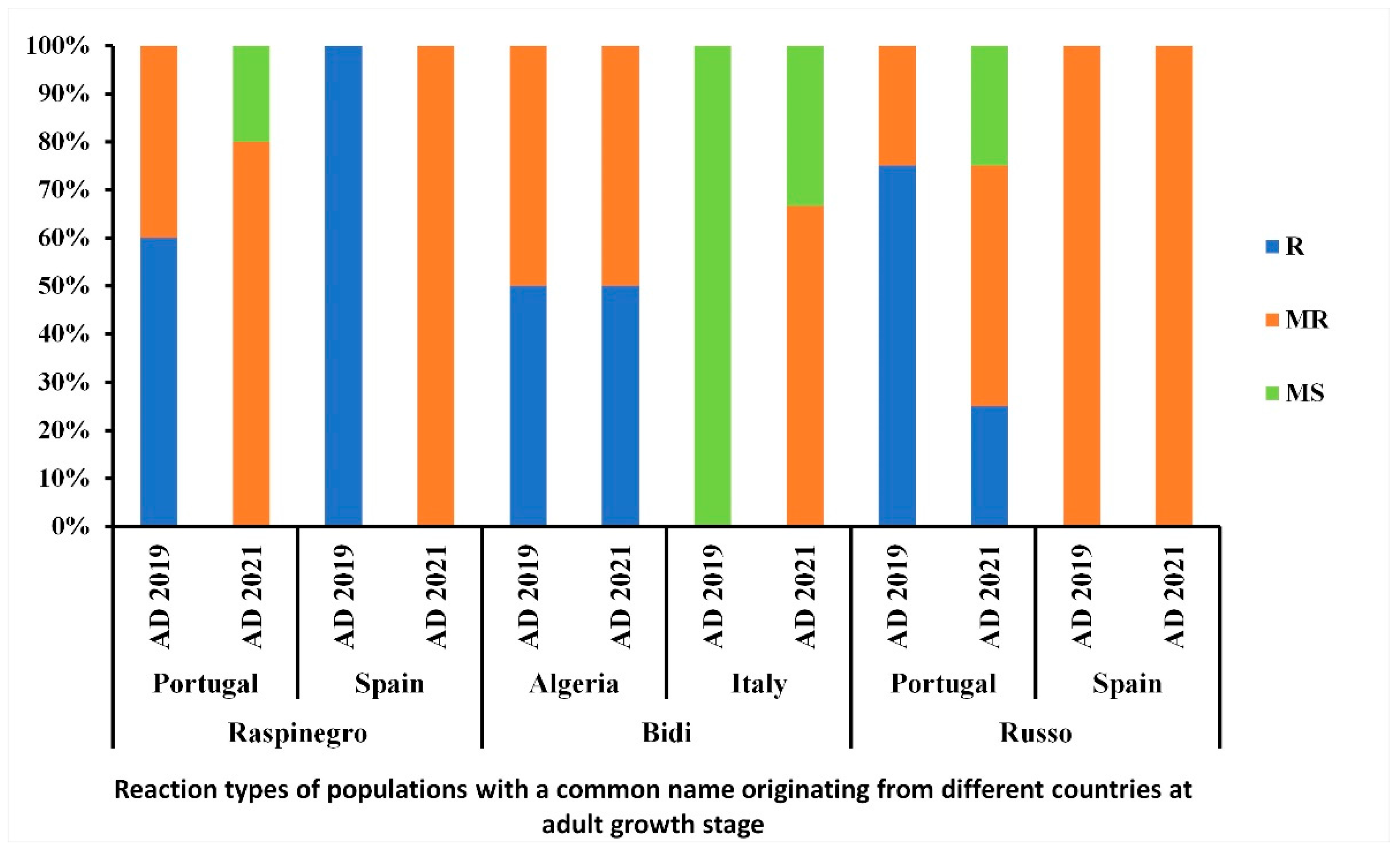
3.6. Distribution of the Reaction Types among and between Populations
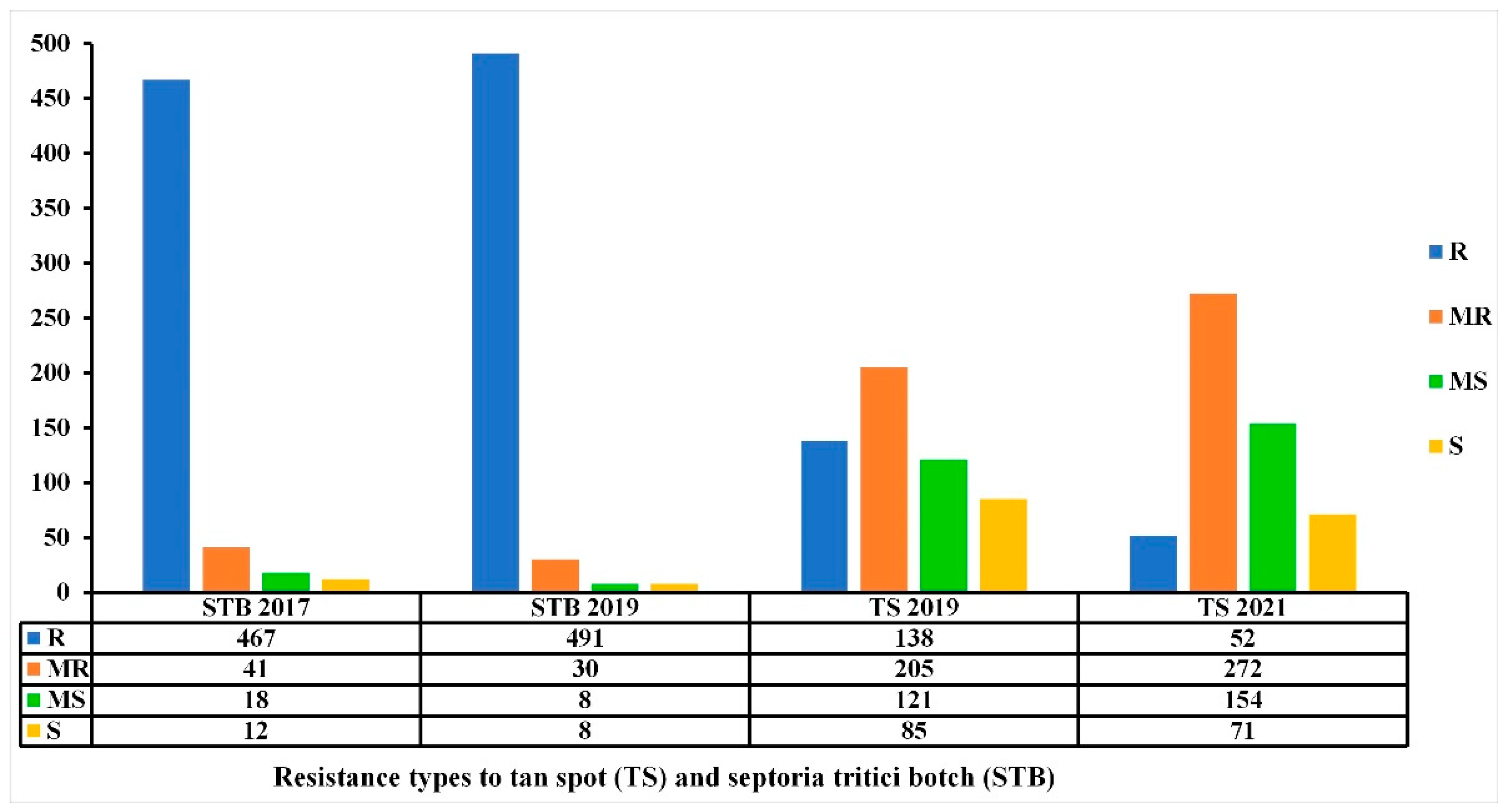
3.7. Multiple Disease Resistance: TS and STB Association with Regard to Plant Resistance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feldman, M. Origin of Cultivated Wheat. In The World Wheat Book: A History of Wheat Breeding; Bonjean, A.P., Angus, W.J., Eds.; Intercept Ltd.: London, UK, 2001; pp. 3–56. [Google Scholar]
- Nazco, R.; Villegas, D.; Ammar, K.; Peña, R.J.; Moragues, M.; Royo, C. Can Mediterranean Durum Wheat Landraces Contribute to Improved Grain Quality Attributes in Modern Cultivars? Euphytica 2012, 185, 1–17. [Google Scholar] [CrossRef]
- Mercer, K.L.; Perales, H.R. Evolutionary Response of Landraces to Climate Change in Centers of Crop Diversity: Climate Change and Crop Diversity. Evol. Appl. 2010, 3, 480–493. [Google Scholar] [CrossRef] [PubMed]
- López-Bellido, L.; López-Bellido, R.J.; Redondo, R. Nitrogen Efficiency in Wheat under Rainfed Mediterranean Conditions as Affected by Split Nitrogen Application. Field Crops Res. 2005, 94, 86–97. [Google Scholar] [CrossRef]
- Soriano, J.M.; Villegas, D.; Sorrells, M.E.; Royo, C. Durum Wheat Landraces from East and West Regions of the Mediterranean Basin Are Genetically Distinct for Yield Components and Phenology. Front. Plant Sci. 2018, 9, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, M.S.; El-Basyoni, I.; Baenziger, P.S.; Singh, S.; Royo, C.; Ozbek, K.; Aktas, H.; Ozer, E.; Ozdemir, F.; Manickavelu, A.; et al. Exploiting Genetic Diversity from Landraces in Wheat Breeding for Adaptation to Climate Change. J. Exp. Bot. 2015, 66, 3477–3486. [Google Scholar] [CrossRef] [PubMed]
- USDA. Tunisia: Grain and Feed Annual; USDA Foreign Agricultural Service: Washington, DC, USA, 2019.
- Bahri, B.A.; Leconte, M.; Rebai, H.; Pope De Vallavieille, C. Wheat Yellow Rust Dynamics in Tunisia since 2013 and Resistance Genes in Durum Wheat. Phytopathology 2016, 106, 84. [Google Scholar]
- Chedli, R.B.H.; M’Barek, S.B.; Yahyaoui, A.; Kehel, Z.; Rezgui, S. Occurrence of Septoria Tritici Blotch (Zymoseptoria tritici) Disease on Durum Wheat, Triticale, and Bread Wheat in Northern Tunisia. Chil. J. Agric. Res. 2018, 78, 559–568. [Google Scholar] [CrossRef]
- Ben M’Barek, S.; Karisto, P.; Abdedayem, W.; Laribi, M.; Fakhfakh, M.; Kouki, H.; Mikaberidze, A.; Yahyaoui, A. Improved Control of Septoria Tritici Blotch in Durum Wheat Using Cultivar Mixtures. Plant Pathol. 2020, 69, 1655–1665. [Google Scholar] [CrossRef]
- Kamel, S.; Cherif, M. Tan Spot of Wheat in Northern Tunisia: Distribution, Prevalence, Incidence and Severity. Cereal Res. Commun. 2021, 49, 421–432. [Google Scholar] [CrossRef]
- Rharrabti, Y.; Villegas, D.; Royo, C.; Martos-Núñez, V.; García del Moral, L.F. Durum Wheat Quality in Mediterranean Environments. Field Crops Res. 2003, 80, 133–140. [Google Scholar] [CrossRef]
- Grassini, P.; Eskridge, K.M.; Cassman, K.G. Distinguishing between Yield Advances and Yield Plateaus in Historical Crop Production Trends. Nat. Commun. 2013, 4, 2918. [Google Scholar] [CrossRef] [PubMed]
- Laribi, M.; Ben M’Barek, S.; Fakhfakh, M.; Yahyaoui, A.H.; Sassi, K. Durum Wheat Mediterranean Landraces: A Valuable Source for Resistance to Tan Spot Disease. Agriculture 2021, 11, 1148. [Google Scholar] [CrossRef]
- Krupinsky, J.M. Observations on the Host Range of Isolates of Pyrenophora trichostoma. Can. J. Plant Pathol. 1982, 4, 42–46. [Google Scholar] [CrossRef]
- De Wolf, E.D.; Effertz, R.J.; Ali, S.; Francl, L.J. Vistas of Tan Spot Research. Can. J. Plant Pathol. Rev. Can. Phytopathol. 1998, 20, 349–370. [Google Scholar] [CrossRef]
- Ali, S.; Francl, L.J. Population Race Structure of Pyrenophora tritici-repentis Prevalent on Wheat and Noncereal Grasses in the Great Plains. Plant Dis. 2003, 87, 418–422. [Google Scholar] [CrossRef] [Green Version]
- Wei, B.; Moscou, M.J.; Sato, K.; Gourlie, R.; Strelkov, S.; Aboukhaddour, R. Identification of a Locus Conferring Dominant Susceptibility to Pyrenophora tritici-repentis in Barley. Front. Plant Sci. 2020, 11, 158. [Google Scholar] [CrossRef] [Green Version]
- Kamel, S.; Cherif, M.; Hafez, M.; Despins, T.; Aboukhaddour, R. Pyrenophora Tritici–Repentis in Tunisia: Race Structure and Effector Genes. Front. Plant Sci. 2019, 10, 1562. [Google Scholar] [CrossRef] [Green Version]
- Lamari, L.; Strelkov, S.E. Minireview/ Minisynthèse The Wheat/ Pyrenophora tritici-repentis Interaction: Progress towards an Understanding of Tan Spot Disease. Can. J. Plant Pathol. 2010, 32, 4–10. [Google Scholar] [CrossRef]
- Benslimane, H.; LamarI, L.; Benbelkacem, A.; Sayoud, R.; Bouznad, Z. Distribution of Races of Pyrenophora Tritici-Repentis in Algeria and Identication of a New Virulence Type. Phytopathol. Mediterr. 2011, 50, 203–211. [Google Scholar]
- Benslimane, H.; Bouznad, Z.; Aouali, S.; Khalfi, A.; Benbelkacem, A.; Sayoud, R. Prévalence de La Tache Bronzée Du Blé Causée Pyrenophora tritici-repentis En Algérie. In Proceedings of the 6eme Journees Scientifiques et Techniques Phytosanitaires, Alger, Algeria, 20–21 June 2006. [Google Scholar]
- Gamba, F.M.; Bassi, F.M.; Finckh, M.R. Race Structure of Pyrenophora tritici-repentis in Morocco. Phytopathol. Mediterr. 2017, 56, 119–126. [Google Scholar]
- Laribi, M.; Gamba, F.M.; Hassine, M.; Singh, P.K.; Yahyaoui, A.; Sassi, K. Race Structure and Distribution of Pyrenophora Tritici-Repentis in Tunisia. Phytopathol. Mediterr. 2019, 58, 473–483. [Google Scholar] [CrossRef]
- Laribi, M.; Akhavan, A.; M’Barek, S.; Yahyaoui, A.; Strelkov, S.E.; Sassi, K. Characterization of Pyrenophora Tritici-Repentis in Tunisia and Comparison with a Global Pathogen Population. Plant Dis. 2021. [Google Scholar] [CrossRef] [PubMed]
- Ballance, G.M.; Lamari, L.; Bernier, C.C. Purification and Characterization of a Host-Selective Necrosis Toxin from Pyrenophora tritici-repentis. Physiol. Mol. Plant Pathol. 1989, 35, 203–213. [Google Scholar] [CrossRef]
- Effertz, R.J.; Meinhardt, S.W.; Anderson, J.A.; Jordahl, J.G.; Francl, L.J. Identification of a Chlorosis-Inducing Toxin from Pyrenophora tritici-repentis and the Chromosomal Location of an Insensitivity Locus in Wheat. Phytopathology 2002, 92, 527–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orolaza, N.P.; Lamari, L.; Ballance, G.M. Evidence of a Host-Specific Chlorosis Toxin from Pyrenophora Tritici-Repentis, the Causal Agent of Tan Spot of Wheat. Phytopathology 1995, 85, 1282–1287. [Google Scholar] [CrossRef]
- Ciuffetti, L.M.; Manning, V.A.; Martinez, J.P.; Pandelova, I.; Andrie, R.M. Proteinaceous Toxins of Pyrenophora tritici-repentis and Investigation of the Site-of-Action of Ptr ToxA. In Proceedings of the Fourth International Wheat Tan Spot and Spot Blotch Workshop, Bemidji, MN, USA, 21–24 July 2002; pp. 96–102. [Google Scholar]
- Meinhardt, S.; Ali, S.; Ling, H.; Francl, L. A New Race of Pyrenophora tritici-repentis That Produces a Putative Host-Selective Toxin. In Proceedings of the Fourth International Wheat Tan Spot and Spot Blotch Workshop, Bemidji, MN, USA, 21–24 July 2002; pp. 117–121. [Google Scholar]
- Pandelova, I.; Ciuffetti, L.M. A Proteomics-Based Approach for Identification of the ToxD Gene. Fungal Genet Newsl 2005, 52. [Google Scholar]
- Ali, S.; Gurung, S.; Adhikari, T.B. Identification and Characterization of Novel Isolates of Pyrenophora tritici-repentis from Arkansas. Plant Dis. 2010, 94, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Andrie, R.M.; Pandelova, I.; Ciuffetti, L.M. A Combination of Phenotypic and Genotypic Characterization Strengthens Pyrenophora tritici-repentis Race Identification. Phytopathology 2007, 97, 694–701. [Google Scholar] [CrossRef] [Green Version]
- Faris, J.D.; Liu, Z.; Xu, S.S. Genetics of Tan Spot Resistance in Wheat. Theor. Appl. Genet. 2013, 126, 2197–2217. [Google Scholar] [CrossRef]
- Singh, P.K.; Singh, R.P.; Duveiller, E.; Mergoum, M.; Adhikari, T.B.; Elias, E.M. Genetics of Wheat–Pyrenophora tritici-repentis Interactions. Euphytica 2010, 171, 1–13. [Google Scholar] [CrossRef]
- Aboukhaddour, R.; Hafez, M.; Strelkov, S.E.; Fernandez, M.R. Tan Spot Disease under the Lenses of Plant Pathologists. In Burleigh Dodds Series in Agricultural Science; Oliver, R., Ed.; Burleigh Dodds Science Publishing: Cambridge, UK, 2021; pp. 589–622. ISBN 978-1-78676-601-4. [Google Scholar]
- Agrios, G.N. Plant Pathology, 5th ed.; Elsevier Academic Press: Burlingt, MA, USA, 2005; pp. 79–103. [Google Scholar]
- Nsarellah, N.; Nachit, M.; Lhaloui, S. Breeding Durum Wheat for Biotic Stresses in the Mediterranean Region. In Durum Wheat Improvement in the Mediterranean Region: New Challenges; Options Méditerranéennes: Série A. Séminaires Méditerranéens n. 40; CIHEAM: Zaragoza, Spain, 2000; pp. 341–347. [Google Scholar]
- Tissaoui, S.; Mougou-Hamdane, A.; Omri-Benyoussef, N.; Nasraoui, B. Variability of Pyrenophora tritici-repentis Isolated from Different Wheat Areas of Tunisia: Morpho-Cultural Characterization, Pathogenic Analysis and Virulence Effector Genes. Arch. Phytopathol. Plant Prot. 2022, 55, 44–62. [Google Scholar] [CrossRef]
- Mundt, C.C. Use of Multiline Cultivars and Cultivar Mixtures for Disease Management. Annu. Rev. Phytopathol. 2002, 40, 381–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strelkov, S.E.; Lamari, L. Host–Parasite Interactions in Tan Spot [Pyrenophora tritici-repentis ] of Wheat. Can. J. Plant Pathol. 2003, 25, 339–349. [Google Scholar] [CrossRef]
- Ben M’Barek, S.; Laribi, M.; Kouki, H.; Castillo, D.; Araar, C.; Nefzaoui, M.; Ammar, K.; Saint Pierre, C.; Yahyaoui, A.H. Phenotyping Mediterranean durum wheat landraces for resistance to Zymoseptoria tritici in Tunisia. Genes 2022. [Google Scholar]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A Decimal Code for the Growth Stages of Cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Données Climatiques Station El Kodia Bousalim—AgriDATA. Available online: http://www.agridata.tn/dataset/donnees-climatiques-station-el-kodia-boussalem (accessed on 27 March 2020).
- Saari, E.E.; Prescott, J.M. Scale for Appraising the Foliar Intensity of Wheat Diseases. Plant Dis. Report. 1975, 59, 377–380. [Google Scholar]
- Simko, I.; Piepho, H.-P. The Area Under the Disease Progress Stairs: Calculation, Advantage, and Application. Phytopathology 2012, 102, 381–389. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 27 March 2020).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics; Fourth, S., Ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95457-0. [Google Scholar]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses; R Package Version 1.0.7; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://CRAN.R-project.org/package=factoextra (accessed on 1 April 2020).
- Maechler, M.; Rousseeuw, P.; Struyf, A.; Hubert, M.; Hornik, K. Cluster: Cluster Analysis Basics and Extensions; R Package Version 2.1.1; Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Elias, E.; Cantrell, R.G.; Hosford, R.M. Heritability of Resistance to Tan Spot in Durum Wheat and Its Association with Other Agronomic Traits. Crop Sci. 1989, 29, 299. [Google Scholar] [CrossRef]
- Evans, C.K.; Hunger, R.M.; Siegerist, W.C. Comparison of Greenhouse and Field Testing to Identify Wheat Resistant to Tan Spot. Plant Dis. 1999, 83, 269–273. [Google Scholar] [CrossRef]
- Perello, A.; Moreno, V.; Simón, M.R.; Sisterna, M. Tan Spot of Wheat (Triticum aestivum L.) Infection at Different Stages of Crop Development and Inoculum Type. Crop Prot. 2003, 22, 157–169. [Google Scholar] [CrossRef]
- Tadesse, W.; Reents, H.J.; Hsam, S.L.K.; Zeller, F.J. Relationship of Seedling and Adult Plant Resistance and Evaluation of Wheat Germplasm against Tan Spot (Pyrenophora tritici-repentis). Genet. Resour. Crop Evol. 2011, 58, 339–346. [Google Scholar] [CrossRef]
- Dinglasan, E.G.; Godwin, I.D.; Phan, H.T.T.; Tan, K.-C.; Platz, G.J.; Hickey, L.T. Vavilov Wheat Accessions Provide Useful Sources of Resistance to Tan Spot (Syn. Yellow Spot) of Wheat. Plant Pathol. 2018, 67, 1076–1087. [Google Scholar] [CrossRef]
- Rees, R.G.; Platz, G.J. Effectiveness of Incomplete Resistance to Pyrenophora tritici-repentis in Wheat. Aust. J. Agric. Res. 1989, 40, 43–48. [Google Scholar] [CrossRef]
- Dinglasan, E.G.; Peressini, T.; Marathamuthu, K.A.; See, P.T.; Snyman, L.; Platz, G.; Godwin, I.; Voss-Fels, K.P.; Moffat, C.S.; Hickey, L.T. Genetic Characterization of Adult-Plant Resistance to Tan Spot (Syn, Yellow Spot) in Wheat. Theor. Appl. Genet. 2021, 134, 2823–2839. [Google Scholar] [CrossRef]
- Dinglasan, E.; Godwin, I.D.; Mortlock, M.Y.; Hickey, L.T. Resistance to Yellow Spot in Wheat Grown under Accelerated Growth Conditions. Euphytica 2016, 209, 693–707. [Google Scholar] [CrossRef]
- Gamba, F.M.; Lamari, L. Mendelian Inheritance of Resistance to Tan Spot [Pyrenophora tritici-repentis ] in Selected Genotypes of Durum Wheat (Triticum turgidum). Can. J. Plant Pathol. 1998, 20, 408–414. [Google Scholar] [CrossRef]
- Gamba, F.M.; Lamari, L.; Brülé-Babel, A.L. Inheritance of Race-Specific Necrotic and Chlorotic Reactions Induced by Pyrenophora tritici-repentis in Hexaploid Wheats. Can. J. Plant Pathol. 1998, 20, 401–407. [Google Scholar] [CrossRef]
- Lamari, L.; Bernier, C.C.; Smith, R.B. Wheat Genotypes That Develop Both Tan Necrosis and Extensive Chlorosis in Response to Isolates of Pyrenophora tritici-repentis. Plant Dis. 1991, 75, 121–122. [Google Scholar] [CrossRef]
- Singh, P.K.; Crossa, J.; Duveiller, E.; Singh, R.P.; Djurle, A. Association Mapping for Resistance to Tan Spot Induced by Pyrenophora tritici-repentis Race 1 in CIMMYTs Historical Bread Wheat Set. Euphytica 2016, 207, 515–525. [Google Scholar] [CrossRef]
- Singh, P.K.; Gonzalez-Hernandez, J.L.; Mergoum, M.; Ali, S.; Adhikari, T.B.; Kianian, S.F.; Elias, E.M.; Hughes, G.R. Identification and Molecular Mapping of a Gene Conferring Resistance to Pyrenophora Tritici-Repentis Race 3 in Tetraploid Wheat. Phytopathology 2006, 96, 885–889. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Bockus, W.W.; Sharma, I.; Bowden, R.L. A Novel Source of Resistance in Wheat to Pyrenophora tritici-repentis Race 1. Plant Dis. 2008, 92, 91–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.K.; Duveiller, E.; Singh, R.P. Resistance Breeding for Tan Spot (Pyrenophora tritici-repentis) of Wheat. In Disease Resistance in Wheat; Sharma, I., Ed.; CABI: Wallingford, UK, 2012; pp. 136–150. [Google Scholar]
- Chu, C.-G.; Friesen, T.L.; Xu, S.S.; Faris, J.D. Identification of Novel Tan Spot Resistance Loci beyond the Known Host-Selective Toxin Insensitivity Genes in Wheat. Theor. Appl. Genet. 2008, 117, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Faris, J.D.; Overlander, M.E.; Kariyawasam, G.K.; Carter, A.; Xu, S.S.; Liu, Z. Identification of a Major Dominant Gene for Race-Nonspecific Tan Spot Resistance in Wild Emmer Wheat. Theor. Appl. Genet. 2020, 133, 829–841. [Google Scholar] [CrossRef]
- Friesen, T.L.; Faris, J.D. Molecular Mapping of Resistance to Pyrenophora tritici-repentis Race 5 and Sensitivity to Ptr ToxB in Wheat. Theor. Appl. Genet. 2004, 109, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Tadesse, W.; Hsam, S.L.K.; Wenzel, G.; Zeller, F.J. Chromosome Location of a Gene Conferring Resistance to Pyrenophora tritici-repentis in Ethiopian Wheat Cultivars. Euphytica 2008, 162, 423–430. [Google Scholar] [CrossRef]
- Tadesse, W.; Schmolke, M.; Hsam, S.L.K.; Mohler, V.; Wenzel, G.; Zeller, F.J. Molecular Mapping of Resistance Genes to Tan Spot [Pyrenophora tritici-repentis Race 1] in Synthetic Wheat Lines. Theor. Appl. Genet. 2007, 114, 855–862. [Google Scholar] [CrossRef]
- Tadesse, W.; Hsam, S.L.; Wenzel, G.; Zeller, F.J. Identification and Monosomic Analysis of Tan Spot Resistance Genes in Synthetic Wheat Lines (Triticum turgidum L. Aegilops Tauschii Coss.). Crop Sci. 2006, 46, 1212–1217. [Google Scholar] [CrossRef]
- Chu, C.-G.; Chao, S.; Friesen, T.L.; Faris, J.D.; Zhong, S.; Xu, S.S. Identification of Novel Tan Spot Resistance QTLs Using an SSR-Based Linkage Map of Tetraploid Wheat. Mol. Breed. 2010, 25, 327–338. [Google Scholar] [CrossRef]
- Muqaddasi, Q.H.; Kamal, R.; Mirdita, V.; Rodemann, B.; Ganal, M.W.; Reif, J.C.; Röder, M.S. Genome-Wide Association Studies and Prediction of Tan Spot (Pyrenophora Tritici-Repentis) Infection in European Winter Wheat via Different Marker Platforms. Genes 2021, 12, 490. [Google Scholar] [CrossRef]
- Kokhmetova, A.; Sehgal, D.; Ali, S.; Atishova, M.; Kumarbayeva, M.; Leonova, I.; Dreisigacker, S. Genome-Wide Association Study of Tan Spot Resistance in a Hexaploid Wheat Collection From Kazakhstan. Front. Genet. 2021, 11, 581214. [Google Scholar] [CrossRef]
- Li, H.B.; Yan, W.; Liu, G.R.; Wen, S.M.; Liu, C.J. Identification and Validation of Quantitative Trait Loci Conferring Tan Spot Resistance in the Bread Wheat Variety Ernie. Theor. Appl. Genet. 2011, 122, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Kollers, S.; Rodemann, B.; Ling, J.; Korzun, V.; Ebmeyer, E.; Argillier, O.; Hinze, M.; Plieske, J.; Kulosa, D.; Ganal, M.W.; et al. Genome-Wide Association Mapping of Tan Spot Resistance (Pyrenophora tritici-repentis) in European Winter Wheat. Mol. Breed. 2014, 34, 363–371. [Google Scholar] [CrossRef]
- Billiard, S.; López-Villavicencio, M.; Hood, M.E.; Giraud, T. Sex, Outcrossing and Mating Types: Unsolved Questions in Fungi and beyond: Sexy Fungi. J. Evol. Biol. 2012, 25, 1020–1038. [Google Scholar] [CrossRef] [PubMed]
- Billiard, S.; López-Villavicencio, M.; Devier, B.; Hood, M.E.; Fairhead, C.; Giraud, T. Having Sex, Yes, but with Whom? Inferences from Fungi on the Evolution of Anisogamy and Mating Types. Biol. Rev. 2011, 86, 421–442. [Google Scholar] [CrossRef] [PubMed]
- Tollenaere, C.; Laine, A.-L. Investigating the Production of Sexual Resting Structures in a Plant Pathogen Reveals Unexpected Self-Fertility and Genotype-by-Environment Effects. J. Evol. Biol. 2013, 26, 1716–1726. [Google Scholar] [CrossRef] [PubMed]
- Laine, A.-L.; Hanski, I. Large-Scale Spatial Dynamics of a Specialist Plant Pathogen in a Fragmented Landscape. J. Ecol. 2006, 94, 217–226. [Google Scholar] [CrossRef]
- Schnathorst, W.C. Environmental Relationships in the Powdery Mildews. Annu. Rev. Phytopathol. 1965, 3, 343–366. [Google Scholar] [CrossRef]
- Gadoury, D.M.; Wakefield, L.M.; Cadle-Davidson, L.; Dry, I.B.; Seem, R.C. Effects of Prior Vegetative Growth, Inoculum Density, Light, and Mating on Conidiation of Erysiphe necator. Phytopathology 2012, 102, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Fakhfakh, M.; (Comptoir Multiservices Agricoles, Tunis, Tunisia). Personal communication, 2019.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reaction Types 1 | rAUDPC Range | Symptom’s Description |
|---|---|---|
| R | <0.5 | small apparent infection at low leaves |
| MR | 0.5–0.7 | Midheight infection and apparent lesions not well developed |
| MS | 0.7–0.9 | Well-developed lesions up to F−1 * |
| S | >0.9 | Well-developed lesions up to flag leaf |
| Variables | rAUDPC 2019 | PH 2021 |
|---|---|---|
| Seedling 2019 | r = 0.289 p =4.685 × 10−11 *** | - |
| PH 2019 | r = −0.086 p = 0.045 * | r = 0.605 p < 2.2 × 10−16 *** |
| rAUDPC 2021 | r = 0.531 p < 2.2 × 10−16 *** | r = −0.347 p < 2.2 × 10−16 *** |
| Physiological Stage | Source of Variation | df 1 | Sum of Squares | Mean of Squares | F Value | Pr (>F) |
|---|---|---|---|---|---|---|
| Adult | Year | 1 | 7.8 | 7.755 | 9.064 | 0.00267 ** |
| residuals | 1096 | 937.7 | 0.856 | |||
| Genotype | 548 | 658.0 | 1.2007 | 2.293 | <2 × 10−16 *** | |
| residuals | 549 | 287.5 | 0.5237 | |||
| Genotype × Year | 547 | 8.722 | 0.01594 | 6.532 | 0.3042 | |
| residuals | 1 | 0.002 | 0.00244 | |||
| Origin | 4 | 150.0 | 37.49 | 51.51 | <2 × 10−16 *** | |
| Residuals | 1093 | 795.5 | 0.73 | |||
| Level of improvement | 4 | 60.9 | 15.223 | 18.9 | 5.02 × 10−15 *** | |
| Residuals | 1071 | 862.8 | 0.806 | |||
| PH | 27 | 3.99 | 0.14796 | 4.98 | 4.19 × 10−15 *** | |
| Residuals | 1070 | 31.79 | 0.02971 | |||
| Seedling | 3 | 41.4 | 13.794 | 14.21 | 6.7 × 10−9 *** | |
| Residuals | 503 | 488.4 | 0.971 |
| Level of Improvement | % R-MR Seedling 2019 | % R-MR Adult 2019 | % R-MR Adult 2021 | % R-MR Adult 2019 and 2021 |
|---|---|---|---|---|
| Breeding material | 50 | 56 | 52 | 39 |
| Cultivar | 30 | 32 | 25 | 18 |
| Genetic material | 69 | 92 | 92 | 85 |
| Landrace | 61 | 65 | 65 | 49 |
| Unknown improvement status | 73 | 67 | 51 | 37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laribi, M.; Yahyaoui, A.H.; Abdedayem, W.; Kouki, H.; Sassi, K.; Ben M’Barek, S. Characterization of Mediterranean Durum Wheat for Resistance to Pyrenophora tritici-repentis. Genes 2022, 13, 336. https://doi.org/10.3390/genes13020336
Laribi M, Yahyaoui AH, Abdedayem W, Kouki H, Sassi K, Ben M’Barek S. Characterization of Mediterranean Durum Wheat for Resistance to Pyrenophora tritici-repentis. Genes. 2022; 13(2):336. https://doi.org/10.3390/genes13020336
Chicago/Turabian StyleLaribi, Marwa, Amor Hassine Yahyaoui, Wided Abdedayem, Hajer Kouki, Khaled Sassi, and Sarrah Ben M’Barek. 2022. "Characterization of Mediterranean Durum Wheat for Resistance to Pyrenophora tritici-repentis" Genes 13, no. 2: 336. https://doi.org/10.3390/genes13020336
APA StyleLaribi, M., Yahyaoui, A. H., Abdedayem, W., Kouki, H., Sassi, K., & Ben M’Barek, S. (2022). Characterization of Mediterranean Durum Wheat for Resistance to Pyrenophora tritici-repentis. Genes, 13(2), 336. https://doi.org/10.3390/genes13020336