
Transcriptomics of Wet Skin Biopsies Predict Early Radiation-Induced Hematological Damage in a Mouse Model

 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Ethics
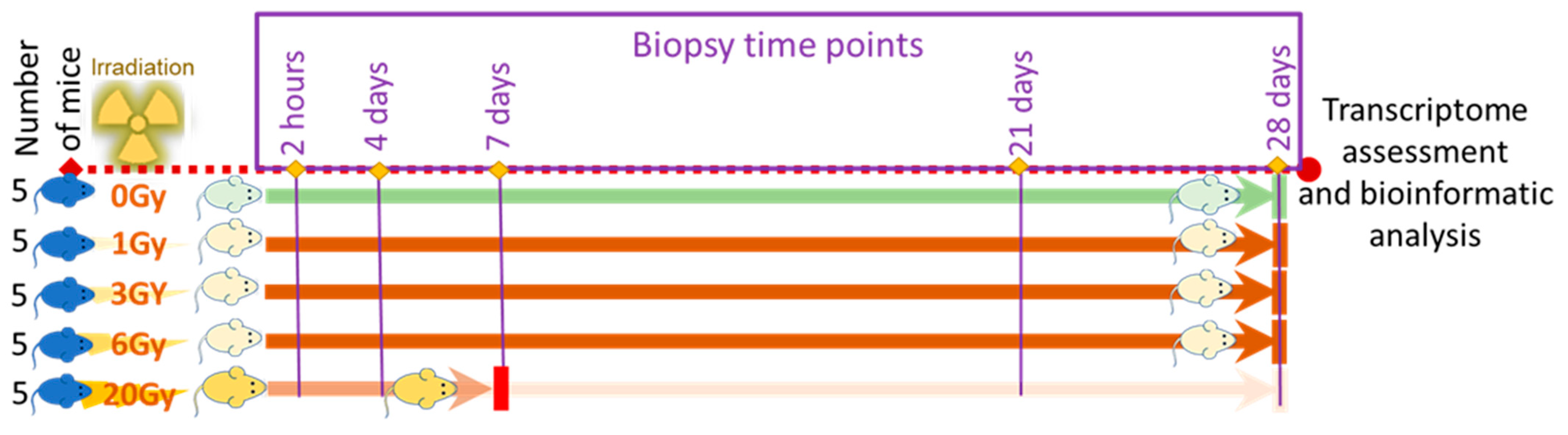
2.2. Animal Preparation and Radiation Treatment
2.2.1. Radiation Treatment
2.2.2. Sample Collection and Post-Irradiation Observation
2.3. Microarrays and Data Preparation
2.3.1. RNA Extraction
2.3.2. Microarrays
2.3.3. Data Preparation and Analysis
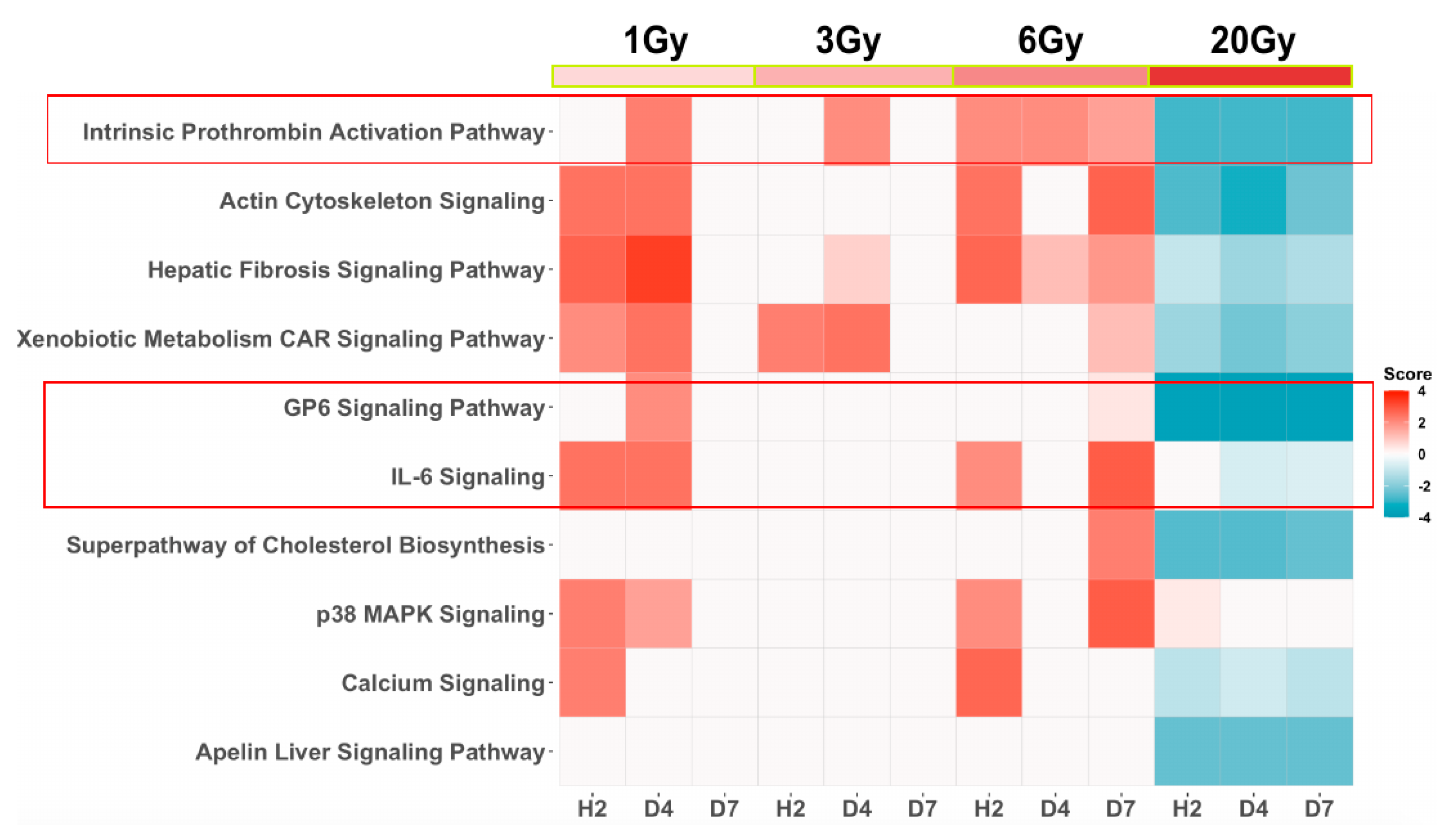
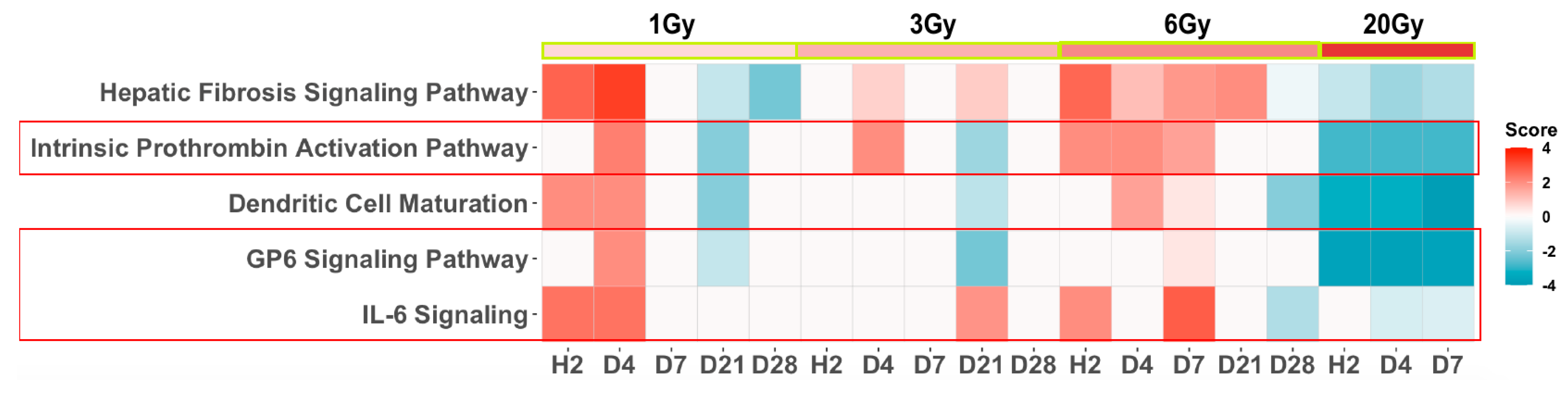
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Disclaimer
References
- Ledney, G.D.; Elliott, T.B. Combined injury: Factors with potential to impact radiation dose assessments. Health Phys. 2010, 98, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Howard, A.; West, R.; Iball, G.; Panteli, M.; Pandit, H.; Giannoudis, P.V. An Estimation of Lifetime Fatal Carcinogenesis Risk Attributable to Radiation Exposure in the First Year Following Polytrauma: A Major Trauma Center’s Experience over 10 Years. JBJS 2019, 101, 1375–1380. [Google Scholar] [CrossRef] [PubMed]
- DiCarlo, A.L.; Hatchett, R.J.; Kaminski, J.M.; Ledney, G.D.; Pellmar, T.C.; Okunieff, P.; Ramakrishnan, N. Medical Countermeasures for Radiation Combined Injury: Radiation with Burn, Blast, Trauma and/or Sepsis. Report of an NIAID Workshop, 26–27 March 2007. Radiat. Res. 2008, 169, 712–721. [Google Scholar] [CrossRef] [PubMed]
- Anno, G.H.; Baum, S.J.; Withers, H.R.; Young, R.W. Symptomatology of acute radiation effects in humans after exposure to doses of 0.5–30 Gy. Health Phys. 1989, 56, 821–838. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, J.M.; Prasanna, P.G.S.; Grace, M.B.; Wathen, L.K.; Wallace, R.L.; Koerner, J.F.; Coleman, C.N. Assessment of biodosimetry methods for a mass-casualty radiological incident: Medical response and management considerations. Health Phys. 2013, 105, 540–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karellas, A. Challenges in Dosimetry and Radiation Dose Trends. Radiology 2020, 294, 360–361. [Google Scholar] [CrossRef] [PubMed]
- Flood, A.B.; Ali, A.N.; Boyle, H.K.; Du, G.; Satinsky, V.A.; Swarts, S.G.; Williams, B.B.; Demidenko, E.; Schreiber, W.; Swartz, H.M. Evaluating the Special Needs of The Military for Radiation Biodosimetry for Tactical Warfare Against Deployed Troops: Comparing Military to Civilian Needs for Biodosimetry Methods. Health Phys. 2016, 111, 169–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, A.R.; Guyon, T.; Headley, V.; Nair, M.; Ricketts, W.; Gray, G.; Wong, J.Y.C.; Chao, N.; Terbrueggen, R. Role of a high throughput biodosimetry test in treatment prioritization after a nuclear incident. Int. J. Radiat. Biol. 2020, 96, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Wathen, L.K.; Eder, P.S.; Horwith, G.; Wallace, R.L. Using biodosimetry to enhance the public health response to a nuclear incident. Int. J. Radiat. Biol. 2020, 97, S6–S9. [Google Scholar] [CrossRef]
- Thrall, K.D.; Mahendra, S.; Lovaglio, J.; Jackson, M.K. The impact of supportive care on survival in large animal models of total body irradiation. Int. J. Radiat. Biol. 2021, 97, S88–S99. [Google Scholar] [CrossRef] [PubMed]
- Coleman, C.N.; Koerner, J.F. Biodosimetry: Medicine, Science, and Systems to Support the Medical Decision-Maker Following a Large Scale Nuclear or Radiation Incident. Radiat. Prot. Dosim. 2016, 172, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Garty, G.; Amundson, S.; Laiakis, E.; Fornace, A.; Brenner, D. High throughput biodosimetry methods. In Radiobiology and Methods Public Textbook; Chapter XXXII; University of Pittsburgh: Pittsburgh, PA, USA; Centers for Medical Countermeasures against Radiation, National Institute of Allergy and Infectious Disease: Bethesda, MD, USA, 2017; pp. 1–18. [Google Scholar]
- Vral, A.; Fenech, M.; Thierens, H. The micronucleus assay as a biological dosimeter of in vivo ionising radiation exposure. Mutagenesis 2011, 26, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Fenech, M.; Morley, A.A. Cytokinesis-block micronucleus method in human lymphocytes: Effect of in vivo ageing and low dose X-irradiation. Mutat. Res. 1986, 161, 193–198. [Google Scholar] [CrossRef]
- Balajee, A.S.; Bertucci, A.; Taveras, M.; Brenner, D.J. Multicolour FISH analysis of ionising radiation induced micronucleus formation in human lymphocytes. Mutagenesis 2014, 29, 447–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertucci, A.; Smilenov, L.B.; Turner, H.C.; Amundson, S.A.; Brenner, D.J. In vitro RABiT measurement of dose rate effects on radiation induction of micronuclei in human peripheral blood lymphocytes. Radiat Environ. Biophys. 2016, 55, 53–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghandhi, S.A.; Shuryak, I.; Morton, S.R.; Amundson, S.A.; Brenner, D.J. New Approaches for Quantitative Reconstruction of Radiation Dose in Human Blood Cells. Sci. Rep. 2019, 9, 18441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorobiev, A.I. Acute radiation disease and biological dosimetry in 1993. Stem Cells 1997, 15 (Suppl. 2), 269–274. [Google Scholar] [CrossRef] [PubMed]
- Demidenko, E.; Williams, B.B.; Swartz, H.M. Radiation dose prediction using data on time to emesis in the case of nuclear terrorism. Radiat. Res. 2009, 171, 310–319. [Google Scholar] [CrossRef]
- American Academy of Pediatrics; American College of Emergency Physicians; American College of Surgeons—Committee on Trauma; American Trauma Society; Children’s National Medical Center, Child Health Advocacy Institute, Emergency Medical Services for Children National Resource Center; International Association of Emergency Medical Services Chiefs; National Association of County and City Health Officials; National Association of Emergency Medical Technicians; National Association of EMS Physicians; National Association of State EMS Officials; et al. Model uniform core criteria for mass casualty triage. Disaster Med. Public Health Prep. 2011, 5, 125–128. [Google Scholar] [CrossRef]
- SALT mass casualty triage: Concept endorsed by the American College of Emergency Physicians, American College of Surgeons Committee on Trauma, American Trauma Society, National Association of EMS Physicians, National Disaster Life Support Education Consortium, and State and Territorial Injury Prevention Directors Association. Disaster Med. Public Health Prep. 2008, 2, 245–246.
- Lerner, E.B.; Schwartz, R.B.; Coule, P.L.; Weinstein, E.S.; Cone, D.C.; Hunt, R.C.; Sasser, S.M.; Liu, J.M.; Nudell, N.G.; Wedmore, I.S.; et al. Mass casualty triage: An evaluation of the data and development of a proposed national guideline. Disaster Med. Public Health Prep. 2008, 2 (Suppl. 1), S25–S34. [Google Scholar] [CrossRef] [PubMed]
- Bussel, J.B.; Soff, G.; Balduzzi, A.; Cooper, N.; Lawrence, T.; Semple, J.W. A Review of Romiplostim Mechanism of Action and Clinical Applicability. Drug Des. Dev. Ther. 2021, 15, 2243–2268. [Google Scholar] [CrossRef] [PubMed]
- Park, J.G.; Paul, S.; Briones, N.; Zeng, J.; Gillis, K.; Wallstrom, G.; LaBaer, J.; Amundson, S.A. Developing Human Radiation Biodosimetry Models: Testing Cross-Species Conversion Approaches Using an Ex Vivo Model System. Radiat. Res. 2017, 187, 708–721. [Google Scholar] [CrossRef] [PubMed]
- Macaeva, E.; Saeys, Y.; Tabury, K.; Janssen, A.; Michaux, A.; Benotmane, M.A.; De Vos, W.H.; Baatout, S.; Quintens, R. Radiation-induced alternative transcription and splicing events and their applicability to practical biodosimetry. Sci. Rep. 2016, 6, 19251. [Google Scholar] [CrossRef] [PubMed]
- Forrester, H.B.; Li, J.; Hovan, D.; Ivashkevich, A.N.; Sprung, C.N. DNA repair genes: Alternative transcription and gene expression at the exon level in response to the DNA damaging agent, ionizing radiation. PLoS ONE 2012, 7, e53358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quintens, R.; Verreet, T.; Janssen, A.; Neefs, M.; Leysen, L.; Michaux, A.; Verslegers, M.; Samari, N.; Pani, G.; Verheyde, J.; et al. Identification of novel radiation-induced p53-dependent transcripts extensively regulated during mouse brain development. Biol. Open 2015, 4, 331–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprung, C.N.; Li, J.; Hovan, D.; McKay, M.J.; Forrester, H.B. Alternative transcript initiation and splicing as a response to DNA damage. PLoS ONE 2011, 6, e25758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghandhi, S.A.; Smilenov, L.B.; Elliston, C.D.; Chowdhury, M.; Amundson, S.A. Radiation dose-rate effects on gene expression for human biodosimetry. BMC Med. Genom. 2015, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broustas, C.G.; Xu, Y.; Harken, A.D.; Chowdhury, M.; Garty, G.; Amundson, S.A. Impact of Neutron Exposure on Global Gene Expression in a Human Peripheral Blood Model. Radiat. Res. 2017, 187, 433–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S.; Amundson, S.A. Development of gene expression signatures for practical radiation biodosimetry. Int. J. Radiat. Oncol. Biol. Phys. 2008, 71, 1236–1244. [Google Scholar] [CrossRef] [Green Version]
- Doyle-Eisele, M.; Brower, J.; Aiello, K.; Ferranti, E.; Yaeger, M.; Wu, G.; Weber, W. Developing and comparing models of hematopoietic-acute radiation syndrome in Göttingen and Sinclair minipigs. Int. J. Radiat. Biol. 2020, 97, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Jackson, I.L.; Gurung, G.; Poirier, Y.; Gopalakrishnan, M.; Cohen, E.P.; Donohue, T.-S.; Newman, D.; Vujaskovic, Z. A New Zealand White rabbit model of thrombocytopenia and coagulopathy following total body irradiation across the dose range to induce the hematopoietic-subsyndrome of acute radiation syndrome. Int. J. Radiat. Biol. 2021, 97 (Suppl. 1), S19–S31. [Google Scholar] [CrossRef] [PubMed]
- Amundson, S.A.; Grace, M.B.; McLeland, C.B.; Epperly, M.W.; Yeager, A.; Zhan, Q.; Greenberger, J.S.; Fornace, A.J., Jr. Human in vivo radiation-induced biomarkers: Gene expression changes in radiotherapy patients. Cancer Res. 2004, 64, 6368–6371. [Google Scholar] [CrossRef] [Green Version]
- Heath, W.R.; Carbone, F.R. The skin-resident and migratory immune system in steady state and memory: Innate lymphocytes, dendritic cells and T cells. Nat. Immunol. 2013, 14, 978–985. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, C. Role of the microenvironment on hematopoiesis. I. Stem cell differentiation into granulocytic and megakaryocytic cell lineage. J. Lab. Clin. Med. 1989, 114, 378–381. [Google Scholar] [PubMed]
- Yoder, M.C.; Williams, D.A. Matrix molecule interactions with hematopoietic stem cells. Exp. Hematol. 1995, 23, 961–967. [Google Scholar] [PubMed]
- Katagiri, T.; Uemura, S.; Ushiki, T.; Nakajima-Takagi, Y.; Oshima, M.; Mikami, T.; Kawasaki, A.; Ishiguro, H.; Tanaka, T.; Sone, H.; et al. Distinct effects of chondroitin sulfate on hematopoietic cells and the stromal microenvironment in bone marrow hematopoiesis. Exp. Hematol. 2021, 96, 52–62.e5. [Google Scholar] [CrossRef] [PubMed]
- Geutskens, S.B.; Andrews, W.D.; van Stalborch, A.M.; Brussen, K.; Holtrop-de Haan, S.E.; Parnavelas, J.G.; Hordijk, P.L.; van Hennik, P.B. Control of human hematopoietic stem/progenitor cell migration by the extracellular matrix protein Slit3. Lab. Investig. 2012, 92, 1129–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sackstein, R. The Bone Marrow Is Akin to Skin: HCELL and the Biology of Hematopoietic Stem Cell Homing11Reprinted from J Invest Dermatol 122:1061-1069, 2004. J. Investig. Dermatol. Symp. Proc. 2004, 9, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Immunity in the tissues. Nat. Immunol. 2013, 14, 977. [CrossRef] [PubMed] [Green Version]
- i Garau, M.M.; Lucas Calduch, A.; López, E.C. Radiobiology of the acute radiation syndrome. Rep. Pract. Oncol. Radiother. 2011, 16, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Barabanova, A.V.; Bushmanov, A.J.; Kotenko, K.V. Acute Radiation Sickness From Chernobyl. In Reference Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2019. [Google Scholar]
- Khalil, S.; Ariel Gru, A.; Saavedra, A.P. Cutaneous extramedullary haematopoiesis: Implications in human disease and treatment. Exp. Dermatol. 2019, 28, 1201–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Au-Lei, B.U.W.; Au-Yamada, M.; Au-Hoang, V.L.T.; Au-Belt, P.J.; Au-Moore, M.H.; Au-Lin, L.L.; Au-Flewell-Smith, R.; Au-Dang, N.; Au-Tomihara, S.; Au-Prow, T.W. Absorbent Microbiopsy Sampling and RNA Extraction for Minimally Invasive, Simultaneous Blood and Skin Analysis. JoVE 2019, e58614. [Google Scholar] [CrossRef]
- DiCarlo, A.L.; Perez Horta, Z.; Rios, C.I.; Satyamitra, M.M.; Taliaferro, L.P.; Cassatt, D.R. Study logistics that can impact medical countermeasure efficacy testing in mouse models of radiation injury. Int. J. Radiat. Biol. 2021, 97 (Suppl. 1), S151–S167. [Google Scholar] [CrossRef] [PubMed]
- Almond, P.R.; Biggs, P.J.; Coursey, B.M.; Hanson, W.F.; Huq, M.S.; Nath, R.; Rogers, D.W. AAPM’s TG-51 protocol for clinical reference dosimetry of high-energy photon and electron beams. Med. Phys. 1999, 26, 1847–1870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dainiak, N.; Waselenko, J.; Armitage, J.; Macvittie, T.; Farese, A. The hematologist and radiation casualties. Hematol. Educ. Program Am. Soc. Hematol. 2003, 2003, 473–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friesecke, I.; Beyrer, K.; Fliedner, T.M. How to cope with radiation accidents: The medical management. Br. J. Radiol. 2001, 74, 121–122. [Google Scholar] [CrossRef]
- Alkhalil, A.; Clifford, J.L.; Ball, R.; Day, A.; Chan, R.; Carney, B.C.; Miller, S.A.; Campbell, R.; Kumar, R.; Gautam, A.; et al. Blood RNA Integrity is a Direct and Simple Reporter of Radiation Exposure and Prognosis: A Pilot Study. Radiat. Res. 2020, 193, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, K.; Ozasa, K.; Akiba, S.; Niwa, O.; Kodama, K.; Takamura, N.; Zaharieva, E.K.; Kimura, Y.; Wakeford, R. Long-term effects of radiation exposure on health. Lancet 2015, 386, 469–478. [Google Scholar] [CrossRef]
- Mazurek, K.; Siemianowicz, K.; Likus, W.; Pierzchała, E.; Kwiatkowski, R.; Markowski, J. Collagen Type III Metabolism Evaluation in Patients with Malignant Head and Neck Cancer Treated with Radiotherapy. BioMed Res. Int. 2018, 2018, 8702605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Nabout, R.; Martin, M.; Remy, J.; Kern, P.; Robert, L.; Lafuma, C. Collagen synthesis and deposition in cultured fibroblasts from subcutaneous radiation-induced fibrosis. Modification as a function of cell aging. Matrix 1989, 9, 411–420. [Google Scholar] [CrossRef]
- Kim, J.H.; Kolozsvary, A.J.; Jenrow, K.A.; Brown, S.L. Mechanisms of radiation-induced skin injury and implications for future clinical trials. Int. J. Radiat. Biol. 2013, 89, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Kiang, J.G.; Garrison, B.R.; Burns, T.M.; Zhai, M.; Dews, I.C.; Ney, P.H.; Cary, L.H.; Fukumoto, R.; Elliott, T.B.; Ledney, G.D. Wound trauma alters ionizing radiation dose assessment. Cell Biosci. 2012, 2, 20. [Google Scholar] [CrossRef] [Green Version]
- Jackson, S.C.; Odiaman, L.; Card, R.T.; van der Bom, J.G.; Poon, M.C. Suspected collagen disorders in the bleeding disorder clinic: A case-control study. Haemophilia 2013, 19, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.; Odiaman, L.; Brown, M.; Spitzer, C.; Card, R.T.; Poon, M.-C. Prevalence of Suspected Collagen Disorders In the Adult Bleeding Disorder Clinic. Blood 2010, 116, 1399. [Google Scholar] [CrossRef]
- Simões, F.C.; Cahill, T.J.; Kenyon, A.; Gavriouchkina, D.; Vieira, J.M.; Sun, X.; Pezzolla, D.; Ravaud, C.; Masmanian, E.; Weinberger, M.; et al. Macrophages directly contribute collagen to scar formation during zebrafish heart regeneration and mouse heart repair. Nat. Commun. 2020, 11, 600. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, T.; Yamamoto, T.; Tsuchiya, E.; Hongo, H.; Tsuboi, K.; Kudo, A.; Abe, M.; Yoshida, T.; Nagai, T.; Khadiza, N.; et al. Ultrastructural and biochemical aspects of matrix vesicle-mediated mineralization. Jpn. Dent. Sci. Rev. 2017, 53, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Oeffinger, K.C.; Mertens, A.C.; Sklar, C.A.; Kawashima, T.; Hudson, M.M.; Meadows, A.T.; Friedman, D.L.; Marina, N.; Hobbie, W.; Kadan-Lottick, N.S.; et al. Chronic health conditions in adult survivors of childhood cancer. N. Engl. J. Med. 2006, 355, 1572–1582. [Google Scholar] [CrossRef] [PubMed]
- Florin, T.A.; Fryer, G.E.; Miyoshi, T.; Weitzman, M.; Mertens, A.C.; Hudson, M.M.; Sklar, C.A.; Emmons, K.; Hinkle, A.; Whitton, J.; et al. Physical inactivity in adult survivors of childhood acute lymphoblastic leukemia: A report from the childhood cancer survivor study. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1356–1363. [Google Scholar] [CrossRef] [Green Version]
- Guise, T.A. Bone loss and fracture risk associated with cancer therapy. Oncologist 2006, 11, 1121–1131. [Google Scholar] [CrossRef]
- Brown, S.A.; Guise, T.A. Cancer treatment-related bone disease. Crit. Rev. Eukaryot. Gene Expr. 2009, 19, 47–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, N.N.; Habermann, E.B.; Tepper, J.E.; Durham, S.B.; Virnig, B.A. Risk of pelvic fractures in older women following pelvic irradiation. JAMA 2005, 294, 2587–2593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, A.R.; Maity, A.; Sanzari, J.K. A Review of Radiation-Induced Coagulopathy and New Findings to Support Potential Prevention Strategies and Treatments. Radiat. Res. 2016, 186, 121–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takada, S.; Fujimori, S.; Shinozuka, T.; Takada, R.; Mii, Y. Differences in the secretion and transport of Wnt proteins. J. Biochem. 2017, 161, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, A.R.; Lee, C.L.; Oh, P.; Luo, L.; Ma, Y.; Kirsch, D.G. Inhibiting Glycogen Synthase Kinase-3 Mitigates the Hematopoietic Acute Radiation Syndrome in a Sex- and Strain-dependent Manner in Mice. Health Phys. 2020, 119, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Fajardo, L.F.; Berthrong, M. Vascular lesions following radiation. Pathol. Annu. 1988, 23, 297–330. [Google Scholar] [PubMed]
- Yang, E.H.; Marmagkiolis, K.; Balanescu, D.V.; Hakeem, A.; Donisan, T.; Finch, W.; Virmani, R.; Herrman, J.; Cilingiroglu, M.; Grines, C.L.; et al. Radiation-Induced Vascular Disease—A State-of-the-Art Review. Front. Cardiovasc. Med. 2021, 8, 223. [Google Scholar] [CrossRef] [PubMed]
- Mailloux, A.W.; Bai, F.; Zhang, L.; Padron, E.; Yang, L.; Wei, C.; Schmidt, A.; Sokol, L.; Chen, D.-T.; Loughran, T.P., Jr.; et al. Altered Collagen Production by Dysfunctional Mesenchymal Stem Cells Is Linked to T Cell Large Granular Lymphocyte Leukemia Pathophysiology. Blood 2010, 116, 2595. [Google Scholar] [CrossRef]
- Bernardo, M.E.; Fibbe, W.E. Mesenchymal stromal cells and hematopoietic stem cell transplantation. Immunol. Lett. 2015, 168, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Cavallero, S.; Riccobono, D.; Drouet, M.; François, S. MSC-Derived Extracellular Vesicles: New Emergency Treatment to Limit the Development of Radiation-Induced Hematopoietic Syndrome? Health Phys. 2020, 119, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Piryani, S.O.; Jiao, Y.; Kam, A.Y.F.; Liu, Y.; Vo-Dinh, T.; Chen, B.J.; Chao, N.J.; Doan, P.L. Endothelial Cell-Derived Extracellular Vesicles Mitigate Radiation-Induced Hematopoietic Injury. Int. J. Radiat. Oncol. Biol. Phys. 2019, 104, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Han, X.; Zhao, Y.; Xue, X.; Fan, S. Mouse serum protects against total body irradiation-induced hematopoietic system injury by improving the systemic environment after radiation. Free Radic. Biol. Med. 2019, 131, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Fernández, M.; Minguell, J.J. The role of collagen in hematopoiesis. Braz. J. Med. Biol. Res. 1996, 29, 1201–1207. [Google Scholar] [PubMed]
- Marquez Romero, S.; Hettler, F.; Hausinger, R.; Schreck, C.; Landspersky, T.; Henkel, L.; Angerpointner, C.; Demir, I.E.; Schiemann, M.; Bassermann, F.; et al. Secreted factors from mouse embryonic fibroblasts maintain repopulating function of single cultured hematopoietic stem cells. Haematologica 2020, 106, 2633–2640. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, E.; Roberts, D.; Corbo, T.; Jacenko, O. Congenic Mice Confirm That Collagen X Is Required for Proper Hematopoietic Development. PLoS ONE 2010, 5, e9518. [Google Scholar] [CrossRef] [PubMed]
- Grskovic, I.; Kutsch, A.; Frie, C.; Groma, G.; Stermann, J.; Schlötzer-Schrehardt, U.; Niehoff, A.; Moss, S.E.; Rosenbaum, S.; Pöschl, E.; et al. Depletion of annexin A5, annexin A6, and collagen X causes no gross changes in matrix vesicle–mediated mineralization, but lack of collagen X affects hematopoiesis and the Th1/Th2 response. J. Bone Miner. Res. 2012, 27, 2399–2412. [Google Scholar] [CrossRef] [PubMed]
- Jacenko, O.; Roberts, D.W.; Campbell, M.R.; McManus, P.M.; Gress, C.J.; Tao, Z. Linking Hematopoiesis to Endochondral Skeletogenesis through Analysis of Mice Transgenic for Collagen X. Am. J. Pathol. 2002, 160, 2019–2034. [Google Scholar] [CrossRef] [Green Version]
- Stefanini, A.C.B.; da Cunha, B.R.; Henrique, T.; Tajara, E.H. Involvement of Kallikrein-Related Peptidases in Normal and Pathologic Processes. Dis. Markers 2015, 2015, 946572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emami, N.; Diamandis, E.P. New insights into the functional mechanisms and clinical applications of the kallikrein-related peptidase family. Mol. Oncol. 2007, 1, 269–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniels, G. Functions of red cell surface proteins. Vox Sang. 2007, 93, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, K.E.; Park, H.J.; Cho, D. The Role of Erythroid Differentiation Regulator 1 (ERDR1) in the Control of Proliferation and Photodynamic Therapy (PDT) Response. Int. J. Mol. Sci. 2020, 21, 2603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashouri, R.; Fangman, M.; Burris, A.; Ezenwa, M.O.; Wilkie, D.J.; Doré, S. Critical Role of Hemopexin Mediated Cytoprotection in the Pathophysiology of Sickle Cell Disease. Int. J. Mol. Sci. 2021, 22, 6408. [Google Scholar] [CrossRef] [PubMed]
- Castellino, F.J.; Ploplis, V.A. Structure and function of the plasminogen/plasmin system. Thromb. Haemost. 2005, 93, 647–654. [Google Scholar] [PubMed]
- Doumas, S.; Kolokotronis, A.; Stefanopoulos, P. Anti-inflammatory and antimicrobial roles of secretory leukocyte protease inhibitor. Infect. Immun. 2005, 73, 1271–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiang, J.G.; Smith, J.T.; Cannon, G.; Anderson, M.N.; Ho, C.; Zhai, M.; Cui, W.; Xiao, M. Ghrelin, a novel therapy, corrects cytokine and NF-κB-AKT-MAPK network and mitigates intestinal injury induced by combined radiation and skin-wound trauma. Cell Biosci. 2020, 10, 63. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GP6 Signaling | Intrinsic Prothrombin | Symbol | Entrez Gene Name | 1 Gy | 3 Gy | 6 Gy | 20 Gy | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| h2 | d4 | d7 | d21 | d28 | h2 | d4 | d7 | d21 | d28 | h2 | d4 | d7 | d21 | d28 | h2 | d4 | d7 | ||||
| × | LAMB1 | laminin subunit bate 1 | 2.078 | 2.041 | |||||||||||||||||
| × | × | COL1A1 | collagen type I alpha 1 chain | 2.44 | −2.485 | −2.467 | −2.007 | 2.612 | 2.378 | 2.1 | −2.314 | −24.32 | −23.33 | −35.28 | |||||||
| × | × | COL1A2 | collagen type I alpha 2 chain | −2.063 | −2.935 | −22.21 | −20.32 | −31.16 | |||||||||||||
| × | × | COL2A1 | collagen type II alpha 1 chain | −2.093 | 2.306 | 2.587 | −3.441 | −2.971 | −4.35 | ||||||||||||
| × | × | COL3A1 | collagen type III alpha 1 chain | −2.081 | −3.277 | −31.75 | −26.58 | −47.74 | |||||||||||||
| × | COL4A4 | collagen type IV alpha 1 chain | −2.175 | ||||||||||||||||||
| × | COL5A1 | collagen type V alpha 1 chain | −4.282 | −3.514 | −5.265 | ||||||||||||||||
| × | COL5A2 | collagen type V alpha 2 chain | −2.437 | −2.762 | |||||||||||||||||
| × | × | COL5A3 | collagen type V alpha 3 chain | −2.03 | −3.209 | −3.717 | −4.282 | ||||||||||||||
| × | COL6A1 | collagen type VI alpha 1 chain | −2.053 | ||||||||||||||||||
| × | COL6A2 | collagen type VI alpha 2 chain | −2.071 | −2.024 | −2.409 | ||||||||||||||||
| × | COL6A3 | collagen type VI alpha 3 chain | −4.002 | −3.526 | −5.218 | ||||||||||||||||
| × | COL7A1 | collagen type VII alpha 1 chain | 2.143 | −2.741 | −3.039 | −3.194 | |||||||||||||||
| × | COL15A1 | collagen type XV alpha 1 chain | −2.051 | −2.231 | −6.312 | −5.899 | −7.502 | ||||||||||||||
| × | COL16A1 | collagen type XVI alpha 1 chain | −2.501 | −2.24 | −2.314 | ||||||||||||||||
| × | × | COL18A1 | collagen type XVIII alpha 1 chain | −2.287 | −2.378 | −2.218 | |||||||||||||||
| × | KLK7 | kallikrein related peptidase 7 | 2.427 | 2.629 | −2.687 | 2.092 | 2.015 | 3.021 | −3.516 | −2.416 | −2.591 | −2.46 | |||||||||
| × | KLK8 | kallikrein related peptidase 8 | 2.6 | 2.544 | 2.194 | 2.348 | 2.026 | 2.166 | −2.366 | −2.534 | −2.983 | ||||||||||
| × | CALML5 | calmodulin like 5 | 2.045 | 2.438 | −2.683 | 2.995 | −3.383 | −3.302 | −3.33 | −2.989 | |||||||||||
| Gene Name | 1 Gy | 3 Gy | 6 Gy | 20 Gy | Location | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| h2 | d4 | d7 | d21 | d28 | h2 | d4 | d7 | d21 | d28 | h2 | d4 | d7 | d21 | d28 | h2 | d4 | d7 | ||
| Erythroblast membrane associated protein (ERMAP) | 2.86 | 2.88 | 3.07 | Cytoplasm | |||||||||||||||
| Erythroid differentiation regulator 1 (EErdr1) | 2.40 | 2.32 | Other | ||||||||||||||||
| HBA1/HBA2 hemoglobin subunit α 1 and 2 | 2.00 | −2.18 | −2.72 | −3.70 | −6.39 | −4.40 | Extracellular space | ||||||||||||
| Hemoglobin subunit β (HBB) | 3.15 | 2.31 | −2.12 | 2.35 | −2.83 | −3.01 | −3.41 | −6.06 | −4.87 | Cytoplasm | |||||||||
| Hemopexin (HPX) | 2.22 | Extracellular space | |||||||||||||||||
| Hemicentin 2 (HMCN2) | 2.41 | −2.79 | −2.70 | −2.73 | Cytoplasm | ||||||||||||||
| Heme binding protein 2 (HEBP2) | 2.19 | 2.26 | 2.03 | 2.04 | 2.78 | 2.54 | 2.31 | 3.46 | −2.15 | −2.06 | −2.33 | Extracellular space | |||||||
| Hemojuvelin BMP coreceptor (HJV) | −2.40 | Plasma Membrane | |||||||||||||||||
| Secretory leukocyte peptidase inhibitor (SLP1) | 2.01 | 2.19 | 4.73 | 4.00 | 3.99 | Cytoplasm | |||||||||||||
| Plasminogen (PLG) | 2.26 | 2.28 | 2.36 | 2.51 | Extracellular space | ||||||||||||||
| Leukocyte immunoglobulin like receptor B3 (LILRB3) | 2.01 | −2.90 | −2.36 | −2.85 | Plasma Membrane | ||||||||||||||
| Platelet factor 4 | 2.62 | −2.36 | |||||||||||||||||
| Platelet derived growth factor like | −2.53 | 2.03 | −3.14 | −2.33 | −3.67 | Plasma Membrane | |||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alkhalil, A.; Clifford, J.; Miller, S.A.; Gautam, A.; Jett, M.; Hammamieh, R.; Moffatt, L.T.; Shupp, J.W. Transcriptomics of Wet Skin Biopsies Predict Early Radiation-Induced Hematological Damage in a Mouse Model. Genes 2022, 13, 538. https://doi.org/10.3390/genes13030538
Alkhalil A, Clifford J, Miller SA, Gautam A, Jett M, Hammamieh R, Moffatt LT, Shupp JW. Transcriptomics of Wet Skin Biopsies Predict Early Radiation-Induced Hematological Damage in a Mouse Model. Genes. 2022; 13(3):538. https://doi.org/10.3390/genes13030538
Chicago/Turabian StyleAlkhalil, Abdulnaser, John Clifford, Stacy Ann Miller, Aarti Gautam, Marti Jett, Rasha Hammamieh, Lauren T. Moffatt, and Jeffrey W. Shupp. 2022. "Transcriptomics of Wet Skin Biopsies Predict Early Radiation-Induced Hematological Damage in a Mouse Model" Genes 13, no. 3: 538. https://doi.org/10.3390/genes13030538
APA StyleAlkhalil, A., Clifford, J., Miller, S. A., Gautam, A., Jett, M., Hammamieh, R., Moffatt, L. T., & Shupp, J. W. (2022). Transcriptomics of Wet Skin Biopsies Predict Early Radiation-Induced Hematological Damage in a Mouse Model. Genes, 13(3), 538. https://doi.org/10.3390/genes13030538