
Feature Fusion and Detection in Alzheimer’s Disease Using a Novel Genetic Multi-Kernel SVM Based on MRI Imaging and Gene Data



Abstract
:1. Introduction
2. Materials and Methods
2.1. Imaging and Gene Data
2.2. Feature Construction
2.3. Genetic Multi-Kernel SVM Construction
2.4. Gene Identification and Biological Significance Assessment
3. Results
3.1. Results of Parameter Optimization
3.2. Comparison with Other Methods
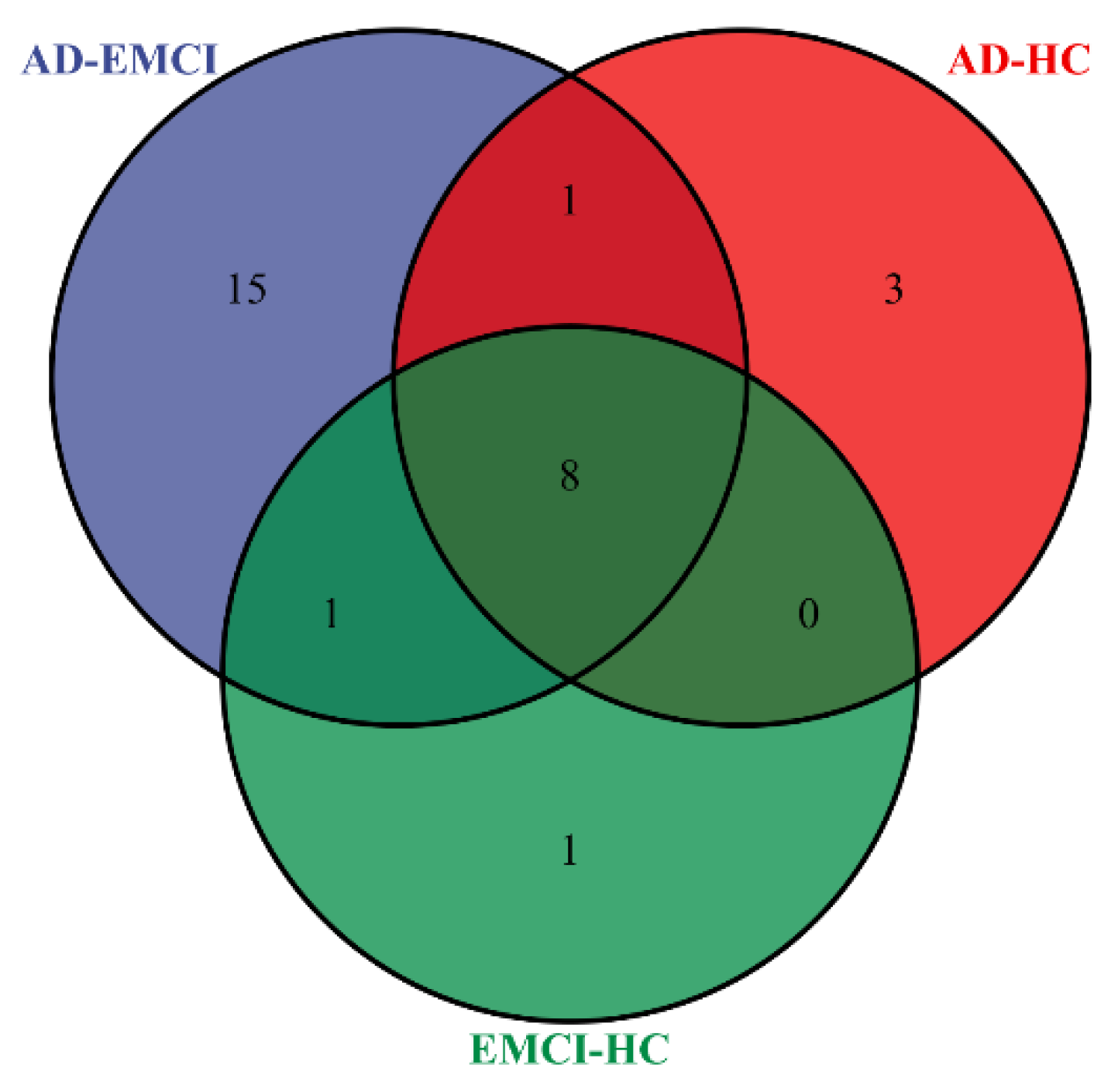
3.3. Identification of Brain Regions and Genes
3.4. Biological Significance Assessment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newton-Cheh, C.; Hirschhorn, J.N. Genetic association studies of complex traits: Design and analysis issues. Mutat. Res. /Fundam. Mol. Mech. Mutagenesis 2005, 573, 54–69. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Thompson, P.; Wang, Y.; Yu, Y.; Zhang, J.; Kong, D.; Colen, R.R.; Knickmeyer, R.C.; Zhu, H.; Alzheimer’s Disease Neuroimaging, I. FGWAS: Functional genome wide association analysis. NeuroImage 2017, 159, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Stein, J.L.; Hua, X.; Lee, S.; Ho, A.J.; Leow, A.D.; Toga, A.W.; Saykin, A.J.; Shen, L.; Foroud, T.; Pankratz, N.; et al. Voxelwise genome-wide association study (vGWAS). NeuroImage 2010, 53, 1160–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibar, D.P.; Stein, J.L.; Kohannim, O.; Jahanshad, N.; Saykin, A.J.; Shen, L.; Kim, S.; Pankratz, N.; Foroud, T.; Huentelman, M.J.; et al. Voxelwise gene-wide association study (vGeneWAS): Multivariate gene-based association testing in 731 elderly subjects. NeuroImage 2011, 56, 1875–1891. [Google Scholar] [CrossRef] [Green Version]
- Vounou, M.; Nichols, T.E.; Montana, G.; Alzheimer’s Disease Neuroimaging, I. Discovering genetic associations with high-dimensional neuroimaging phenotypes: A sparse reduced-rank regression approach. NeuroImage 2010, 53, 1147–1159. [Google Scholar] [CrossRef] [Green Version]
- Rajeesh, J.; Moni, R.S.; Gopalakrishnan, T. Discrimination of Alzheimer’s disease using hippocampus texture features from MRI. Asian Biomed. 2012, 6, 87–94. [Google Scholar]
- Guenther, F.; Brandl, C.; Winkler, T.W.; Wanner, V.; Stark, K.; Kuechenhoff, H.; Heid, I.M. Chances and challenges of machine learning-based disease classification in genetic association studies illustrated on age-related macular degeneration. Genet. Epidemiol. 2020, 44, 759–777. [Google Scholar] [CrossRef]
- Seo, D.; Cho, S.; Manjula, P.; Choi, N.; Kim, Y.-K.; Koh, Y.J.; Lee, S.H.; Kim, H.-Y.; Lee, J.H. Identification of Target Chicken Populations by Machine Learning Models Using the Minimum Number of SNPs. Animals 2021, 11, 241. [Google Scholar] [CrossRef]
- Li, Y.; Haber, A.; Preuss, C.; John, C.; Uyar, A.; Yang, H.S.; Logsdon, B.A.; Philip, V.; Karuturi, R.K.M.; Carter, G.W.; et al. Transfer learning-trained convolutional neural networks identify novel MRI biomarkers of Alzheimer’s disease progression. Alzheimer’s Dement. Diagn. Assess. Dis. Monit. 2021, 13, e12140. [Google Scholar] [CrossRef]
- Huang, W.; Li, X.; Li, X.; Kang, G.; Han, Y.; Shu, N. Combined Support Vector Machine Classifier and Brain Structural Network Features for the Individual Classification of Amnestic Mild Cognitive Impairment and Subjective Cognitive Decline Patients. Front. Aging Neurosci. 2021, 13, 687927. [Google Scholar] [CrossRef]
- Díez Díaz, F.; Sánchez Lasheras, F.; Moreno, V.; Moratalla-Navarro, F.; Molina De La Torre, A.J.; Martín Sánchez, V. GASVeM: A New Machine Learning Methodology for Multi-SNP Analysis of GWAS Data Based on Genetic Algorithms and Support Vector Machines. Mathematics 2021, 9, 654. [Google Scholar] [CrossRef]
- Kinreich, S.; Meyers, J.L.; Maron-Katz, A.; Kamarajan, C.; Pandey, A.K.; Chorlian, D.B.; Zhang, J.; Pandey, G.; Subbie-Saenz De Viteri, S.; Pitti, D.; et al. Predicting risk for Alcohol Use Disorder using longitudinal data with multimodal biomarkers and family history: A machine learning study. Mol. Psychiatry 2021, 26, 1133–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brabec, J.L.; Lara, M.K.; Tyler, A.L.; Mahoney, J.M. System-Level Analysis of Alzheimer’s Disease Prioritizes Candidate Genes for Neurodegeneration. Front. Genet. 2021, 12, 625246. [Google Scholar] [CrossRef]
- Matthews, P.M.; Hampshire, A. Clinical Concepts Emerging from fMRI Functional Connectomics. Neuron 2016, 91, 511–528. [Google Scholar] [CrossRef] [Green Version]
- Amunts, K.; Mohlberg, H.; Bludau, S.; Zilles, K. Julich-Brain: A 3D probabilistic atlas of the human brain’s cytoarchitecture. Science 2020, 369, 988–992. [Google Scholar] [CrossRef]
- Yao, X.; Cong, S.; Yan, J.; Risacher, S.L.; Saykin, A.J.; Moore, J.H.; Shen, L. Regional imaging genetic enrichment analysis. Bioinformatics 2020, 36, 2554–2560. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Risacher, S.L.; Nho, K.; Saykin, A.J.; Wang, Z.; Shen, L. Targeted genetic analysis of cerebral blood flow imaging phenotypes implicates the INPP5D gene. Neurobiol. Aging 2019, 81, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saykin, A.J.; Shen, L.; Foroud, T.M.; Potkin, S.G.; Swaminathan, S.; Kim, S.; Risacher, S.L.; Nho, K.; Huentelman, M.J.; Craig, D.W.; et al. Alzheimer’s Disease Neuroimaging Initiative biomarkers as quantitative phenotypes: Genetics core aims, progress, and plans. Alzheimer’s Dement. 2010, 6, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Lambert, J.-C.; Ibrahim-Verbaas, C.A.; Harold, D.; Naj, A.C.; Sims, R.; Bellenguez, C.; Jun, G.; DeStefano, A.L.; Bis, J.C.; Beecham, G.W.; et al. Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer’s disease. Nat. Genet. 2013, 45, 1452–1458. [Google Scholar] [CrossRef] [Green Version]
- Hinrichs, C.; Singh, V.; Peng, J.; Johnson, S.C. Q-MKL: Matrix-induced Regularization in Multi-Kernel Learning with Applications to Neuroimaging. Adv. Neural Inf. Process. Syst. 2012, 2012, 1430–1438. [Google Scholar] [PubMed]
- Peng, Z.; Hu, Q.; Dang, J. Multi-kernel SVM based depression recognition using social media data. Int. J. Mach. Learn. Cybern. 2019, 10, 43–57. [Google Scholar] [CrossRef]
- Cortes, C.; Vapnik, V. Support-Vector Networks. Mach. Learn. 1995, 20, 273–297. [Google Scholar] [CrossRef]
- Li, M.; Jiang, L.; Mak, T.S.H.; Kwan, J.S.H.; Xue, C.; Chen, P.; Leung, H.C.-M.; Cui, L.; Li, T.; Sham, P.C. A powerful conditional gene-based association approach implicated functionally important genes for schizophrenia. Bioinformatics 2019, 35, 628–635. [Google Scholar] [CrossRef]
- Bu, D.; Luo, H.; Huo, P.; Wang, Z.; Zhang, S.; He, Z.; Wu, Y.; Zhao, L.; Liu, J.; Guo, J.; et al. KOBAS-i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef] [PubMed]
- Parcerisas, A.; Rubio, S.E.; Muhaisen, A.; Gómez-Ramos, A.; Pujadas, L.; Puiggros, M.; Rossi, D.; Ureña, J.; Burgaya, F.; Pascual, M.; et al. Somatic signature of brain-specific single nucleotide variations in sporadic Alzheimer’s disease. J. Alzheimer’s Dis. JAD 2014, 42, 1357–1382. [Google Scholar] [CrossRef] [Green Version]
- Raghavan, N.S.; Dumitrescu, L.; Mormino, E.; Mahoney, E.R.; Lee, A.J.; Gao, Y.; Bilgel, M.; Goldstein, D.; Harrison, T.; Engelman, C.D.; et al. Association Between Common Variants in RBFOX1, an RNA-Binding Protein, and Brain Amyloidosis in Early and Preclinical Alzheimer Disease. JAMA Neurol. 2020, 77, 1288–1298. [Google Scholar] [CrossRef]
- Uhl, G.R.; Martinez, M.J. PTPRD: Neurobiology, genetics, and initial pharmacology of a pleiotropic contributor to brain phenotypes. Ann. N. Y. Acad. Sci. 2019, 1451, 112–129. [Google Scholar] [CrossRef]
- Liu, F.F.; Zhang, Z.; Chen, W.; Gu, H.Y.; Yan, Q.J. Regulatory mechanism of microRNA-377 on CDH13 expression in the cell model of Alzheimer’s disease. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 2801–2808. [Google Scholar] [CrossRef]
- Hsu, C.Y.; Lee, K.T.; Sun, T.Y.; Sze, C.I.; Huang, S.S.; Hsu, L.J.; Chang, N.S. WWOX and Its Binding Proteins in Neurodegeneration. Cells 2021, 10, 1781. [Google Scholar] [CrossRef]
- Huang, M.; Deng, C.; Yu, Y.; Lian, T.; Yang, W.; Feng, Q. Spatial correlations exploitation based on nonlocal voxel-wise GWAS for biomarker detection of AD. NeuroImage Clin. 2019, 21, 101642. [Google Scholar] [CrossRef] [PubMed]
- Ouellette, A.R.; Neuner, S.M.; Dumitrescu, L.; Anderson, L.C.; Gatti, D.M.; Mahoney, E.R.; Bubier, J.A.; Churchill, G.; Peters, L.; Huentelman, M.J.; et al. Cross-Species Analyses Identify Dlgap2 as a Regulator of Age-Related Cognitive Decline and Alzheimer’s Dementia. Cell Rep. 2020, 32, 108091. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Lee, T.; Choi, J.K.; Jeong, Y. Polymorphism in the MAGI2 Gene Modifies the Effect of Amyloid β on Neurodegeneration. Alzheimer Dis. Assoc. Disord. 2021, 35, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.P.; Soni, U. A review of dementia, focusing on the distinct roles of viral protein corona and MMP9 in dementia: Potential pharmacotherapeutic priorities. Ageing Res. Rev. 2022, 75, 101560. [Google Scholar] [CrossRef] [PubMed]
- Shang, Z.; Lv, H.; Zhang, M.; Duan, L.; Wang, S.; Li, J.; Liu, G.; Ruijie, Z.; Jiang, Y. Genome-wide haplotype association study identify TNFRSF1A, CASP7, LRP1B, CDH1 and TG genes associated with Alzheimer’s disease in Caribbean Hispanic individuals. Oncotarget 2015, 6, 42504–42514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreple, C.J.; Lu, Y.; Taugher, R.J.; Schwager-Gutman, A.L.; Du, J.; Stump, M.; Wang, Y.; Ghobbeh, A.; Fan, R.; Cosme, C.V.; et al. Acid-sensing ion channels contribute to synaptic transmission and inhibit cocaine-evoked plasticity. Nature neuroscience 2014, 17, 1083–1091. [Google Scholar] [CrossRef]
- Koran, M.E.; Hohman, T.J.; Meda, S.A.; Thornton-Wells, T.A. Genetic interactions within inositol-related pathways are associated with longitudinal changes in ventricle size. J. Alzheimer’s Dis. JAD 2014, 38, 145–154. [Google Scholar] [CrossRef]
- Ben-Avraham, D.; Karasik, D.; Verghese, J.; Lunetta, K.L.; Smith, J.A.; Eicher, J.D.; Vered, R.; Deelen, J.; Arnold, A.M.; Buchman, A.S.; et al. The complex genetics of gait speed: Genome-wide meta-analysis approach. Aging 2017, 9, 209–246. [Google Scholar] [CrossRef] [Green Version]
- James, A.W.; Shen, J.; Zhang, X.; Asatrian, G.; Goyal, R.; Kwak, J.H.; Jiang, L.; Bengs, B.; Culiat, C.T.; Turner, A.S.; et al. NELL-1 in the treatment of osteoporotic bone loss. Nat. Commun. 2015, 6, 7362. [Google Scholar] [CrossRef] [Green Version]
- Dong, C.; Beecham, A.; Wang, L.; Blanton, S.H.; Rundek, T.; Sacco, R.L. Follow-up association study of linkage regions reveals multiple candidate genes for carotid plaque in Dominicans. Atherosclerosis 2012, 223, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Miao, B.; Wang, S.; Dong, W.; Xu, H.; Si, C.; Wang, W.; Duan, S.; Lou, J.; Bao, Z.; et al. Hiplot: A comprehensive and easy-to-use web service boosting publication-ready biomedical data visualization. bioRxiv 2022. [Google Scholar] [CrossRef]
- Bi, X.a.; Hu, X.; Wu, H.; Wang, Y. Multimodal Data Analysis of Alzheimer’s Disease Based on Clustering Evolutionary Random Forest. IEEE J. Biomed. Health Inform. 2020, 24, 2973–2983. [Google Scholar] [CrossRef] [PubMed]
- Bi, X.a.; Zhou, W.; Li, L.; Xing, Z. Detecting Risk Gene and Pathogenic Brain Region in EMCI Using a Novel GERF Algorithm Based on Brain Imaging and Genetic Data. IEEE J. Biomed. Health Inform. 2021, 25, 3019–3028. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, W.; Cao, L.; Luo, H.; Xu, S.; Bao, P.; Meng, X.; Liang, H.; Fang, S. Hippocampal Subregion and Gene Detection in Alzheimer’s Disease Based on Genetic Clustering Random Forest. Genes 2021, 12, 683. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Li, Y.; Li, C.; Fan, L.; Liu, T.; Wang, J. Repeated anodal high-definition transcranial direct current stimulation over the left dorsolateral prefrontal cortex in mild cognitive impairment patients increased regional homogeneity in multiple brain regions. PLoS ONE 2021, 16, e0256100. [Google Scholar] [CrossRef]
- Heldmann, M.; Heeren, J.; Klein, C.; Rauch, L.; Hagenah, J.; Münte, T.F.; Kasten, M.; Brüggemann, N. Neuroimaging abnormalities in individuals exhibiting Parkinson’s disease risk markers. Mov. Disord. 2018, 33, 1412–1422. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.-W. Calcium-regulated signaling pathways. NeuroMolecular Med. 2004, 6, 53–64. [Google Scholar] [CrossRef]
- Kuchibhotla, K.V.; Goldman, S.T.; Lattarulo, C.R.; Wu, H.-Y.; Hyman, B.T.; Bacskai, B.J. Abeta plaques lead to aberrant regulation of calcium homeostasis in vivo resulting in structural and functional disruption of neuronal networks. Neuron 2008, 59, 214–225. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, I.L.; Bajouco, L.M.; Mota, S.I.; Auberson, Y.P.; Oliveira, C.R.; Rego, A.C. Amyloid beta peptide 1–42 disturbs intracellular calcium homeostasis through activation of GluN2B-containing N-methyl-d-aspartate receptors in cortical cultures. Cell Calcium. 2012, 51, 95–106. [Google Scholar] [CrossRef]
- Briggs, C.A.; Chakroborty, S.; Stutzmann, G.E. Emerging pathways driving early synaptic pathology in Alzheimer’s disease. Biochem. Biophys Res. Commun. 2017, 483, 988–997. [Google Scholar] [CrossRef] [Green Version]
- Bruno, A.M.; Huang, J.Y.; Bennett, D.A.; Marr, R.A.; Hastings, M.L.; Stutzmann, G.E. Altered ryanodine receptor expression in mild cognitive impairment and Alzheimer’s disease. Neurobiol Aging 2012, 33, 1001.e1–1001.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stutzmann, G.E.; Smith, I.; Caccamo, A.; Oddo, S.; Laferla, F.M.; Parker, I. Enhanced ryanodine receptor recruitment contributes to Ca2+ disruptions in young, adult, and aged Alzheimer’s disease mice. J. Neurosci. 2006, 26, 5180–5189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikkonen, M.; Soininen, H.; Tapiola, T.; Alafuzoff, I.; Miettinen, R. Hippocampal plasticity in Alzheimer’s disease: Changes in highly polysialylated NCAM immunoreactivity in the hippocampal formation. Eur. J. Neurosci. 1999, 11, 1754–1764. [Google Scholar] [CrossRef]
- Yew, D.T.; Li, W.P.; Webb, S.E.; Lai, H.W.L.; Zhang, L. Neurotransmitters, peptides, and neural cell adhesion molecules in the cortices of normal elderly humans and alzheimer patients: A comparison. Exp. Gerontol. 1999, 34, 117–133. [Google Scholar] [CrossRef]
- Aisa, B.; Gil-Bea, F.J.; Solas, M.; García-Alloza, M.; Chen, C.P.; Lai, M.K.; Francis, P.T.; Ramírez, M.J. Altered NCAM Expression Associated with the Cholinergic System in Alzheimer’s Disease. J. Alzheimer’s Dis. 2010, 20, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Jin, K.; Peel, A.L.; Mao, X.O.; Xie, L.; Cottrell, B.A.; Henshall, D.C.; Greenberg, D.A. Increased hippocampal neurogenesis in Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2004, 101, 343–347. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, H.; Kawamata, T.; Dedhar, S.; McGeer, P.L. Immunohistochemical localization of vitronectin, its receptor and beta-3 integrin in Alzheimer brain tissue. J. Neuroimmunol. 1991, 32, 19–28. [Google Scholar] [CrossRef]
- Van Gool, D.; Carmeliet, G.; Triau, E.; Cassiman, J.-J.; Dom, R. Appearance of localized immunoreactivity for the α4 integrin subunit and for fibronectin in brains from Alzheimer’s, Lewy body dementia patients and aged controls. Neurosci. Lett. 1994, 170, 71–73. [Google Scholar] [CrossRef]
- Koenigsknecht, J.; Landreth, G. Microglial phagocytosis of fibrillar beta-amyloid through a beta1 integrin-dependent mechanism. J. Neurosci. 2004, 24, 9838–9846. [Google Scholar] [CrossRef] [Green Version]
- Ashby, E.L.; Kehoe, P.G. Current status of renin–aldosterone angiotensin system-targeting anti-hypertensive drugs as therapeutic options for Alzheimer’s disease. Expert Opin. Investig. Drugs 2013, 22, 1229–1242. [Google Scholar] [CrossRef]
- Eren, N.; Deni, Z.; Yildiz, Z.; Gö, N.; Gü, L.; Karabiyik, T. P200-levels of Apelin-13 and total oxidant/antioxidant status in sera of Alzheimer patients. Turk. J. Biochem./Turk Biyokim. Derg. 2012, 37, 341. [Google Scholar]
- Dai, T.-T.; Wang, B.; Xiao, Z.-Y.; You, Y.; Tian, S.-W. Apelin-13 Upregulates BDNF Against Chronic Stress-induced Depression-like Phenotypes by Ameliorating HPA Axis and Hippocampal Glucocorticoid Receptor Dysfunctions. Neuroscience 2018, 390, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Haghparast, E.; Esmaeili-Mahani, S.; Abbasnejad, M.; Sheibani, V. Apelin-13 ameliorates cognitive impairments in 6-hydroxydopamine-induced substantia nigra lesion in rats. Neuropeptides 2018, 68, 28–35. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subjects | HC | EMCI | AD | p |
|---|---|---|---|---|
| Number | 353 | 273 | 296 | - |
| Gender (M/F) | 187/166 | 153/120 | 166/130 | <0.001 |
| Age (mean ± sd) | 72.2 ± 7.6 | 71.3 ± 7.1 | 75.1 ± 5.5 | <0.001 |
| Edu (mean ± sd) | 16.1 ± 2.7 | 16.1 ± 2.6 | 16.3 ± 2.6 | <0.001 |
| AD-HC | AD-EMCI | EMCI-HC | |||
|---|---|---|---|---|---|
| Brain Region | Number of Features | Brain Region | Number of Features | Brain Region | Number of Features |
| Frontal_Sup_R | 9 | Temporal_Inf_R | 7 | Temporal_Sup_R | 6 |
| Frontal_Mid_L | 5 | Precuneus_R | 6 | Frontal_Sup_L | 5 |
| Lingual_R | 5 | Frontal_Mid_L | 5 | Frontal_Inf_Orb_L | 5 |
| SupraMarginal_R | 5 | Precuneus_L | 5 | Frontal_Sup_Medial_L | 5 |
| Temporal_Mid_L | 5 | Postcentral_L | 4 | Calcarine_R | 5 |
| Frontal_Sup_L | 4 | Temporal_Sup_R | 4 | Fusiform_L | 5 |
| Frontal_Mid_R | 4 | Frontal_Mid_R | 3 | SupraMarginal_L | 5 |
| Lingual_L | 4 | Calcarine_L | 3 | Precuneus_R | 5 |
| Fusiform_L | 4 | Occipital_Mid_L | 3 | Temporal_Mid_L | 5 |
| Postcentral_R | 4 | Occipital_Mid_R | 3 | Temporal_Inf_R | 5 |
| Genes | AD-HC | AD-EMCI | EMCI-HC | References |
|---|---|---|---|---|
| p-Value | p-Value | p-Value | ||
| CSMD1 | 2.998108 × 10−36 | 1.02583 × 10−29 | 1.61113 × 10−35 | Parcerisas et al. [26] |
| RBFOX1 | 5.84303 × 10−22 | 1.37062 × 10−20 | 6.3792 × 10−26 | Raghavan et al. [27] |
| PTPRD | 3.43579 × 10−21 | 3.81205 × 10−24 | 1.52404 × 10−26 | Uhl et al. [28] |
| CDH13 | 5.58042 × 10−20 | 1.85248 × 10−14 | 6.10705 × 10−13 | Liu et al. [29] |
| WWOX | 7.1123 × 10−17 | 2.9447 × 10−20 | 2.46024 × 10−22 | Hsu et al. [30] |
| Group | Gene | p-Value | References |
|---|---|---|---|
| AD-HC | MIR8063 | 1.827314 × 10−221 | - |
| MEIS2 | 3.0419 × 10−147 | Huang et al. [31] | |
| DLGAP2 | 3.57803 × 10−19 | Ouellette et al. [32] | |
| MAGI2 | 8.108022 × 10−15 | Kim et al. [33] | |
| AD-EMCI | PRKN | 2.52709 × 10−15 | Panda et al. [34] |
| LRP1B | 1.50983 × 10−13 | Shang et al. [35] | |
| ASIC2 | 4.2832 × 10−13 | Kreple et al. [36] | |
| PRKG1 | 5.64312 × 10−13 | Koran et al. [37] | |
| EMCI-HC | PTPRT | 1.11037 × 10−14 | Ben et al. [38] |
| NELL1 | 1.14303 × 10−12 | James et al. [39] | |
| AGBL1 | 2.88479 × 10−11 | Dong et al. [40] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, X.; Wei, Q.; Meng, L.; Liu, J.; Wu, Y.; Liu, W. Feature Fusion and Detection in Alzheimer’s Disease Using a Novel Genetic Multi-Kernel SVM Based on MRI Imaging and Gene Data. Genes 2022, 13, 837. https://doi.org/10.3390/genes13050837
Meng X, Wei Q, Meng L, Liu J, Wu Y, Liu W. Feature Fusion and Detection in Alzheimer’s Disease Using a Novel Genetic Multi-Kernel SVM Based on MRI Imaging and Gene Data. Genes. 2022; 13(5):837. https://doi.org/10.3390/genes13050837
Chicago/Turabian StyleMeng, Xianglian, Qingpeng Wei, Li Meng, Junlong Liu, Yue Wu, and Wenjie Liu. 2022. "Feature Fusion and Detection in Alzheimer’s Disease Using a Novel Genetic Multi-Kernel SVM Based on MRI Imaging and Gene Data" Genes 13, no. 5: 837. https://doi.org/10.3390/genes13050837
APA StyleMeng, X., Wei, Q., Meng, L., Liu, J., Wu, Y., & Liu, W. (2022). Feature Fusion and Detection in Alzheimer’s Disease Using a Novel Genetic Multi-Kernel SVM Based on MRI Imaging and Gene Data. Genes, 13(5), 837. https://doi.org/10.3390/genes13050837