
The Pid Family Has Been Diverged into Xian and Geng Type Resistance Genes against Rice Blast Disease



Abstract
:1. Introduction
2. Materials and Methods
2.1. Development of a Comprehensive FNP Marker System
2.2. Marker Verification
2.3. Genotyping and Data Analysis
2.4. Validation of Candidate Functional Allele
3. Results
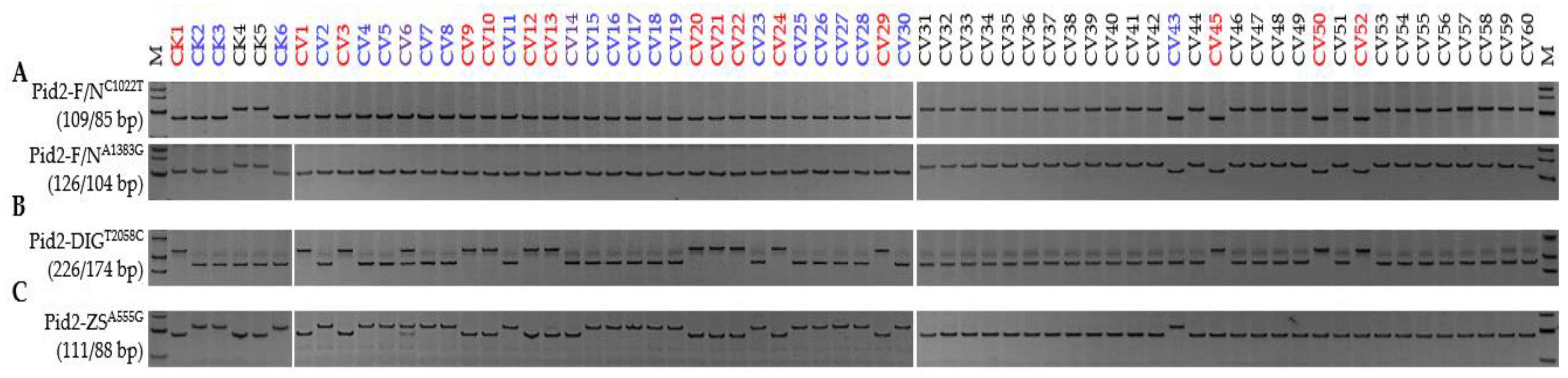
3.1. Pid2 Alleles
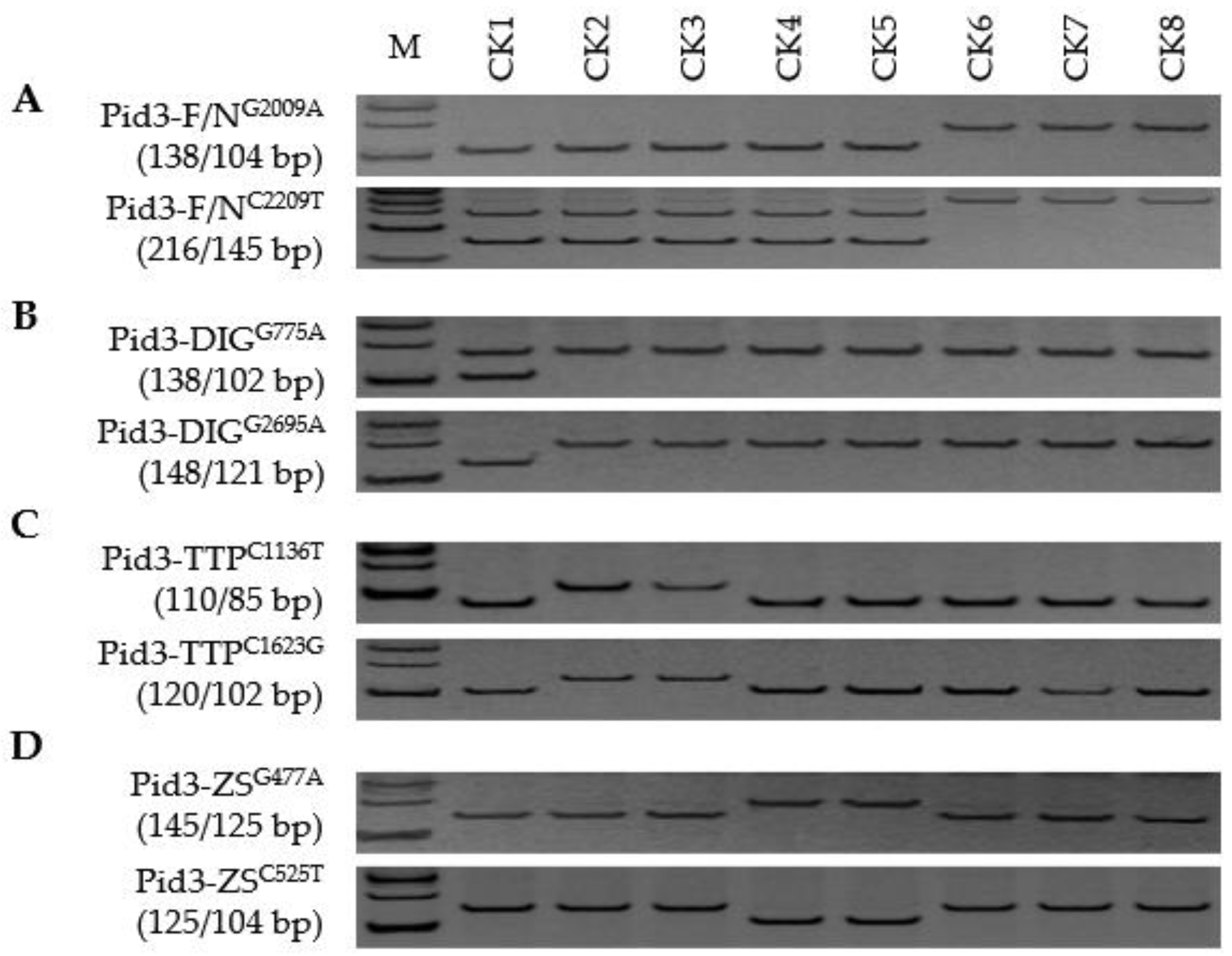
3.2. Pid3 Alleles
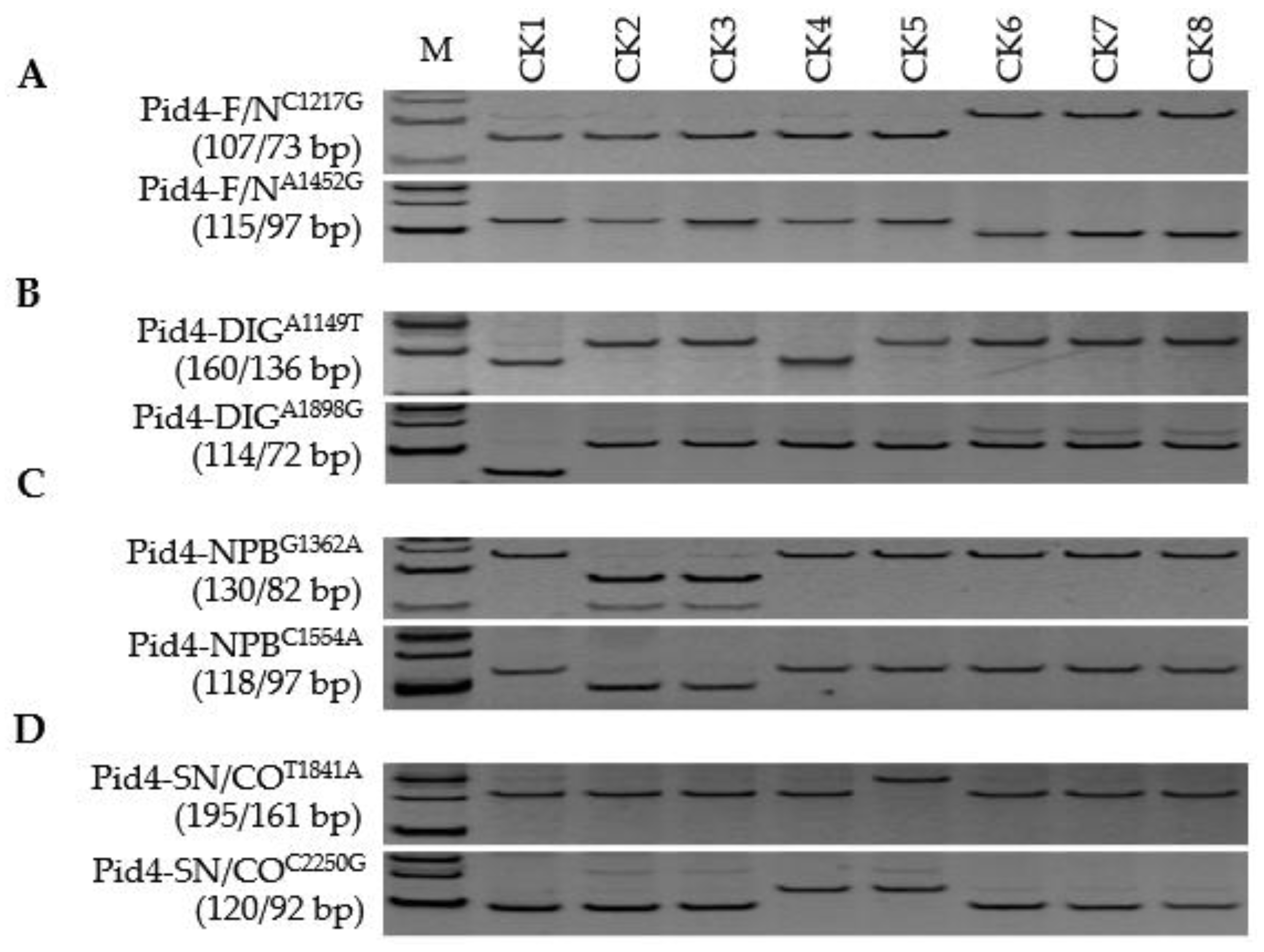
3.3. Pid4 Alleles
3.4. Performance of Candidate Functional Allele
4. Discussion
4.1. The Comprehensive FNP Marker Systems Have Largely Improved the Marker Works
4.2. The Pid Family Has Strictly Diverged into Xian and Geng Subspecies
4.3. Rather Limited Genotypes of the Pid Family Are Effective in Both the Xian and Geng Rice Groups
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, X.; Zhao, Y.; Wei, X.; Li, C.; Wang, A.; Zhao, Q.; Li, W.; Guo, Y.; Deng, L.; Zhu, C.; et al. Genome-wide association study of flowering time and grain yield traits in a worldwide collection of rice germplasm. Nat. Genet. 2012, 44, 32–39. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, L.; Xing, F.; Kudrna, D.; Yao, D.; Copetti, W.; Mu, T.; Li, W.; Song, J.; Xie, W.; et al. Zhang, Extensive sequence divergence between the reference genomes of two elite Xian rice varieties Zhenshan 97 and Minghui 63. Proc. Natl. Acad. Sci. USA 2016, 113, E5163–E5171. [Google Scholar]
- Du, H.; Yu, Y.; Ma, Y.; Gao, Q.; Cao, Y.; Zhen, Z.; Ma, B.; Qi, M.; Li, Y.; Zhao, X.; et al. Sequencing and de novo assembly of a near complete Xian rice genome. Nat. Commun. 2017, 8, 15324. [Google Scholar] [CrossRef]
- Wang, W.; Mauleon, R.; Hu, Z.; Chebotarov, D.; Tai, S.; Wu, Z.; Li, M.; Zheng, T.; Fuentes, R.; Zhang, F.; et al. Genomic variation in 3010 diverse accessions of Asian cultivated rice. Nature 2018, 557, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Li, W.; Sun, Z.; Ouyang, N.; Jing, X.; He, Q.; Wu, J.; Zheng, J.; Zheng, J.; Tang, S.; et al. Resequencing of 1,143 Xian rice accessions reveals important genetic variations and different heterosis patterns. Nat. Commun. 2020, 11, 4778. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Purugganan, M. Multiple origin but single domestication led to Oryza sativa. G3 Genes Genomes Genet. 2018, 8, 797–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, Y.; Zhang, Q. Understanding reproductive isolation based on the rice model. Annu. Rev. Plant Biol. 2013, 64, 111–135. [Google Scholar] [CrossRef] [Green Version]
- Dean, R.; Van Kan, J.; Pretorius, Z.; Hammond-Kosack, K.; Pietro, A.; Spanu, P.; Rudd, J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plan. Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Wang, G. Plant innate immunity in rice: A defense against pathogen infection. Natl. Sci. Rev. 2016, 3, 295–308. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhu, Q.; Yao, Y.; Zhao, Z.; Correll, J.; Wang, L.; Pan, Q. The race structure of the rice blast pathogen across southern and northeastern China. Rice 2017, 10, 46. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, J.; Yao, Y.; Jin, X.; Correll, J.; Wang, L.; Pan, Q. Dynamics of race structures of the rice blast pathogen populations in Heilongjiang province, China from 2006 through 2015. Plant Dis. 2019, 103, 2759–2763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.; Wang, X.; Jia, Y.; Minkenberg, B.; Wheatley, M.; Fan, J.; Jia, M.; Famoso, A.; Edwards, J.; Wamishe, Y.; et al. The rice blast resistance gene Ptr encodes an atypical protein required for broad-spectrum disease resistance. Nat. Commun. 2018, 9, 2039. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Wang, J.; Zhang, Y.; Yao, Y.; Huang, L.; Yang, X.; Wang, L.; Pan, Q. Dynamics of race structures of Pyricularia oryzae populations across 18 seasons in Guangdong province, China. Plant Dis. 2021, 105, 144–148. [Google Scholar] [CrossRef]
- He, X.; Liu, X.; Wang, L.; Lin, F.; Cheng, Y.; Chen, Z.; Liao, Y.; Pan, Q. Identification of the novel recessive gene pi55(t) conferring resistance to Magnaporthe oryzae. Sci. China Life Sci. 2012, 55, 141–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, C.; Zhang, Y.; Yao, N.; Lin, F.; Liu, Z.; Dong, Z.; Wang, L.; Pan, Q. Function and interaction of the coupled genes responsible for Pik-h encoded rice blast resistance. PLoS ONE 2014, 9, e98067. [Google Scholar] [CrossRef]
- Deng, Y.; Zhai, K.; Xie, Z.; Yang, D.; Zhu, X.; Liu, J.; Wang, X.; Qin, P.; Yang, Y.; Zhang, G.; et al. Epigenetic regulation of antagonistic receptors confers rice blast resistance with yield balance. Science 2017, 355, 962–965. [Google Scholar] [CrossRef]
- Flor, H. Current status of the gene for gene concept. Ann. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Ebbole, D. Magnaporthe as a model for understanding host-pathogen interactions. Annu. Rev. Phytopathol. 2007, 45, 437–456. [Google Scholar] [CrossRef]
- Wu, W.; Wang, L.; Zhang, S.; Li, Z.; Zhang, Y.; Lin, F.; Pan, Q. Stepwise arms race between AvrPik and Pik alleles in the rice blast pathosystem. Mol. Plant-Microbe Interac. 2014, 27, 759–769. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Wang, L.; Wu, W.; He, L.; Yang, X.; Pan, Q. Function and evolution of Magnaporthe oryzae avirulence gene AvrPib responding to the rice blast resistance gene Pib. Sci. Rep. 2015, 5, 11642. [Google Scholar] [CrossRef] [Green Version]
- Zeigler, R.; Cuoc, L.; Scott, R.; Bernardo, M.; Chen, D.; Valent, B.; Nelson, R. The relationship between lineage and virulence in Pyricularia grisea in the Philippines. Phytopathology 1995, 85, 443–451. [Google Scholar] [CrossRef]
- Wang, X.; Jia, Y.; Wamishe, Y.; Jia, M.; Valent, B. Dynamic changes in the rice blast population in the USA over six decades. Mol. Plant-Microbe Interac. 2017, 30, 803–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryan, G.; Wu, K.; Farrall, L.; Jia, Y.; Hershey, H.; McAdams, S.; Faulk, K.; Donaldson, G.; Tarchini, R.; Valent, B. A single amino acid difference distinguishes resistant and susceptible alleles of the rice blast resistance gene Pita. Plant Cell 2000, 12, 2033–2045. [Google Scholar] [PubMed] [Green Version]
- Lin, F.; Chen, S.; Que, Z.; Wang, L.; Liu, X.; Pan, Q. The blast resistance gene Pi37 encodes an NBS-LRR protein and is a member of a resistance gene cluster on rice chromosome 1. Genetics 2007, 177, 1871–1880. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Lin, F.; Wang, L.; Pan, Q. The in silico map-based cloning of Pi36, a gene encodes a coiled-coil NBS-LRR protein, conferring race-specific resistance to Magnaporthe oryzae. Genetics 2007, 176, 2541–2549. [Google Scholar] [CrossRef] [Green Version]
- Fukuoka, S.; Saka, N.; Koga, H.; Ono, K.; Shimizu, T.; Ebana, K.; Hayashi, N.; Takahashi, A.; Hirochika, H.; Okuno, K.; et al. Loss of function of a proline-containing protein confers durable disease resistance in rice. Science 2009, 325, 998–1001. [Google Scholar] [CrossRef]
- Hua, L.; Wu, J.; Chen, C.; Wu, W.; He, X.; Lin, F.; Wang, L.; Ashikawa, I.; Matsumoto, T.; Wang, L.; et al. The isolation of Pi1, an allele at Pik locus which confers broad spectrum resistance to rice blast. Theor. Appl. Genet. 2012, 125, 1047–1055. [Google Scholar] [CrossRef]
- Zhai, C.; Wang, L.; Dong, Z.; Zeng, X.; Lin, F.; Pan, Q. The rice blast resistance gene, Pik, is the third allele at the Pik cluster that is distinguished by a function-associated SNP. New Phytol. 2011, 189, 321–334. [Google Scholar] [CrossRef]
- Li, C.; Wang, D.; Peng, S.; Chen, Y.; Su, P.; Chen, J.; Zheng, L.; Tan, X.; Liu, J.; Xiao, Y.; et al. Genome-wide association mapping of resistance against rice blast strains in South China and identification of a new Pik allele. Rice 2019, 12, 47. [Google Scholar] [CrossRef]
- Teerasan, W.; Srikaew, I.; Phaitreejit, K.; Kate-Ngam, S.; Jantasuriyarat, C. Gene-specific marker screening and disease reaction validation of blast resistance genes, Pid3, Pigm and Pi54 in Thai landrace rice gerplasm and recommended rice varieties. Plant Genet. Resour. Charact. Util. 2019, 17, 421–426. [Google Scholar] [CrossRef]
- Tian, D.; Lin, Y.; Chen, Z.; Chen, Z.; Yang, F.; Wang, F.; Wang, Z.; Wang, M. Exploring the distribution of blast resistance alleles at the Pi2/9 locus in major rice-producing areas of China by a novel InDel marker. Plant Dis. 2020, 104, 1932–1938. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Yang, J.; Zhu, X.; Wu, J.; Zhou, B. Prevalence of ineffective haplotypes at the rice blast resistance (R) gene loci in Chinese elite hybrid varieties revealed by sequence-based molecular diagnosis. Rice 2020, 13, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Lei, F.; Wang, Q.; He, W.; Yuan, B.; Yuan, W. Identification of novel alleles of the rice blast-resistance gene Pi9 through sequence-based allele mining. Rice 2020, 13, 80. [Google Scholar] [CrossRef]
- Chen, X.; Shang, J.; Chen, D.; Lei, C.; Zou, Y.; Zhai, W.; Liu, G.; Xu, J.; Ling, Z.; Cao, G.; et al. A B-lectin receptor kinase gene conferring rice blast resistance. Plant J. 2006, 46, 794–804. [Google Scholar] [CrossRef]
- Chen, Z.; Zhao, W.; Zhu, X.; Zou, C.; Yin, J.; Chen, M.; Zhou, X.; Ying, H.; Jiang, X.; Li, Y.; et al. Identification and characterization of rice blast resistance gene Pid4 by a combination of transcriptomic profiling and genome analysis. J. Genet. Genom. 2018, 45, 665–672. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Tao, Y.; Chen, X.; Zou, Y.; Lei, C.; Wang, J.; Li, X.; Zhao, X.; Zhang, M.; Li, Z.; et al. Identification of a new rice blast resistance gene, Pid3, by genome-wide comparison of paired nucleotide-binding site-leucine-rich repeat genes and their pseudogene alleles between the two sequenced rice genomes. Genetics 2009, 182, 1303–1311. [Google Scholar] [CrossRef] [Green Version]
- Zeng, S. Identification of Rice Blast Resistance Genes in the Key Parental Cultivars. Master’s Thesis, South China Agri University, Guangzhou, China, June 2020. (In Chinese with English Abstract). [Google Scholar]
- Zhang, R. Resistance Gene Composition of Differential Cultivars for rice Blast Pathosystem. Master’s Thesis, South China Agri University, Guangzhou, China, June 2020. (In Chinese with English Abstract). [Google Scholar]
- Fernando, D.; Zhang, X.; Selin, C.; Zou, Z.; Liban, S.; McLaren, D.; Kubinec, A.; Parks, P.; Rashid, H.; Padmathilake, R.; et al. A six-year investigation of the dynamics of avirulence allele profiles, blackleg incidence, and mating type alleles of Leptosphaeria maculans populations associated with canola crops in Manitoba, Canada. Plant Dis. 2018, 102, 790–798. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Yu, S.; Zeng, D.; Liu, H.; Wang, H.; Yang, Z.; Xie, X.; Shen, R.; Tan, J.; Li, H.; et al. Development of “Purple Endosperm Rice” by engineering anthocyanin biosynthesis in the endosperm with a high-efficiency transgene stacking system. Mol. Plant 2017, 10, 918–929. [Google Scholar] [CrossRef] [Green Version]
- Hiei, Y.; Ohta, S.; Komari, T.; Kumashiro, T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J. 1994, 6, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Pan, Q.; Wang, L.; Ikehashi, H.; Tanisaka, T. Identification of a new blast resistance gene in the Xian rice cultivar Kasalath using Japanese differential cultivars and isozyme markers. Phytopathology 1996, 86, 1071–1075. [Google Scholar] [CrossRef]
- Yuan, B.; Wang, L.; Zeng, X.; Xu, X.; Zhai, C.; Hu, H.; Lin, F.; Pan, Q. The Pik-p resistance to Magnaporthe oryzae in rice is mediated by a pair of closely linked CC-NBS-LRR genes. Theor. Appl. Genet. 2011, 122, 1017–1028. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Huang, Z.; Xu, X.; Tang, L.; Liu, H.; Wang, C.; Zhou, Z.; Xin, Y.; Xing, J.; Peng, Z.; et al. Allelic variation of the rice blast resistance gene Pid3 in cultivated rice worldwide. Sci. Rep. 2017, 7, 10362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Promchuay, A.; Nilthong, S. Investigation of Pid3 rice blast resistance gene in northern upland rice varieties (Oryza sativa L.), Thailand using molecular markers. J. Advanced Agric. Technol. 2017, 4, 209–214. [Google Scholar]
- Chen, J.; Ding, J.; Ouyang, Y.; Du, H.; Yang, J.; Cheng, K.; Zhao, J.; Qiu, S.; Zhang, X.; Yao, E.J.; et al. A triallelic system of S5 is a major regulator of the reproductive barrier and compatibility of Xian-Geng hybrids in rice. Proc. Natl. Acad. Sci. USA 2008, 195, 11436–11441. [Google Scholar] [CrossRef] [Green Version]
- Yano, M.; Katayose, Y.; Ashikari, M.; Yamanouchi, U.; Monna, L.; Fuse, T.; Baba, T.; Yamamoto, K.; Umehara, Y.; Nagamura, Y.; et al. Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANS. Plant Cell 2000, 12, 2473–2483. [Google Scholar] [CrossRef] [Green Version]
- Izawa, T.; Oikawa, T.; Tokutomi, S.; Okuno, K.; Shimamoto, K. Phytochromes confer the photoperiodic control of flowering in rice (a short-day plant). Plant J. 2000, 22, 391–399. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, S.; Cao, Z.; Ouyang, Y.; Zhang, Q.; Xie, L.; Zheng, R.; Guo, M.; Ma, M.; Hu, Z.; et al. Chromatin loops associated with active genes and heterochromatin shape rice genome architecture for transcriptional regulation. Nat. Commun. 2019, 10, 3640. [Google Scholar] [CrossRef] [Green Version]
- Liu, D. Decoding Pedigrees of the Top-10 General Cultivars and Rice F1 hybrid Systems in China. Available online: https://mp.weixin.qq.com/s/7L5PcYMxGFXLJlVjfSM3dQ (accessed on 23 February 2021).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alleles vs. Genotypes | Xian Group (a, Presence/b, Absence) | Geng Group (c, Presence/d, Absence) | χ2 for Homogeneity a |
|---|---|---|---|
| Alleles | |||
| Pid2 alleles | 69/1 | 4/54 | 105.07 *** |
| Pid2-ZS | 49 | 1 | |
| Pid2-DIG | 17 | 3 | |
| Pid2-new | 3 | 0 | |
| Pid2-null | 1 | 54 | |
| Pid3 alleles | 69/1 | 4/54 | 105.07 *** |
| Pid3-ZS | 60 | 3 | |
| Pid3-DIG | 5 | 1 | |
| Pid3-TTP | 2 | 0 | |
| Pid3-new | 2 | 0 | |
| Pid3-null | 1 | 54 | |
| Pid4 alleles | 19/51 | 56/2 | 60.16 *** |
| Pid4-NPB | 1 | 21 | |
| Pid4-SN | 3 | 16 | |
| Pid4-CO | 4 | 14 | |
| Pid4-new | 9 | 5 | |
| Pid4-DIG | 2 | 0 | |
| Pid4-null | 51 | 2 | |
| Genotypes | |||
| d4 | 1/69 | 54/4 | 105.07 *** |
| d2-d3 | 51/19 | 2/56 | 60.16 *** |
| d2-d3-d4 | 18/52 | 2/56 | 10.30 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chai, R.; Wang, J.; Wang, X.; Wen, J.; Liang, Z.; Ye, X.; Zhang, Y.; Yao, Y.; Zhang, J.; Zhang, Y.; et al. The Pid Family Has Been Diverged into Xian and Geng Type Resistance Genes against Rice Blast Disease. Genes 2022, 13, 891. https://doi.org/10.3390/genes13050891
Chai R, Wang J, Wang X, Wen J, Liang Z, Ye X, Zhang Y, Yao Y, Zhang J, Zhang Y, et al. The Pid Family Has Been Diverged into Xian and Geng Type Resistance Genes against Rice Blast Disease. Genes. 2022; 13(5):891. https://doi.org/10.3390/genes13050891
Chicago/Turabian StyleChai, Ruipeng, Jinyan Wang, Xing Wang, Jianqiang Wen, Zhijian Liang, Xuemei Ye, Yaling Zhang, Yongxiang Yao, Jianfu Zhang, Yihua Zhang, and et al. 2022. "The Pid Family Has Been Diverged into Xian and Geng Type Resistance Genes against Rice Blast Disease" Genes 13, no. 5: 891. https://doi.org/10.3390/genes13050891
APA StyleChai, R., Wang, J., Wang, X., Wen, J., Liang, Z., Ye, X., Zhang, Y., Yao, Y., Zhang, J., Zhang, Y., Wang, L., & Pan, Q. (2022). The Pid Family Has Been Diverged into Xian and Geng Type Resistance Genes against Rice Blast Disease. Genes, 13(5), 891. https://doi.org/10.3390/genes13050891