
Fish as Model Systems to Study Epigenetic Drivers in Human Self-Domestication and Neurodevelopmental Cognitive Disorders

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Collection
2.1.1. European Sea Bass Early Domesticates
2.1.2. Anatomically Modern Humans (AMH)
2.1.3. Neurodevelopmental Cognitive Disorders
2.2. Comparative Analyses
3. Results
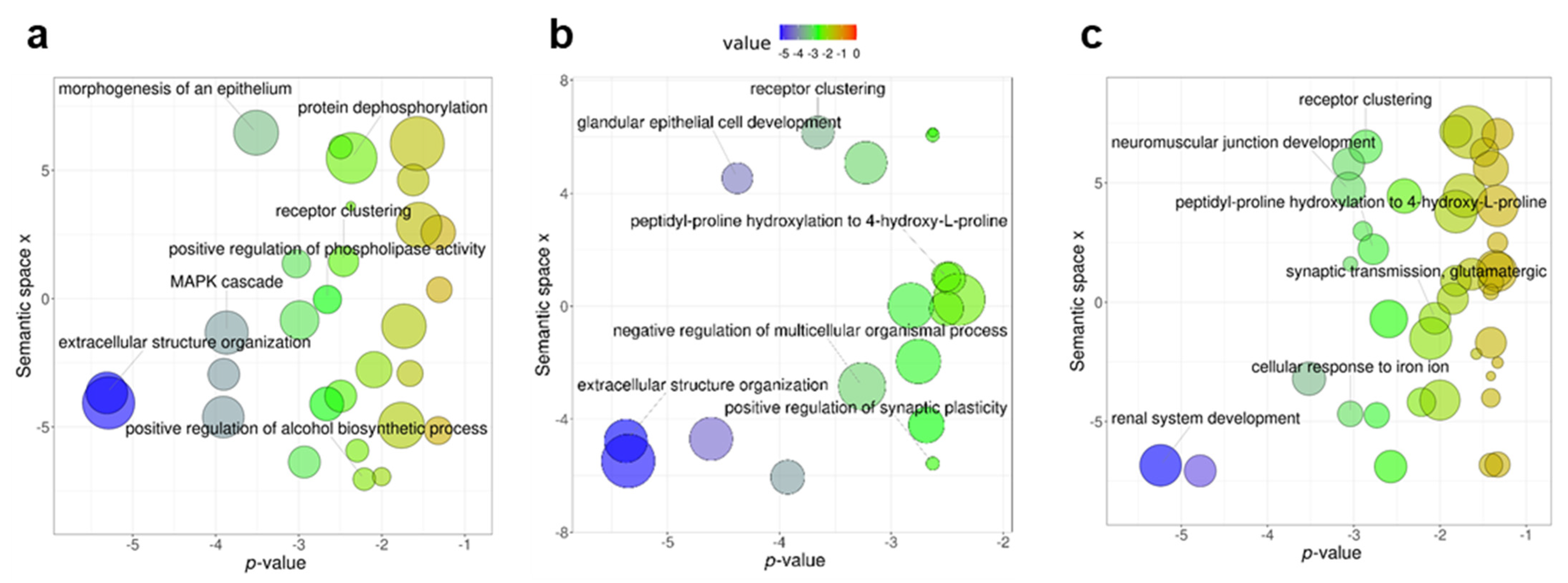
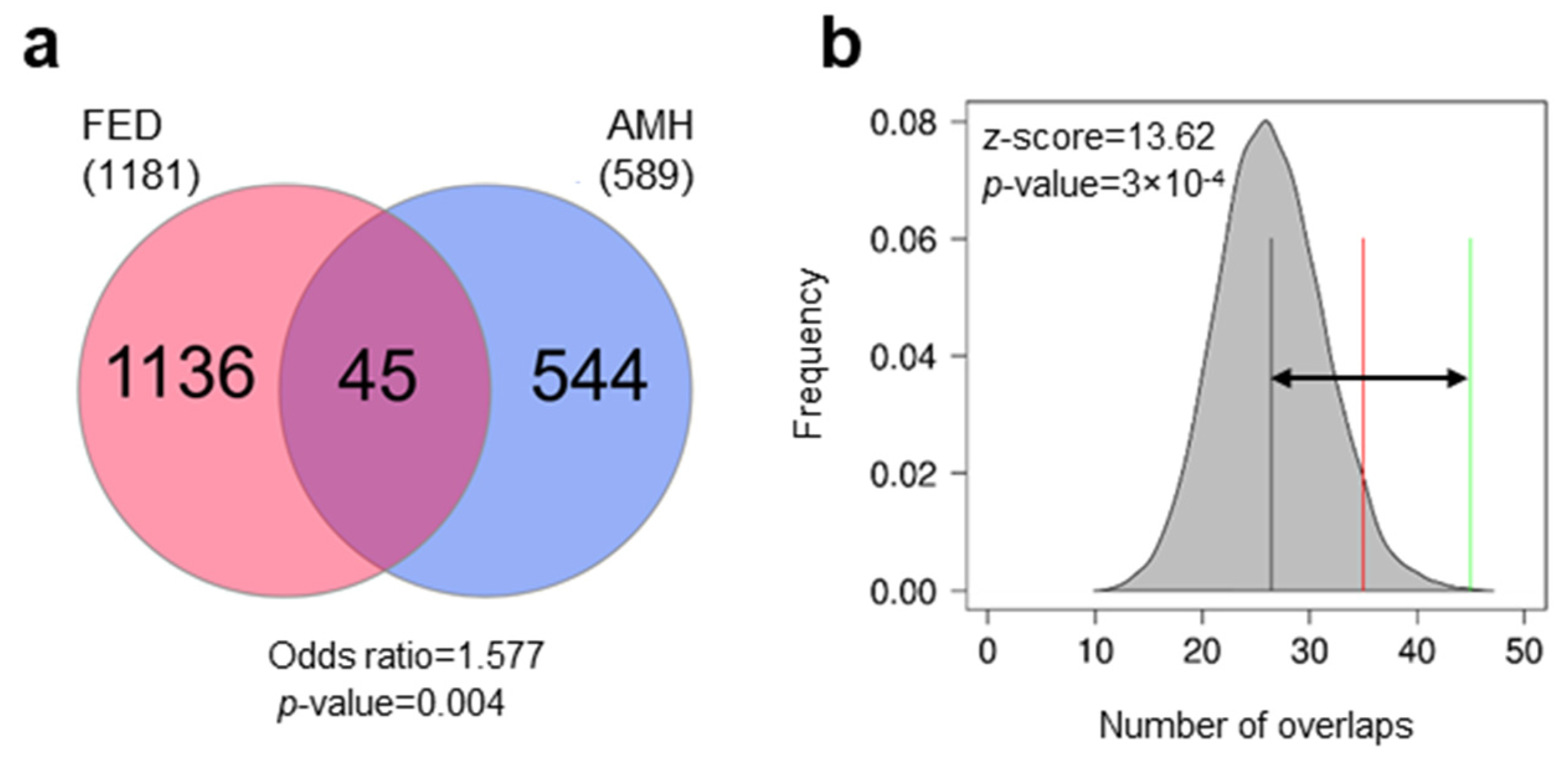
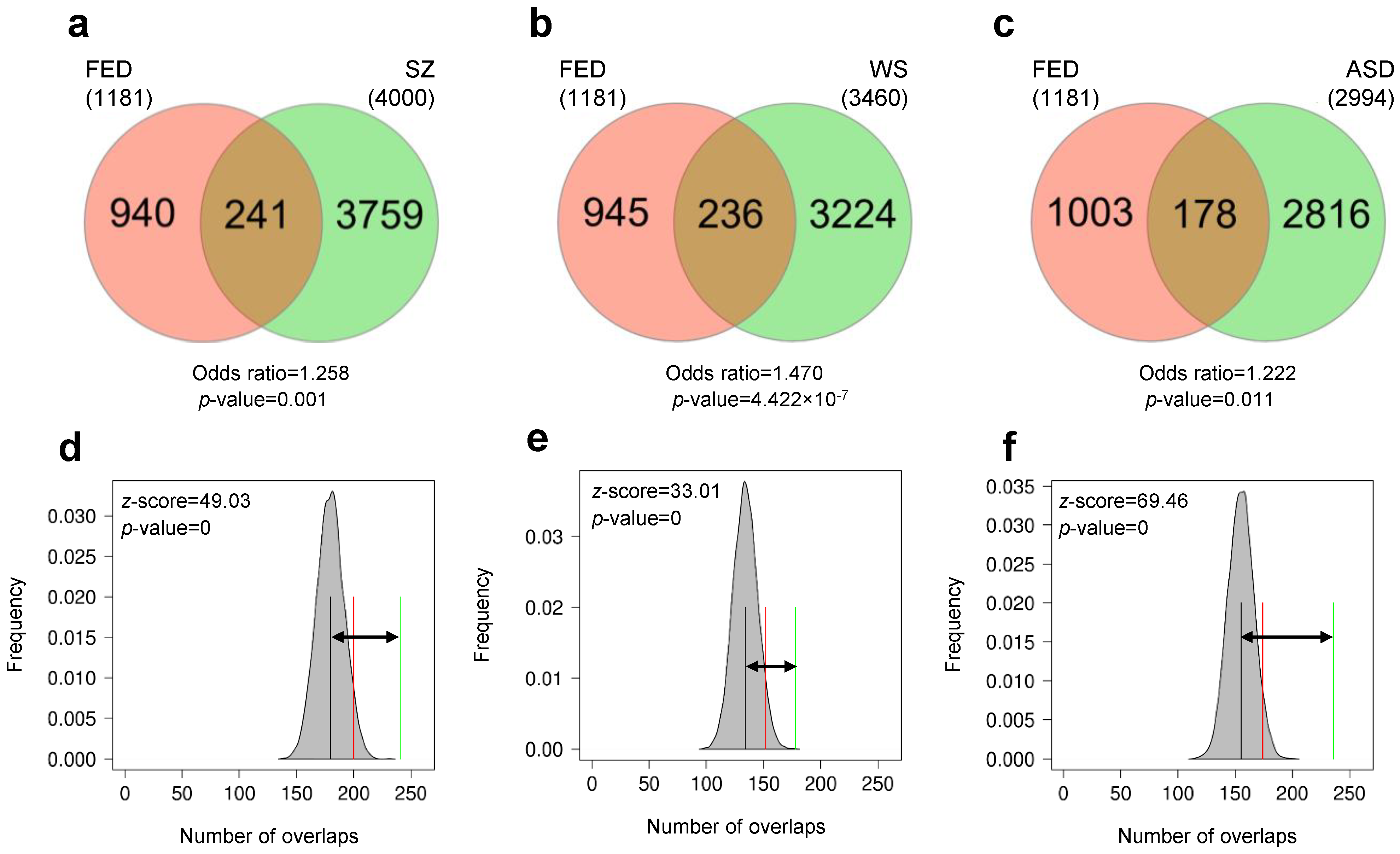
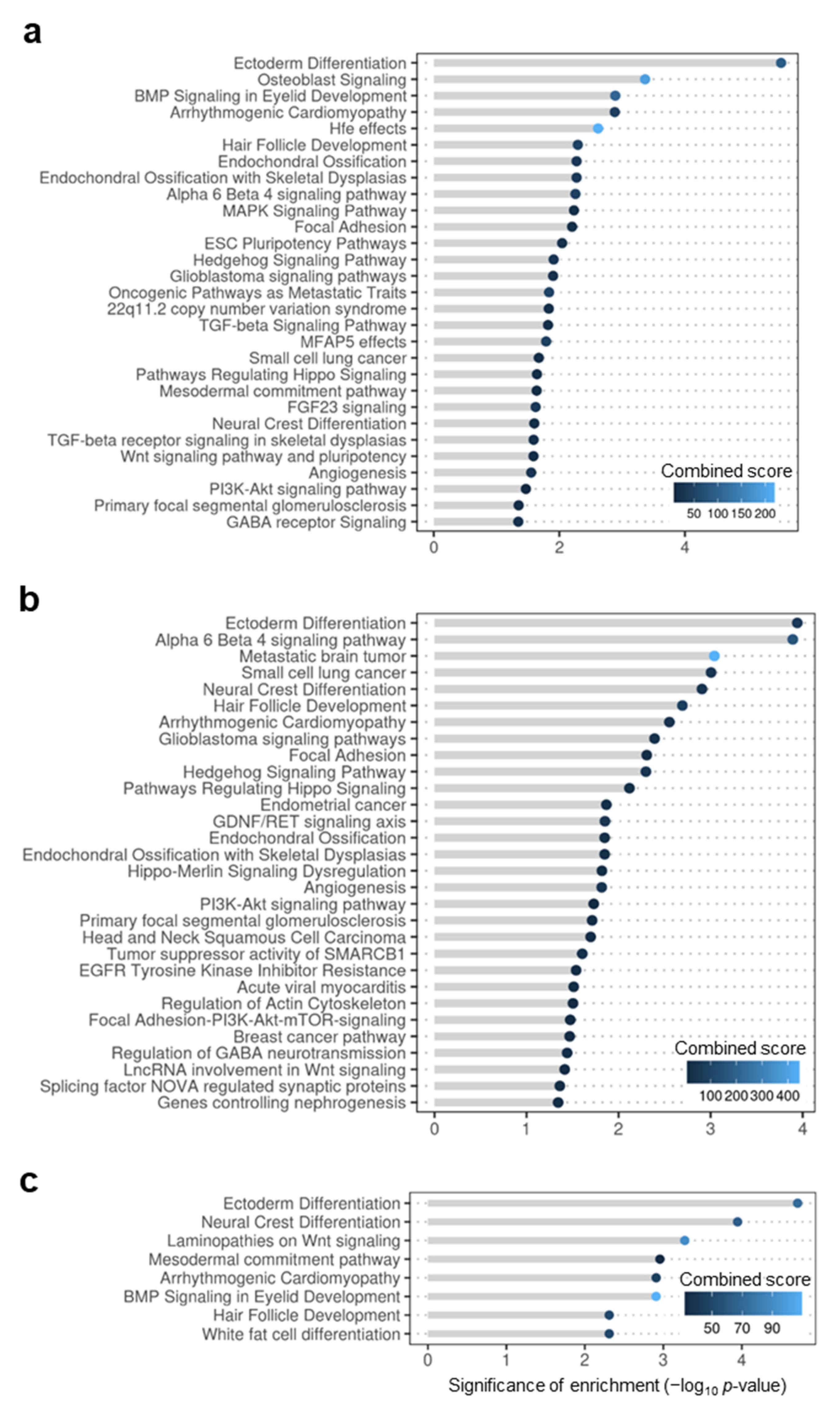
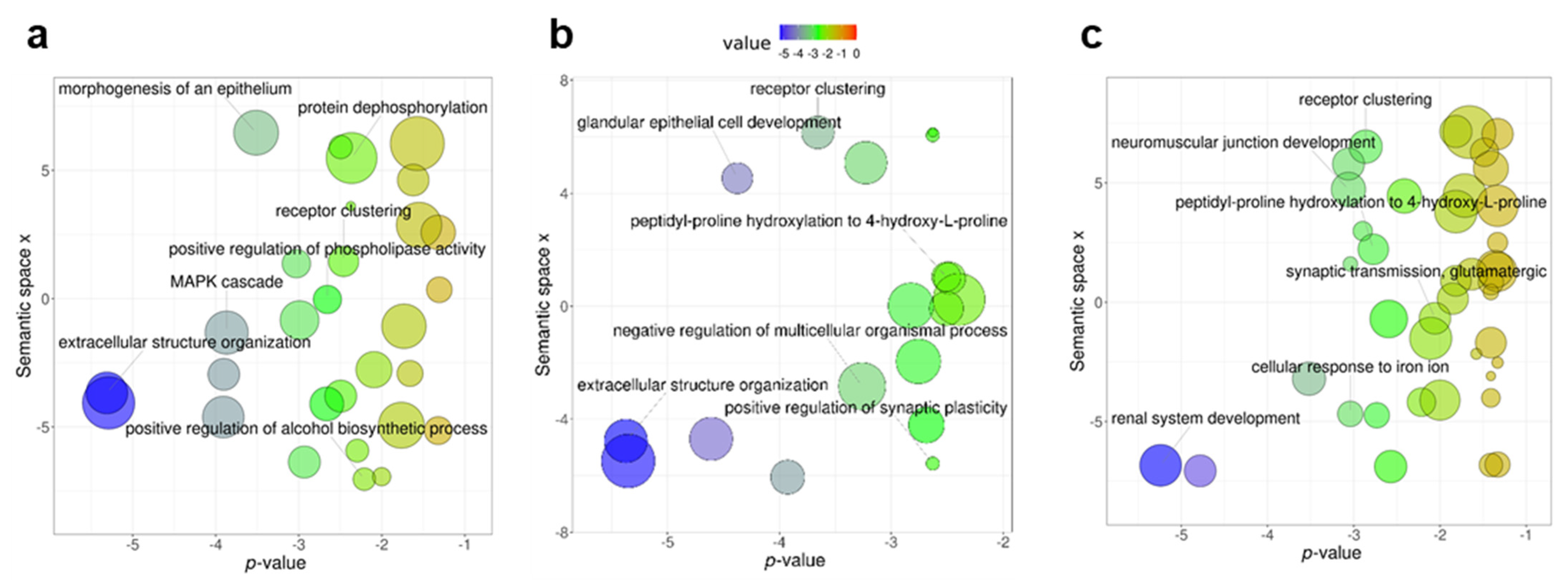
3.1. Differentially Methylated Genes during Early Domestication in European Sea Bass and in Humans Are Shared
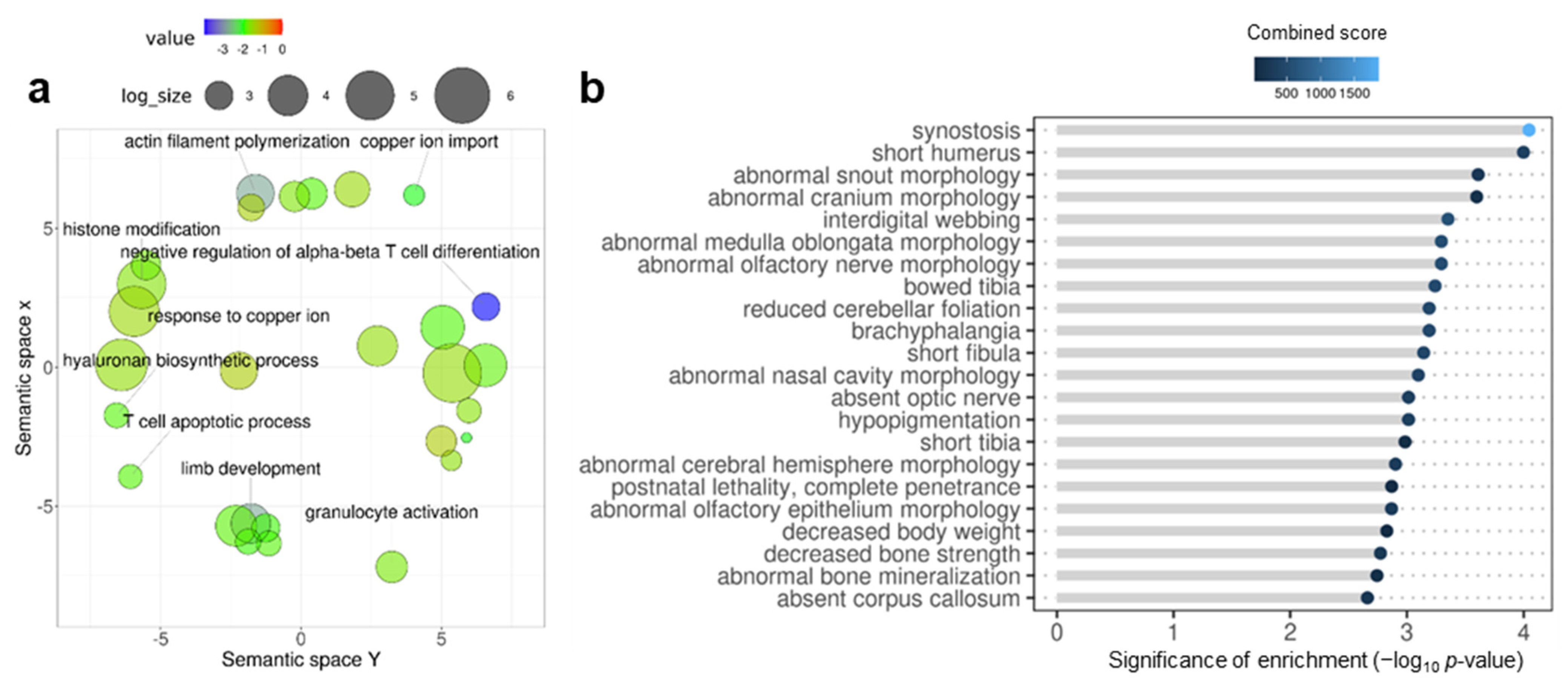
3.2. Early Domestication in European Sea Bass and Neurodevelopmental Cognitive Disorders Affect Paralogue Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Range, F.; Marshall-Pescini, S. Comparing Wolves and Dogs: Current Status and Implications for Human ‘Self-Domestication’. Trends Cogn. Sci. 2022, 26, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, A.S.; Wrangham, R.W.; Fitch, W.T. The “Domestication Syndrome” in Mammals: A Unified Explanation Based on Neural Crest Cell Behavior and Genetics. Genetics 2014, 197, 795–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Villagra, M.R.; Geiger, M.; Schneider, R.A. The Taming of the Neural Crest: A Developmental Perspective on the Origins of Morphological Covariation in Domesticated Mammals. R. Soc. Open Sci. 2016, 3, 160107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkins, A.S. A Striking Example of Developmental Bias in an Evolutionary Process: The “Domestication Syndrome”. Evol. Dev. 2020, 22, 143–153. [Google Scholar] [CrossRef] [Green Version]
- Wilkins, A.S. Revisiting two hypotheses on the “domestication syndrome” in light of genomic data. Vavilov J. Genet. Breed 2017, 21, 435–442. [Google Scholar] [CrossRef]
- Hare, B. Survival of the Friendliest: Homo Sapiens Evolved via Selection for Prosociality. Annu. Rev. Psychol. 2017, 68, 155–186. [Google Scholar] [CrossRef]
- Sánchez-Villagra, M.R.; van Schaik, C.P. Evaluating the Self-Domestication Hypothesis of Human Evolution. Evol. Anthropol. Issues News Rev. 2019, 28, 133–143. [Google Scholar] [CrossRef]
- Hare, B.; Wobber, V.; Wrangham, R. The Self-Domestication Hypothesis: Evolution of Bonobo Psychology Is Due to Selection against Aggression. Anim. Behav. 2012, 83, 573–585. [Google Scholar] [CrossRef]
- Niego, A.; Benítez-Burraco, A. Williams Syndrome, Human Self-Domestication, and Language Evolution. Front. Psychol. 2019, 10, 521. [Google Scholar] [CrossRef] [Green Version]
- Spikins, P.; French, J.C.; John-Wood, S.; Dytham, C. Theoretical and Methodological Approaches to Ecological Changes, Social Behaviour and Human Intergroup Tolerance 300,000 to 30,000 BP. J. Archaeol. Method Theory 2021, 28, 53–75. [Google Scholar] [CrossRef]
- Brooks, J.; Yamamoto, S. The Founder Sociality Hypothesis. Ecol. Evol. 2021, 11, 14392–14404. [Google Scholar] [CrossRef] [PubMed]
- Theofanopoulou, C.; Gastaldon, S.; O’Rourke, T.; Samuels, B.D.; Messner, A.; Martins, P.T.; Delogu, F.; Alamri, S.; Boeckx, C. Self-Domestication in Homo Sapiens: Insights from Comparative Genomics. PLoS ONE 2017, 12, e0185306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanella, M.; Vitriolo, A.; Andirko, A.; Martins, P.T.; Sturm, S.; O’Rourke, T.; Laugsch, M.; Malerba, N.; Skaros, A.; Trattaro, S.; et al. Dosage Analysis of the 7q11.23 Williams Region Identifies BAZ1B as a Major Human Gene Patterning the Modern Human Face and Underlying Self-Domestication. Sci. Adv. 2019, 5, eaaw7908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkins, A.S. A Molecular Investigation of Human Self-Domestication. Trends Genet. 2020, 36, 227–228. [Google Scholar] [CrossRef]
- Shilton, D.; Breski, M.; Dor, D.; Jablonka, E. Human Social Evolution: Self-Domestication or Self-Control? Front. Psychol. 2020, 11, 134. [Google Scholar] [CrossRef]
- Gleeson, B.T.; Kushnick, G. Female Status, Food Security, and Stature Sexual Dimorphism: Testing Mate Choice as a Mechanism in Human Self-Domestication. Am. J. Phys. Anthr. 2018, 167, 458–469. [Google Scholar] [CrossRef]
- Gibson, G.; Lacek, K.A. Canalization and Robustness in Human Genetics and Disease. Annu. Rev. Genet. 2020, 54, 189–211. [Google Scholar] [CrossRef]
- Benton, M.L.; Abraham, A.; LaBella, A.L.; Abbot, P.; Rokas, A.; Capra, J.A. The Influence of Evolutionary History on Human Health and Disease. Nat. Rev. Genet. 2021, 22, 269–283. [Google Scholar] [CrossRef]
- Benítez-Burraco, A.; Lattanzi, W.; Murphy, E. Language Impairments in ASD Resulting from a Failed Domestication of the Human Brain. Front. Neurosci. 2016, 10, 373. [Google Scholar] [CrossRef] [Green Version]
- Benítez-Burraco, A.; Di Pietro, L.; Barba, M.; Lattanzi, W. Schizophrenia and Human Self-Domestication: An Evolutionary Linguistics Approach. Brain. Behav. Evol. 2017, 89, 162–184. [Google Scholar] [CrossRef]
- Belyaev, D.K.; Ruvinsky, A.O.; Trut, L.N. Inherited Activation-Inactivation of the Star Gene in Foxes: Its Bearing on the Problem of Domestication. J. Hered. 1981, 72, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Trut, L.; Oskina, I.; Kharlamova, A. Animal Evolution during Domestication: The Domesticated Fox as a Model. BioEssays 2009, 31, 349–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, I.J.; Clark, M.M.; Thompson, M.J.; Deere-Machemer, K.A.; Wang, J.; Duarte, L.; Gnanadesikan, G.E.; McCoy, E.L.; Rubbi, L.; Stahler, D.R.; et al. The Concerted Impact of Domestication and Transposon Insertions on Methylation Patterns between Dogs and Grey Wolves. Mol. Ecol. 2016, 25, 1838–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogt, G. Facilitation of Environmental Adaptation and Evolution by Epigenetic Phenotype Variation: Insights from Clonal, Invasive, Polyploid, and Domesticated Animals. Environ. Epigenetics 2017, 3, dvx002. [Google Scholar] [CrossRef]
- Vogt, G. Epigenetic Variation in Animal Populations: Sources, Extent, Phenotypic Implications, and Ecological and Evolutionary Relevance. J. Biosci. 2021, 46, 24. [Google Scholar] [CrossRef]
- Jablonka, E. The Evolutionary Implications of Epigenetic Inheritance. Interface Focus 2017, 7, 20160135. [Google Scholar] [CrossRef]
- Shea, N.; Pen, I.; Uller, T. Three Epigenetic Information Channels and Their Different Roles in Evolution: Epigenetic Mechanisms and Evolution. J. Evol. Biol. 2011, 24, 1178–1187. [Google Scholar] [CrossRef] [Green Version]
- Day, T.; Bonduriansky, R. A Unified Approach to the Evolutionary Consequences of Genetic and Nongenetic Inheritance. Am. Nat. 2011, 178, E18–E36. [Google Scholar] [CrossRef]
- Anastasiadi, D.; Venney, C.J.; Bernatchez, L.; Wellenreuther, M. Epigenetic Inheritance and Reproductive Mode in Plants and Animals. Trends Ecol. Evol. 2021, 36, 1124–1140. [Google Scholar] [CrossRef]
- Kronholm, I.; Collins, S. Epigenetic Mutations Can Both Help and Hinder Adaptive Evolution. Mol. Ecol. 2016, 25, 1856–1868. [Google Scholar] [CrossRef]
- O’Dea, R.E.; Noble, D.W.A.; Johnson, S.L.; Hesselson, D.; Nakagawa, S. The Role of Non-Genetic Inheritance in Evolutionary Rescue: Epigenetic Buffering, Heritable Bet Hedging and Epigenetic Traps. Environ. Epigenetics 2016, 2, dvv014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danchin, E.; Pocheville, A.; Rey, O.; Pujol, B.; Blanchet, S. Epigenetically Facilitated Mutational Assimilation: Epigenetics as a Hub within the Inclusive Evolutionary Synthesis. Biol. Rev. 2019, 94, 259–282. [Google Scholar] [CrossRef] [Green Version]
- Belteky, J.; Agnvall, B.; Bektic, L.; Hoglund, A.; Jensen, P.; Guerrero-Bosagna, C. Epigenetics and Early Domestication: Differences in Hypothalamic DNA Methylation between Red Junglefowl Divergently Selected for High or Low Fear of Humans. Genet. Sel. Evol. 2018, 50, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gokhman, D.; Nissim-Rafinia, M.; Agranat-Tamir, L.; Housman, G.; García-Pérez, R.; Lizano, E.; Cheronet, O.; Mallick, S.; Nieves-Colón, M.A.; Li, H.; et al. Differential DNA Methylation of Vocal and Facial Anatomy Genes in Modern Humans. Nat. Commun. 2020, 11, 1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teletchea, F. Domestication and Genetics: What a Comparison Between Land and Aquatic Species Can Bring? In Evolutionary Biology: Biodiversification from Genotype to Phenotype; Pontarotti, P., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 389–401. ISBN 978-3-319-19932-0. [Google Scholar]
- Teletchea, F. Fish Domestication in Aquaculture: 10 Unanswered Questions. Anim. Front. 2021, 11, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Teletchea, F. Fish Domestication in Aquaculture: Reassessment and Emerging Questions. Cybium 2019, 43, 7–15. [Google Scholar] [CrossRef]
- Houston, R.D.; Bean, T.P.; Macqueen, D.J.; Gundappa, M.K.; Jin, Y.H.; Jenkins, T.L.; Selly, S.L.C.; Martin, S.A.M.; Stevens, J.R.; Santos, E.M.; et al. Harnessing Genomics to Fast-Track Genetic Improvement in Aquaculture. Nat. Rev. Genet. 2020, 21, 389–409. [Google Scholar] [CrossRef]
- Anastasiadi, D.; Piferrer, F. Epimutations in Developmental Genes Underlie the Onset of Domestication in Farmed European Sea Bass. Mol. Biol. Evol. 2019, 36, 2252–2264. [Google Scholar] [CrossRef]
- Wringe, B.F.; Purchase, C.F.; Fleming, I.A. In Search of a “Cultured Fish Phenotype”: A Systematic Review, Meta-Analysis and Vote-Counting Analysis. Rev. Fish Biol. Fish. 2016, 26, 351–373. [Google Scholar] [CrossRef]
- Milla, S.; Pasquet, A.; El Mohajer, L.; Fontaine, P. How Domestication Alters Fish Phenotypes. Rev. Aquac. 2021, 13, 388–405. [Google Scholar] [CrossRef]
- Sarropoulou, E.; Fernandes, J.M.O. Comparative Genomics in Teleost Species: Knowledge Transfer by Linking the Genomes of Model and Non-Model Fish Species. Comp. Biochem. Physiol. Part D Genom. Proteom. 2011, 6, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.F.; Jehangir, M.; Srikulnath, K.; Martins, C. Fish Genomics and Its Impact on Fundamental and Applied Research of Vertebrate Biology. Rev. Fish Biol. Fish. 2021, 32, 357–385. [Google Scholar] [CrossRef]
- Le Luyer, J.; Laporte, M.; Beacham, T.D.; Kaukinen, K.H.; Withler, R.E.; Leong, J.S.; Rondeau, E.B.; Koop, B.F.; Bernatchez, L. Parallel Epigenetic Modifications Induced by Hatchery Rearing in a Pacific Salmon. Proc. Natl. Acad. Sci. USA 2017, 114, 12964–12969. [Google Scholar] [CrossRef] [Green Version]
- Wellband, K.; Roth, D.; Linnansaari, T.; Curry, R.A.; Bernatchez, L. Environment-Driven Reprogramming of Gamete DNA Methylation Occurs during Maturation and Is Transmitted Intergenerationally in Atlantic Salmon. G3 Genes Genomes Genet. 2021, 11, jkab353. [Google Scholar] [CrossRef] [PubMed]
- Konstantinidis, I.; Sætrom, P.; Mjelle, R.; Nedoluzhko, A.V.; Robledo, D.; Fernandes, J.M.O. Major Gene Expression Changes and Epigenetic Remodelling in Nile Tilapia Muscle after Just One Generation of Domestication. Epigenetics 2020, 15, 1052–1067. [Google Scholar] [CrossRef] [Green Version]
- Podgorniak, T.; Dhanasiri, A.; Chen, X.; Ren, X.; Kuan, P.-F.; Fernandes, J. Early Fish Domestication Affects Methylation of Key Genes Involved in the Rapid Onset of the Farmed Phenotype. Epigenetics 2022, 1–18. [Google Scholar] [CrossRef]
- Li, Y.; Fu, B.; Zhang, J.; Xie, J.; Wang, G.; Jiang, P.; Tian, J.; Li, H.; Xia, Y.; Yu, E. Genome-wide analysis of DNA methylation reveals selection signatures of grass carp during the domestication. bioRxiv 2021. [Google Scholar] [CrossRef]
- Leitwein, M.; Laporte, M.; Le Luyer, J.; Mohns, K.; Normandeau, E.; Withler, R.; Bernatchez, L. Epigenomic Modifications Induced by Hatchery Rearing Persist in Germ Line Cells of Adult Salmon after Their Oceanic Migration. Evol. Appl. 2021, 14, 2402–2413. [Google Scholar] [CrossRef]
- Rodriguez Barreto, D.; Garcia de Leaniz, C.; Verspoor, E.; Sobolewska, H.; Coulson, M.; Consuegra, S. DNA Methylation Changes in the Sperm of Captive-Reared Fish: A Route to Epigenetic Introgression in Wild Populations. Mol. Biol. Evol. 2019, 36, 2205–2211. [Google Scholar] [CrossRef] [Green Version]
- Gavery, M.R.; Nichols, K.M.; Goetz, G.W.; Middleton, M.A.; Swanson, P. Characterization of Genetic and Epigenetic Variation in Sperm and Red Blood Cells from Adult Hatchery and Natural-Origin Steelhead, Oncorhynchus Mykiss. G3 Genes Genomes Genet. 2018, 8, 3723–3736. [Google Scholar] [CrossRef] [Green Version]
- Bertolini, F.; Geraci, C.; Schiavo, G.; Sardina, M.T.; Chiofalo, V.; Fontanesi, L. Whole Genome Semiconductor Based Sequencing of Farmed European Sea Bass (Dicentrarchus Labrax) Mediterranean Genetic Stocks Using a DNA Pooling Approach. Mar. Genom. 2016, 28, 63–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Rourke, T.; Boeckx, C. Glutamate Receptors in Domestication and Modern Human Evolution. Neurosci. Biobehav. Rev. 2020, 108, 341–357. [Google Scholar] [CrossRef] [PubMed]
- Strong, E.; Butcher, D.T.; Singhania, R.; Mervis, C.B.; Morris, C.A.; De Carvalho, D.; Weksberg, R.; Osborne, L.R. Symmetrical Dose-Dependent DNA-Methylation Profiles in Children with Deletion or Duplication of 7q11.23. Am. J. Hum. Genet. 2015, 97, 216–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, R.; Lardenoije, R.; Tomiwa, K.; Funabiki, Y.; Nakata, M.; Suzuki, S.; Awaya, T.; Kato, T.; Okazaki, S.; Murai, T.; et al. Integrated DNA Methylation Analysis Reveals a Potential Role for ANKRD30B in Williams Syndrome. Neuropsychopharmacology 2020, 45, 1627–1636. [Google Scholar] [CrossRef]
- Forero, D.A.; González-Giraldo, Y. Integrative In Silico Analysis of Genome-Wide DNA Methylation Profiles in Schizophrenia. J. Mol. Neurosci. 2020, 70, 1887–1893. [Google Scholar] [CrossRef]
- Hu, V.W.; Hong, Y.; Xu, M.; Shu, H.T. Altered DNA Methylation in a Severe Subtype of Idiopathic Autism: Evidence for Sex Differences in Affected Metabolic Pathways. Autism 2021, 25, 887–910. [Google Scholar] [CrossRef]
- Zhang, B.; Hu, X.; Li, Y.; Ni, Y.; Xue, L. Identification of Methylation Markers for Diagnosis of Autism Spectrum Disorder. Metab. Brain Dis. 2021, 37, 219–228. [Google Scholar] [CrossRef]
- Lee, E.C.; Hu, V.W. Phenotypic Subtyping and Re-Analysis of Existing Methylation Data from Autistic Probands in Simplex Families Reveal ASD Subtype-Associated Differentially Methylated Genes and Biological Functions. Int. J. Mol. Sci. 2020, 21, 6877. [Google Scholar] [CrossRef]
- Nardone, S.; Sams, D.S.; Zito, A.; Reuveni, E.; Elliott, E. Dysregulation of Cortical Neuron DNA Methylation Profile in Autism Spectrum Disorder. Cereb. Cortex 2017, 27, 5739–5754. [Google Scholar] [CrossRef] [Green Version]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A Web-Based Tool for the Analysis of Sets through Venn Diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- R Studio Team. R Studio: Integrated Development Environment for R; R Studio, Inc.: Boston, MA, USA, 2015. [Google Scholar]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and Collaborative HTML5 Gene List Enrichment Analysis Tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A Comprehensive Gene Set Enrichment Analysis Web Server 2016 Update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef] [PubMed]
- Supek, F.; Bošnjak, M.; Škunca, N.; Šmuc, T. REVIGO Summarizes and Visualizes Long Lists of Gene Ontology Terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montague, M.J.; Li, G.; Gandolfi, B.; Khan, R.; Aken, B.L.; Searle, S.M.J.; Minx, P.; Hillier, L.W.; Koboldt, D.C.; Davis, B.W.; et al. Comparative Analysis of the Domestic Cat Genome Reveals Genetic Signatures Underlying Feline Biology and Domestication. Proc. Natl. Acad. Sci. USA 2014, 111, 17230–17235. [Google Scholar] [CrossRef] [Green Version]
- Pendleton, A.L.; Shen, F.; Taravella, A.M.; Emery, S.; Veeramah, K.R.; Boyko, A.R.; Kidd, J.M. Comparison of Village Dog and Wolf Genomes Highlights the Role of the Neural Crest in Dog Domestication. BMC Biol. 2018, 16, 64. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Jia, Y.; Almeida, P.; Mank, J.E.; van Tuinen, M.; Wang, Q.; Jiang, Z.; Chen, Y.; Zhan, K.; Hou, S. Whole-Genome Resequencing Reveals Signatures of Selection and Timing of Duck Domestication. GigaScience 2018, 7, giy027. [Google Scholar] [CrossRef]
- Kukekova, A.V.; Johnson, J.L.; Xiang, X.; Feng, S.; Liu, S.; Rando, H.M.; Kharlamova, A.V.; Herbeck, Y.; Serdyukova, N.A.; Xiong, Z.; et al. Red Fox Genome Assembly Identifies Genomic Regions Associated with Tame and Aggressive Behaviours. Nat. Ecol. Evol. 2018, 2, 1479–1491. [Google Scholar] [CrossRef]
- Carneiro, M.; Rubin, C.-J.; Di Palma, F.; Albert, F.W.; Alföldi, J.; Barrio, A.M.; Pielberg, G.; Rafati, N.; Sayyab, S.; Turner-Maier, J. Rabbit Genome Analysis Reveals a Polygenic Basis for Phenotypic Change during Domestication. Science 2014, 345, 1074–1079. [Google Scholar] [CrossRef] [Green Version]
- Benítez-Burraco, A.; Chekalin, E.; Bruskin, S.; Tatarinova, T.; Morozova, I. Recent Selection of Candidate Genes for Mammal Domestication in Europeans and Language Change in Europe: A Hypothesis. Ann. Hum. Biol. 2021, 48, 313–320. [Google Scholar] [CrossRef]
- Torres-Pérez, J.V.; Anagianni, S.; Mech, A.M.; Havelange, W.; García-González, J.; Fraser, S.E.; Vallortigara, G.; Brennan, C.H. Behavioural Analysis of Loss of Function Zebrafish Supports baz1b as Master Regulator of Domestication. bioRxiv 2022. [Google Scholar] [CrossRef]
- Matissek, S.J.; Elsawa, S.F. GLI3: A Mediator of Genetic Diseases, Development and Cancer. Cell Commun. Signal. CCS 2020, 18, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddad-Tóvolli, R.; Heide, M.; Zhou, X.; Blaess, S.; Alvarez-Bolado, G. Mouse Thalamic Differentiation: Gli-Dependent Pattern and Gli-Independent Prepattern. Front. Neurosci. 2012, 6, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanimoto, Y.; Veistinen, L.; Alakurtti, K.; Takatalo, M.; Rice, D.P.C. Prevention of Premature Fusion of Calvarial Suture in GLI-Kruppel Family Member 3 (Gli3)-Deficient Mice by Removing One Allele of Runt-Related Transcription Factor 2 (Runx2). J. Biol. Chem. 2012, 287, 21429–21438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellano, S.; Parra, G.; Sánchez-Quinto, F.A.; Racimo, F.; Kuhlwilm, M.; Kircher, M.; Sawyer, S.; Fu, Q.; Heinze, A.; Nickel, B.; et al. Patterns of Coding Variation in the Complete Exomes of Three Neandertals. Proc. Natl. Acad. Sci. USA 2014, 111, 6666–6671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, K.; Shiga, T.; Ito, Y. Runx Transcription Factors in Neuronal Development. Neural Dev. 2008, 3, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boeckx, C.; Benítez-Burraco, A. Globularity and Language-Readiness: Generating New Predictions by Expanding the Set of Genes of Interest. Front. Psychol. 2014, 5, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Takahata, Y.; Hagino, H.; Kimura, A.; Urushizaki, M.; Kobayashi, S.; Wakamori, K.; Fujiwara, C.; Nakamura, E.; Yu, K.; Kiyonari, H.; et al. Smoc1 and Smoc2 Regulate Bone Formation as Downstream Molecules of Runx2. Commun. Biol. 2021, 4, 1199. [Google Scholar] [CrossRef]
- Williams, S.S.; Mear, J.P.; Liang, H.-C.; Potter, S.S.; Aronow, B.J.; Colbert, M.C. Large-Scale Reprogramming of Cranial Neural Crest Gene Expression by Retinoic Acid Exposure. Physiol. Genom. 2004, 19, 184–197. [Google Scholar] [CrossRef] [Green Version]
- Dityatev, A.; Seidenbecher, C.; Morawski, M. Brain Extracellular Matrix: An Upcoming Target in Neurological and Psychiatric Disorders. Eur. J. Neurosci. 2021, 53, 3807–3810. [Google Scholar] [CrossRef]
- Šimić, G.; Vukić, V.; Kopić, J.; Krsnik, Ž.; Hof, P.R. Molecules, Mechanisms, and Disorders of Self-Domestication: Keys for Understanding Emotional and Social Communication from an Evolutionary Perspective. Biomolecules 2020, 11, 2. [Google Scholar] [CrossRef] [PubMed]
- Rohner, N.; Bercsényi, M.; Orbán, L.; Kolanczyk, M.E.; Linke, D.; Brand, M.; Nüsslein-Volhard, C.; Harris, M.P. Duplication of Fgfr1 Permits Fgf Signaling to Serve as a Target for Selection during Domestication. Curr. Biol. CB 2009, 19, 1642–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meshalkina, D.A.; Kizlyk, M.N.; Kysil, E.V.; Collier, A.D.; Echevarria, D.J.; Abreu, M.S.; Barcellos, L.J.G.; Song, C.; Warnick, J.E.; Kyzar, E.J.; et al. Zebrafish Models of Autism Spectrum Disorder. Exp. Neurol. 2018, 299, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Won, S.Y.; Choi, B.-O.; Chung, K.W.; Lee, J.E. Zebrafish Is a Central Model to Dissect the Peripheral Neuropathy. Genes Genom. 2019, 41, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Bandmann, O.; Burton, E.A. Genetic Zebrafish Models of Neurodegenerative Diseases. Neurobiol. Dis. 2010, 40, 58–65. [Google Scholar] [CrossRef]
- Choi, T.-Y.; Choi, T.-I.; Lee, Y.-R.; Choe, S.-K.; Kim, C.-H. Zebrafish as an Animal Model for Biomedical Research. Exp. Mol. Med. 2021, 53, 310–317. [Google Scholar] [CrossRef]
- Schartl, M. Beyond the Zebrafish: Diverse Fish Species for Modeling Human Disease. Dis. Model. Mech. 2014, 7, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Albertson, R.C.; Cresko, W.; Detrich, H.W.; Postlethwait, J.H. Evolutionary Mutant Models for Human Disease. Trends Genet. TIG 2009, 25, 74–81. [Google Scholar] [CrossRef] [Green Version]
- Beck, E.A.; Healey, H.M.; Small, C.M.; Currey, M.C.; Desvignes, T.; Cresko, W.A.; Postlethwait, J.H. Advancing Human Disease Research with Fish Evolutionary Mutant Models. Trends Genet. 2022, 38, 22–44. [Google Scholar] [CrossRef]
- Parker, M.O.; Brock, A.J.; Walton, R.T.; Brennan, C.H. The Role of Zebrafish (Danio Rerio) in Dissecting the Genetics and Neural Circuits of Executive Function. Front. Neural Circuits 2013, 7, 63. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, C.; Kim, C.H. Zebrafish as a Model for Infectious Disease and Immune Function. Fish Shellfish. Immunol. 2008, 25, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The Zebrafish Reference Genome Sequence and Its Relationship to the Human Genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, H. The Use of Fish Models to Study Human Neurological Disorders. Neurosci. Res. 2017, 120, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Langova, V.; Vales, K.; Horka, P.; Horacek, J. The Role of Zebrafish and Laboratory Rodents in Schizophrenia Research. Front. Psychiatry 2020, 11, 703. [Google Scholar] [CrossRef]
- Komoike, Y.; Fujii, K.; Nishimura, A.; Hiraki, Y.; Hayashidani, M.; Shimojima, K.; Nishizawa, T.; Higashi, K.; Yasukawa, K.; Saitsu, H.; et al. Zebrafish Gene Knockdowns Imply Roles for Human YWHAG in Infantile Spasms and Cardiomegaly. Genesis 2010, 48, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Rea, V.; Van Raay, T.J. Using Zebrafish to Model Autism Spectrum Disorder: A Comparison of ASD Risk Genes Between Zebrafish and Their Mammalian Counterparts. Front. Mol. Neurosci. 2020, 13, 575575. [Google Scholar] [CrossRef]
- Eckhard Witten, P.; Leonor Cancela, M. What Aquaculture Does for Taxonomy, Evo-Devo, Palaeontology, Biomechanics and Biomedical Research. J. Appl. Ichthyol. 2018, 34, 429–430. [Google Scholar] [CrossRef]
- Sánchez-Villagra, M. The Process of Animal Domestication; Princeton University Press: Princeton, NJ, USA, 2022; ISBN 978-0-691-21768-0. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Description | Ensembl Gene Stable ID |
|---|---|---|
| adamts17 agap1 atp7b | ADAM metallopeptidase with thrombospondin type 1 motif, 17 | ENSDLAG00005007818 |
| ArfGAP with GTPase domain, ankyrin repeat and PH domain 1 | ENSDLAG00005018378 | |
| ATPase copper transporting beta | ENSDLAG00005026064 | |
| bcr carm1 | BCR activator of RhoGEF and GTPase | ENSDLAG00005004082 |
| coactivator-associated arginine methyltransferase 1 | ENSDLAG00005025319 | |
| celsr1a cemip coro7 dab2ipb | cadherin EGF LAG seven-pass G-type receptor 1a | ENSDLAG00005009488 |
| cell migration inducing hyaluronidase 1 | ENSDLAG00005002105 | |
| coronin 7 | ENSDLAG00005014078 | |
| DAB2 interacting protein b | ENSDLAG00005020932 | |
| DIP2C ephb3a | disco-interacting protein 2 homolog Ca | ENSDLAG00005023732 |
| eph receptor B3a | ENSDLAG00005000091 | |
| eps8l2 | EPS8 like 2 | ENSDLAG00005011013 |
| EYA2 | EYA transcriptional coactivator and phosphatase 2 | ENSDLAG00005013401 |
| fbrsl1 | fibrosin-like 1 | ENSDLAG00005019385 |
| fgfrl1a | fibroblast growth factor receptor like 1a | ENSDLAG00005002545 |
| galnt18a | UDP-N-acetylalphaDgalactosamine:polypeptideN-acetylgalactosaminyltransferase 18a | ENSDLAG00005020537 |
| gli3 | GLI family zinc finger 3 | ENSDLAG00005018034 |
| itga11b | integrin, alpha 11b | ENSDLAG00005013142 |
| kaznb | kazrin, periplakin interacting protein b | ENSDLAG00005001674 |
| lasp1 | LIM and SH3 protein 1 | ENSDLAG00005018795 |
| lhpp | phospholysine phosphohistidine inorganic pyrophosphate phosphatase | ENSDLAG00005006011 |
| lmx1bb | LIM homeobox transcription factor 1, beta b | ENSDLAG00005025877 |
| magi1b | membrane associated guanylate kinase, WW and PDZ domain containing 1b | ENSDLAG00005022108 |
| mast2 | microtubule associated serine/threonine kinase 2 | ENSDLAG00005007444 |
| meis2a | Meis homeobox 2a | ENSDLAG00005007335 |
| msmo1 | methylsterol monooxygenase 1 | ENSDLAG00005023171 |
| ncor2 | nuclear receptor corepressor 2 | ENSDLAG00005024501 |
| neurl1aa | neuralized E3 ubiquitin protein ligase 1Aa | ENSDLAG00005018019 |
| nfixb | nuclear factor I X | ENSDLAG00005016844 |
| pacs2 | phosphofurin acidic cluster sorting protein 2 | ENSDLAG00005000298 |
| parvb | parvin, beta | ENSDLAG00005021030 |
| phactr3b | phosphatase and actin regulator 3b | ENSDLAG00005012177 |
| prex1 | phosphatidylinositol-3,4,5-trisphosphate-dependent Rac exchange factor 1 | ENSDLAG00005024474 |
| rab3il1 | RAB3A interacting protein (rabin3)-like 1 | ENSDLAG00005012050 |
| runx3 | RUNX family transcription factor 3 | ENSDLAG00005000657 |
| sh3pxd2aa | SH3 and PX domains 2Aa | ENSDLAG00005018046 |
| si:ch211–243o19.4 | si:ch211–243o19.4 | ENSDLAG00005011826 |
| smoc1 | SPARC related modular calcium binding 1 | ENSDLAG00005010838 |
| sorcs2 | sortilin-related VPS10 domain containing receptor 2 | ENSDLAG00005020908 |
| tbc1d22a | TBC1 domain family, member 22a | ENSDLAG00005010430 |
| tgfbr2b | transforming growth factor beta receptor 2b | ENSDLAG00005010792 |
| ZNF423 | zinc finger protein 423 | ENSDLAG00005008914 |
| ENSDLAG00005005197 | ||
| ENSDLAG00005012304 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anastasiadi, D.; Piferrer, F.; Wellenreuther, M.; Benítez Burraco, A. Fish as Model Systems to Study Epigenetic Drivers in Human Self-Domestication and Neurodevelopmental Cognitive Disorders. Genes 2022, 13, 987. https://doi.org/10.3390/genes13060987
Anastasiadi D, Piferrer F, Wellenreuther M, Benítez Burraco A. Fish as Model Systems to Study Epigenetic Drivers in Human Self-Domestication and Neurodevelopmental Cognitive Disorders. Genes. 2022; 13(6):987. https://doi.org/10.3390/genes13060987
Chicago/Turabian StyleAnastasiadi, Dafni, Francesc Piferrer, Maren Wellenreuther, and Antonio Benítez Burraco. 2022. "Fish as Model Systems to Study Epigenetic Drivers in Human Self-Domestication and Neurodevelopmental Cognitive Disorders" Genes 13, no. 6: 987. https://doi.org/10.3390/genes13060987
APA StyleAnastasiadi, D., Piferrer, F., Wellenreuther, M., & Benítez Burraco, A. (2022). Fish as Model Systems to Study Epigenetic Drivers in Human Self-Domestication and Neurodevelopmental Cognitive Disorders. Genes, 13(6), 987. https://doi.org/10.3390/genes13060987