
Human Fetal Liver Parenchyma CD71+ Cells Have AIRE and Tissue-Specific Antigen Gene Expression

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. NanoString Data Analysis
2.2. Study Population
2.3. Cell Isolation
2.4. Cell Sorting
2.5. Viability Staining
2.6. Flow Cytometry
2.7. Total RNA Extraction
2.8. Reverse Transcription
2.9. PCR and Melt Curve Analysis
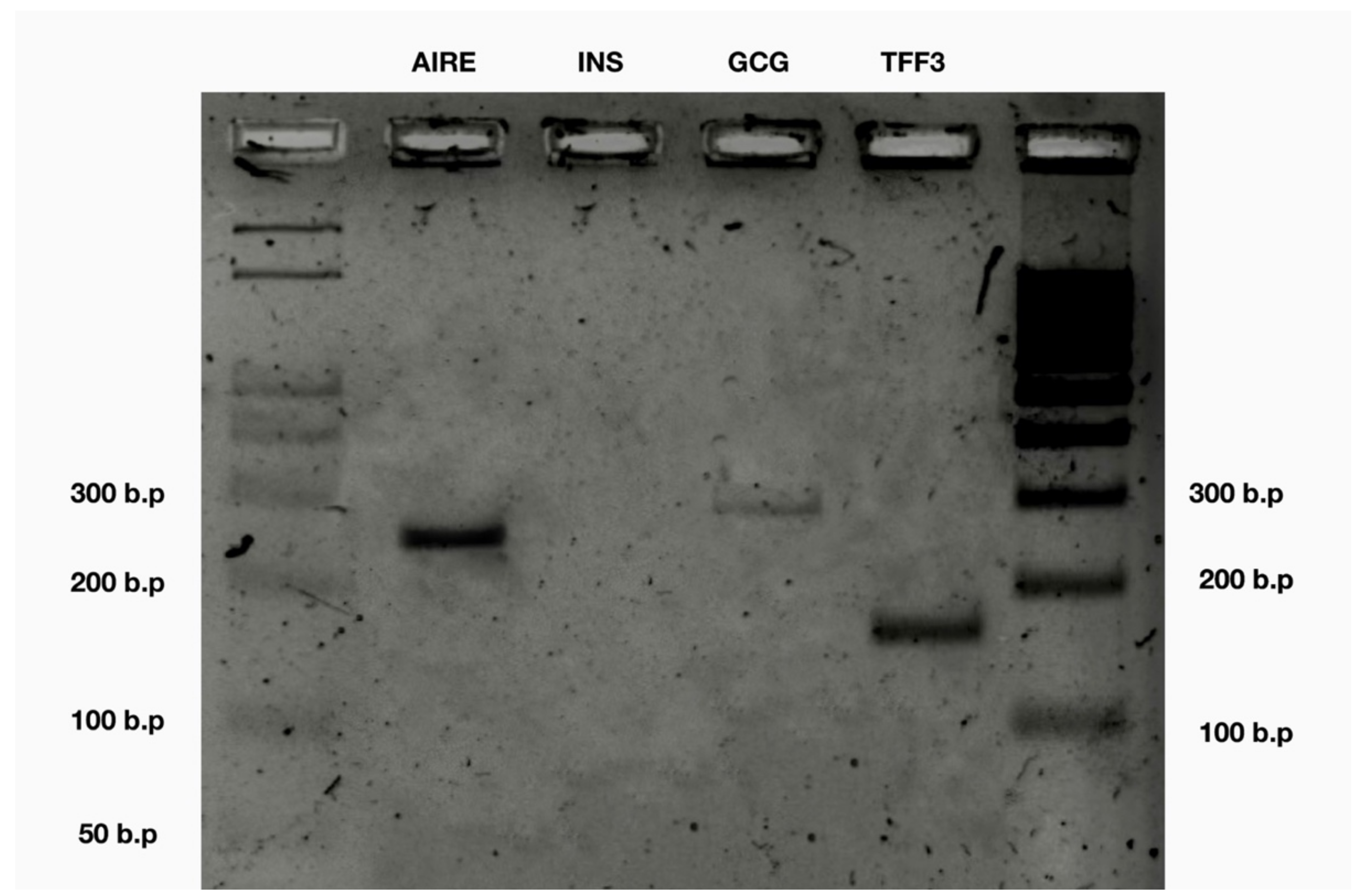
2.10. Agarose Gel-Electrophoresis of PCR Products
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shevyrev, D.; Tereshchenko, V.; Kozlov, V.; Sennikov, S. Phylogeny, Structure, Functions, and Role of AIRE in the Formation of T-Cell Subsets. Cells 2022, 11, 194. [Google Scholar] [CrossRef] [PubMed]
- Perniola, R. Twenty years of AIRE. Front. Immunol. 2018, 9, 98. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.S.; Venanzi, E.S.; Klein, L.; Chen, Z.; Berzins, S.P.; Turley, S.J.; von Boehmer, H.; Bronson, R.; Dierich, A.; Benoist, C.; et al. Projection of an immunological self shadow within the thymus by the aire protein. Science 2002, 298, 1395–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derbinski, J.; Schulte, A.; Kyewski, B.; Klein, L. Promiscuous gene expression in medullary thymic epithelial cells mirrors the peripheral self. Nat. Immunol. 2001, 2, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Hubert, F.X.; Kinkel, S.A.; Davey, G.M.; Phipson, B.; Mueller, S.N.; Liston, A.; Proietto, A.I.; Cannon, P.Z.; Forehan, S.; Smyth, G.K.; et al. Aire regulates the transfer of antigen from mTECs to dendritic cells for induction of thymic tolerance. Blood J. Am. Soc. Hematol. 2011, 118, 2462–2472. [Google Scholar] [CrossRef] [PubMed]
- Nishijima, H.; Kitano, S.; Miyachi, H.; Morimoto, J.; Kawano, H.; Hirota, F.; Morita, R.; Mouri, Y.; Masuda, K.; Imoto, I.; et al. Ectopic aire expression in the thymic cortex reveals inherent properties of aire as a tolerogenic factor within the medulla. J. Immunol. 2015, 195, 4641–4649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, J.M.; Fletcher, A.L.; Anderson, M.S.; Turley, S.J. AIRE in the thymus and beyond. Curr. Opin. Immunol. 2009, 21, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Gardner, J.M.; Metzger, T.C.; McMahon, E.J.; Au-Yeung, B.B.; Krawisz, A.K.; Lu, W.; Price, J.D.; Johannes, K.P.; Satpathy, A.T.; Murphy, K.M.; et al. Extrathymic Aire-expressing cells are a distinct bone marrow-derived population that induce functional inactivation of CD4+ T cells. Immunity 2013, 39, 560–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillis-Buck, E.; Miller, H.; Sirota, M.; Sanders, S.J.; Ntranos, V.; Anderson, M.S.; Gardner, J.M.; MacKenzie, T.C. Extrathymic Aire-expressing cells support maternal-fetal tolerance. Sci. Immunol. 2021, 6, eabf1968. [Google Scholar] [CrossRef] [PubMed]
- Kont, V.; Laan, M.; Kisand, K.; Merits, A.; Scott, H.S.; Peterson, P. Modulation of Aire regulates the expression of tissue-restricted antigens. Mol. Immunol. 2008, 45, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Sennikov, S.V.; Eremina, L.V.; Samarin, D.M.; Avdeev, I.V.; Kozlov, V.A. Cytokine gene expression in erythroid cells. Eur. Cytokine Netw. 1996, 7, 771–774. [Google Scholar] [PubMed]
- Sennikov, S.V.; Injelevskaya, T.V.; Krysov, S.V.; Silkov, A.N.; Kovinev, I.B.; Dyachkova, N.J.; Zenkov, A.N.; Loseva, M.I.; Kozlov, V.A. Production of hemo-and immunoregulatory cytokines by erythroblast antigen+ and glycophorin A+ cells from human bone marrow. BMC Cell Biol. 2004, 5, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sennikov, S.V.; Krysov, S.V.; Injelevskaya, T.V.; Silkov, A.N.; Kozlov, V.A. Production of cytokines by immature erythroid cells derived from human embryonic liver. Eur. Cytokine Netw. 2001, 12, 274–279. [Google Scholar] [PubMed]
- Denisova, V.V.; Kulagin, A.; Lisukov, I.A.; Kryuchkova, I.; Sizikova, S.; Sennikov, S.V.; Kozlov, V.A. Cytokine-producing activity of bone marrow erythrokaryocytes and its regulation under normal conditions. Bull. Exp. Biol. Med. 2007, 143, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Seledtsov, V.I.; Seledtsova, G.V.; Samarin, D.M.; Taraban, V.Y.; Sennikov, S.V.; Kozlov, V.A. Characterization of erythroid cell-derived natural suppressor activity. Immunobiology 1998, 198, 361–374. [Google Scholar] [CrossRef]
- Shahbaz, S.; Bozorgmehr, N.; Koleva, P.; Namdar, A.; Jovel, J.; Fava, R.A.; Elahi, S. CD71+ VISTA+ erythroid cells promote the development and function of regulatory T cells through TGF-β. PLoS Biol. 2018, 16, e2006649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meredith, M.M.; Zemmour, D.; Mathis, D.; Benoist, C. Aire controls gene expression in the thymic epithelium with ordered stochasticity. Nat. Immunol. 2015, 16, 942–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer | Amplicon Length | Melting Temperature | Accession |
|---|---|---|---|---|---|
| AIRE | catctcgaccacttttcagttcag | ccaccatgctgagtaaaataagacag | 250 b.p. | 82.0 °C | NM_000383.4 |
| GCG | ggtgtattctgaggccacattg | tgtggctaccagttcttctattctcc | 295 b.p. | 73.5 °C | NM_002054.5 |
| INS | ggagaactactgcaactagacgcag | ggttcaagggctttattccatctc | 89 b.p. | 82.5 °C | NM_000207.3 |
| TFF3 | cacccacgtcacaggaaagc | cgagagtggttgtgaaataaaggac | 170 b.p. | 78.5 °C | NM_003226.4 |
| Human Fetal Liver Parenchyma CD71+ Cells | ||
|---|---|---|
| Cytokine | Presence | № of Samples Positive |
| AIRE | + | 6/6 |
| INS | − | 0/6 |
| TFF3 | + | 6/6 |
| GCG | + | 6/6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perik-Zavodskii, R.; Perik-Zavodskaya, O.; Shevchenko, Y.; Alrhmoun, S.; Volynets, M.; Zaitsev, K.; Sennikov, S. Human Fetal Liver Parenchyma CD71+ Cells Have AIRE and Tissue-Specific Antigen Gene Expression. Genes 2022, 13, 1278. https://doi.org/10.3390/genes13071278
Perik-Zavodskii R, Perik-Zavodskaya O, Shevchenko Y, Alrhmoun S, Volynets M, Zaitsev K, Sennikov S. Human Fetal Liver Parenchyma CD71+ Cells Have AIRE and Tissue-Specific Antigen Gene Expression. Genes. 2022; 13(7):1278. https://doi.org/10.3390/genes13071278
Chicago/Turabian StylePerik-Zavodskii, Roman, Olga Perik-Zavodskaya, Yulia Shevchenko, Saleh Alrhmoun, Marina Volynets, Konstantin Zaitsev, and Sergey Sennikov. 2022. "Human Fetal Liver Parenchyma CD71+ Cells Have AIRE and Tissue-Specific Antigen Gene Expression" Genes 13, no. 7: 1278. https://doi.org/10.3390/genes13071278