
The Methylation of the p53 Targets the Genes MIR-203, MIR-129-2, MIR-34A and MIR-34B/C in the Tumor Tissue of Diffuse Large B-Cell Lymphoma

 , ,
, , 
Abstract
:
1. Introduction
2. Results
2.1. Methylation of MIR-203, MIR-129-9, MIR-34A and MIR-34B/C Genes in Lymph Node Tissues with Reactive Follicular Hyperplasia
2.2. The MIR-203, MIR-129-2, MIR-34A and MIR-34B/C Methylation in the Lymphoma Samples
2.3. MIR-203, MIR-129-2, MIR-34A and MIR-34B/C Methylation Status and Clinical Features of DLBCL
2.4. Association of MIR-203, MIR-129-2, MIR-34A and MIR-34B/C Methylation with DLBCL Immunohistochemical Features
2.5. MIR-203, MIR-129-2, MIR-34A and MIR-34B/C Genes’ Methylation and Results of the Treatment of DLBCL Patients
3. Discussion
4. Materials and Methods
4.1. Tumor Samples
4.2. Bisulfite Conversion of DNA Samples
4.3. The Assessment of Methylation Status of the MIR-34 Family Genes
4.4. Assessment of the Methylation Status of the MIR-203 and MIR-129-2 Genes
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABC | Activated B cell |
| BCL2 | B-cell lymphoma-2 |
| CDK | Cyclin-dependent kinase |
| DLBCL | Diffuse Large B-cell Lymphoma |
| ECOG | Eastern Cooperative Oncology Group |
| FFPE | Formalin-Fixed, Paraffin-Embedded |
| GCB | Germinal-Center B cell |
| IPI | International Prognostic Index |
| LDH | Lactate Dehydrogenase |
| MCL1 | Myeloid Cell Leukemia-1 |
| MDM2 | Mouse Double Minute-2 |
| MS-HRM | Methylation-Sensitive High-Resolution-Melting |
| MS-PCR | Methyl-Specific Polymerase Chain Reaction |
| OS | Overall Survival |
| PLCNS | Primary Lymphoma of the Central Nervous System |
| R-CHOEP | Rituximab, Cyclophosphamide, Doxorubicin, Vincristine, Etoposide, Prednisolone |
| R-CHOP | Rituximab, Cyclophosphamide, Doxorubicin, Vincristine, Prednisolone |
| WHO | World Health Organization |
References
- Syeda, A.Z.; Langden, S.S.S.; Munkhzul, C.; Lee, M.; Song, S.J. Regulatory Mechanism of MicroRNA Expression in Cancer. Int. J. Mol. Sci. 2020, 21, 1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aushev, V.N. MicroRNA: Small Molecules of Great Significance. Clin. Oncohematology 2015, 8, 1–12. (In Russian) [Google Scholar]
- Deng, G.; Sui, G. Noncoding RNA in oncogenesis: A new era of identifying key players. Int. J. Mol. Sci. 2013, 14, 18319–18349. [Google Scholar] [CrossRef] [PubMed]
- Walter, R.F.H.; Vollbrecht, C.; Werner, R.; Wohlschlaeger, J.; Christoph, D.C.; Schmid, K.W.; Mairinger, F.D. MicroRNAs are differentially regulated between MDM2-positive and negative malignant pleural mesothelioma. Oncotarget 2016, 7, 18713–18721. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Shen, H.; Peng, Q.; Yu, J. TP53/miR-129/MDM2/4/TP53 feedback loop modulates cell proliferation and apoptosis in retinoblastoma. Cell Cycle 2021, 20, 603–615. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, Q.; Zhang, Z. Loss of microRNA-143/145 disturbs cellular growth and apoptosis of human epithelial cancers by impairing the MDM2–p53 feedback loop. Oncogene 2013, 32, 61–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rokavec, M.; Li, H.; Jiang, L.; Hermeking, H. The p53/miR-34 axis in development and disease. J. Mol. Cell Biol. 2014, 3, 214–230. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.I.; Yamagata, K.; Sugimoto, K. Modulation of microRNA processing by p53. Nature 2009, 460, 529–533. [Google Scholar] [CrossRef]
- O’Connell, R.M.; Baltimore, D. MicroRNAs and hematopoietic cell development. Curr. Top. Dev. Biol. 2012, 99, 145–174. [Google Scholar] [CrossRef]
- Tokarz, P.; Pawlowska, E.; Bialkowska-Warzecha, J.; Blasiak, J. The significance of DNA methylation profile in metastasis-related genes for the progression of colorectal cancer. Cell. Mol. Biol. 2017, 63, 79–87. [Google Scholar] [CrossRef]
- Baylin, S.B.; Jones, P.A. Epigenetic Determinants of Cancer. Cold Spring Harb. Perspect. Biol. 2016, 8, a019505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solé, C.; Larrea, E.; Di Pinto, G.; Tellaetxe, M.; Lawrie, C.H. miRNAs in B-cell lymphoma: Molecular mechanisms and biomarker potential. Cancer Lett. 2017, 405, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, S.H.; Campo, E.; Piler, S.A.; Harris, N.L.; Stein, H.; Siebert, R.; Advani, R.; Ghielmini, M.; Salles, G.A.; Zelenetz, A.D.; et al. The 2016 revision of the World Health Organization classification of lymphoid neoplasms. Blood 2016, 127, 2375–2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehn, L.H.; Connors, J.M. Treatment of aggressive non-Hodgkin’s lymphoma: A north American perspective. Oncology 2005, 19 (4 Suppl 1), 26–34. [Google Scholar]
- Craig, V.J.; Cogliatti, S.B.; Rehrauer, H.; Wündisch, T.; Müller, A. Epigenetic silencing of microRNA-203 dysregulates ABL1 expression and drives Helicobacter-associated gastric lymphomagenesis. Cancer Res. 2011, 71, 3616–3624. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.-Y.; Yim, R.L.-H.; Kwong, Y.-L.; Leung, C.-Y.; Hui, P.-K.; Cheung, F.; Liang, R.; Jin, D.-Y.; Chim, C.-S. Epigenetic inactivation of the MIR129-2 in hematological malignancies. J. Hematol. Oncol. 2013, 6, 16. [Google Scholar] [CrossRef] [Green Version]
- Asmar, F.; Hother, C.; Kulosman, G.; Treppendahl, M.B.; Nielsen, H.M.; Ralfkiaer, U.; Pedersen, A.; Møller, M.B.; Ralfkiaer, E.; Brown, P.D.N.; et al. Diffuse large B-cell lymphoma with combined TP53 mutation and MIR34A methylation: Another “double hit” lymphoma with very poor outcome? Oncotarget 2014, 5, 1912–1925. [Google Scholar] [CrossRef] [Green Version]
- Arribas, A.J.; Gómez-Abad, C.; Sánchez-Beato, M.; Martinez, N.; Dilisio, L.; Casado, F.; A Cruz, M.; Algara, P.; A Piris, M.; Mollejo, M. Splenic marginal zone lymphoma: Comprehensive analysis of gene expression and miRNA profiling. Mod. Pathol. 2013, 26, 889–901. [Google Scholar] [CrossRef]
- Fischer, L.; Hummel, M.; Korfel, A.; Lenze, D.; Joehrens, K.; Thiel, E. Differential micro-RNA expression in primary CNS and nodal diffuse large B-cell lymphomas. Neuro-Oncology 2011, 13, 1090–1098. [Google Scholar] [CrossRef] [Green Version]
- Xia, H.; Yamada, S.; Aoyama, M.; Sato, F.; Masaki, A.; Ge, Y.; Ri, M.; Ishida, T.; Ueda, R.; Utsunomiya, A.; et al. Prognostic impact of microRNA-145 down-regulation in adult T-cell leukemia/lymphoma. Hum. Pathol. 2014, 45, 1192–1198. [Google Scholar] [CrossRef]
- Dos Santos Ferreira, A.C.; Robaina, M.C.; de Rezende, L.M.M.; Severino, P.; Klumb, C.E. Histone deacetylase inhibitor prevents cell growth in Burkitt’s lymphoma by regulating PI3K/Akt pathways and leads to upregulation of miR-143, miR-145, and miR-101. Ann. Hematol. 2014, 93, 983–993. [Google Scholar] [CrossRef] [PubMed]
- Chim, C.S.; Wong, K.Y. Epigenetic inactivation of the hsa-miR-203 in haematological malignancies. J. Cell. Mol. Med. 2011, 15, 2760–2767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artemaki, P.I.; Letsos, P.A.; Zoupa, I.C.; Katsaraki, K.; Karousi, P.; Papageorgiou, S.G.; Pappa, V.; Scorilas, A.; Kontos, C.K. The Multifaceted Role and Utility of MicroRNAs in Indolent B-Cell Non-Hodgkin Lymphomas. Biomedicines 2021, 9, 333. [Google Scholar] [CrossRef] [PubMed]
- Hermeking, H. The miR-34 family in cancer and apoptosis. Cell Death Differ. 2010, 17, 193–199. [Google Scholar] [CrossRef]
- Aukema, S.M.; Siebert, R.; Schuuring, E.; van Imhoff, G.W.; Kluin-Nelemans, H.C.; Boerma, E.J.; Kluin, P.M. Double-hit B-cell lymphomas. Blood 2011, 117, 2319–2331. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Chen, Z.; Fu, T.; Jin, X.; Yu, T.; Liang, Y.; Zhao, X.; Huang, L. Ki-67 is a valuable prognostic predictor of lymphoma but its utility varies in lymphoma subtypes: Evidence from a systematic meta-analysis. BMC Cancer 2014, 14, 153. [Google Scholar] [CrossRef]
- Hans, C.P.; Weisenburger, D.D.; Greiner, T.C.; Gascoyne, R.D.; Delabie, R.D.; Delabie, O.; Ott, G.; Müller-Hermelink, H.K.; Campo, E.; Braziel, R.M.; et al. Confirmation of the molecular classification of diffuse large B-cell lymphoma by immunohistochemistry using a tissue microarray. Blood 2004, 103, 275–282. [Google Scholar] [CrossRef]
- Voropaeva, E.N.; Voevoda, M.I.; Maksimov, V.N.; Orlov, Y.L.; Pospelova, T.I.; Seregina, O.B. Clinical aspects of TP53 gene inactivation in diffuse large B-cell lymphoma. BMC Med Genom. 2019, 12 (Suppl 2), 35. [Google Scholar] [CrossRef] [Green Version]
- Piletič, K.; Kunej, T. MicroRNA epigenetic signatures in human disease. Arch. Toxicol. 2016, 90, 2405–2419. [Google Scholar] [CrossRef]
- Strmsek, Z.; Kunej, T. MicroRNA Silencing by DNA Methylation in Human Cancer: A Literature Analysis. Non-Coding RNA 2015, 1, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Voropaeva, E.N.; Voevoda, M.I.; Maksimov, V.N.; Pospelova, T.I. Prognostic impact of the TP53 rs1625895 polymorphism in DLBCL patients. Br. J. Haematol. 2015, 169, 32–35. [Google Scholar] [CrossRef] [PubMed]
- Voropaeva, E.N.; Voevoda, M.I.; Maksimov, V.N.; Pospelova, T.I. Frequency, spectrum, and functional significance of TP53 mutations in patients with diffuse large B-cell lymphoma. Mol. Biol. 2017, 51, 53–60. (In Russian) [Google Scholar] [CrossRef]
- Munch-Petersen, H.D.; Asmar, F.; Dimopoulos, K.; Areškevičiūtė, A.; Brown, P.; Girkov, M.S.; Pedersen, A.; Sjö, L.D.; Heegaard, S.; Broholm, H.; et al. TP53 hotspot mutations are predictive of survival in primary central nervous system lymphoma patients treated with combination chemotherapy. Acta Neuropathol. Commun. 2016, 22, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorofchian, S.; El-Achi, H.; Yan, Y.; Esquenazi, Y.; Ballester, L.Y. Characterization of genomic alterations in primary central nervous system lymphomas. J. Neuro-Oncol. 2018, 140, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Ryabchicov, D.A.; Dudina, I.A.; Vorotnikov, I.K.; Talipov, O.A.; Titov, K.S.; Denchik, D.A.; Kazubskaya, T.P.; Burdennyy, A.M.; Loginov, V.I. The role of microRNA genes methylation in different molecular-biological subtypes of breast cancer. Malig. Tumours 2018, 8, 5–11. (In Russian) [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Lai, G.; Lin, Q.; Issah, M.A.; Fu, H.; Shen, J. A miR-129-5P/ARID3A Negative Feedback Loop Modulates Diffuse Large B Cell Lymphoma Progression and Immune Evasion Through Regulating the PD-1/PD-L1 Checkpoint. Front. Cell Dev. Biol. 2021, 9, 735855. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative Analysis of Complex Cancer Genomics and Clinical Profiles Using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]


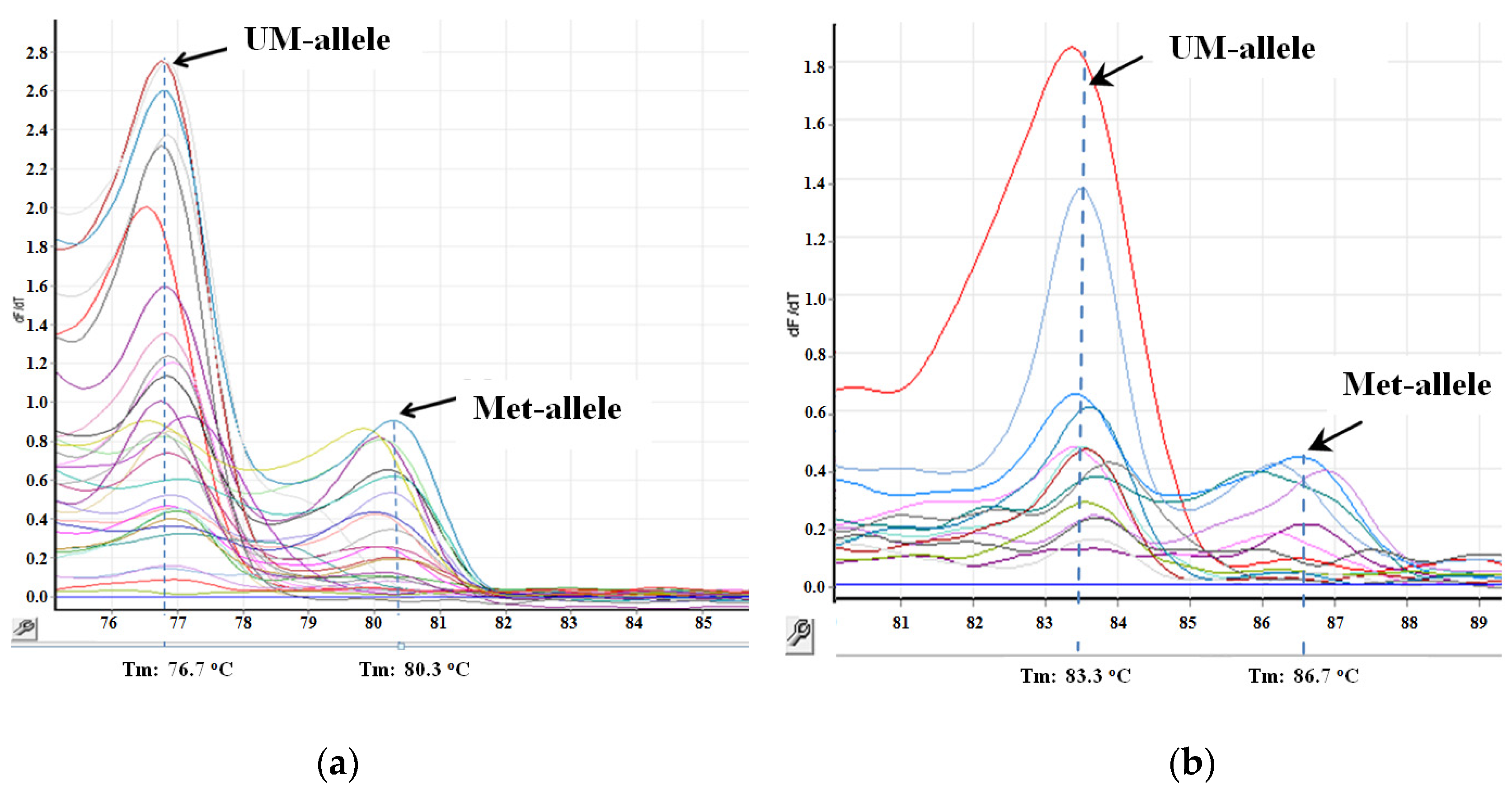



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | MIR-34A | MIR-34B/C | MIR-203 | MIR-129-2 | ||||
|---|---|---|---|---|---|---|---|---|
| M | U | M | U | M | U | M | U | |
| Tumor (n = 73) | 20 | 53 | 45 | 28 | 48 | 25 | 49 | 24 |
| Control (n = 11) | 0 | 11 | 0 | 11 | 0 | 11 | 0 | 11 |
| p-value | 0.047 | <0.001 | <0.001 | <0.001 | ||||
| Gene A | Gene B | p-Value | q-Value | Tendency |
|---|---|---|---|---|
| MIR-203 | MIR-129-2 | 0.003 | 0.018 | Co-occurrence |
| MIR-34B/C | MIR-34A | 0.010 | 0.029 | Co-occurrence |
| MIR-34B/C | MIR-129-2 | 0.014 | 0.029 | Co-occurrence |
| MIR-34B/C | MIR-203 | 0.024 | 0.036 | Co-occurrence |
| MIR-34A | MIR-203 | 0.207 | 0.345 | Co-occurrence |
| MIR-34A | MIR-129-2 | 0.422 | 0.545 | Mutual exclusivity |
| Clinical Parameters | All Group (n = 73) | MIR-34A | MIR-34B/C | MIR-203 | MIR-129-2 | ||||
|---|---|---|---|---|---|---|---|---|---|
| M (n = 20) | U (n = 53) | M (n = 45) | U (n = 28) | M (n = 48) | UM (n = 25) | M (n = 49) | UM (n = 24) | ||
| Sex (p-value) | 0.100 | 0.698 | 0.101 | 0.562 | |||||
| M | 36 | 13 | 23 | 23 | 13 | 27 | 9 | 23 | 13 |
| F | 37 | 7 | 30 | 22 | 15 | 21 | 16 | 26 | 11 |
| Age (p-value) | 0.051 | 0.241 | 0.900 | 0.950 | |||||
| ≤60 years | 46 | 9 | 37 | 26 | 20 | 30 | 16 | 31 | 15 |
| >60 years | 27 | 11 | 16 | 19 | 8 | 18 | 9 | 18 | 9 |
| ECOG score (p-value) | 0.758 | 0.034 | 0.617 | 0.457 | |||||
| ≤1 | 35 | 9 | 26 | 26 | 9 | 22 | 13 | 22 | 13 |
| ≥2 | 38 | 11 | 27 | 19 | 19 | 26 | 12 | 27 | 11 |
| Stage (p-value) | 0.513 | 0.068 | 0.095 | 0.105 | |||||
| I | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 |
| II | 4 | 1 | 3 | 4 | 0 | 4 | 0 | 4 | 0 |
| III | 11 | 2 | 9 | 9 | 2 | 9 | 2 | 8 | 3 |
| IV | 57 | 16 | 41 | 31 | 26 | 34 | 23 | 36 | 21 |
| Extranodal involvement (p-value) | 0.279 | 0.509 | 0.898 | 0.703 | |||||
| ≥2 sites | 14 | 5 | 9 | 9 | 5 | 9 | 5 | 10 | 4 |
| Bone marrow involvement (p-value) | 0.832 | 0.785 | 0.492 | 0.156 | |||||
| Yes | 17 | 5 | 12 | 10 | 7 | 10 | 7 | 9 | 8 |
| LDH level (p-value) | 0.064 | 0.305 | 0.202 | 0.270 | |||||
| >450 IU/L | 42 | 15 | 27 | 28 | 14 | 23 | 19 | 26 | 16 |
| IPI score (p-value) | 0.002 | 0.667 | 0.639 | 0.435 | |||||
| 0–2 | 29 | 2 | 27 | 17 | 12 | 20 | 9 | 21 | 8 |
| 3–5 | 44 | 18 | 26 | 28 | 16 | 28 | 16 | 28 | 16 |
| Parameters | All Group | MIR-34B/C and MIR-203 | MIR-34A | MIR-34B/C | MIR-203 | MIR-129-2 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| MM | UU | M | U | M | U | M | U | M | U | ||
| IGH Ki67+ (p-value) | - | 0.004 | 0.887 | 0.026 | 0.011 | 0.109 | |||||
| >45% cells | 31/54 | 20/26 | 2/9 | 10/17 | 21/37 | 25/37 | 6/17 | 24/34 | 7/20 | 25/39 | 6/15 |
| DLBCL subtype (p-value) | - | - | 0.046 | 0.541 | 0.851 | 0.851 | |||||
| Non-GCB-like | 35/54 | - | - | 6/14 | 29/40 | 21/34 | 14/20 | 23/35 | 12/19 | 23/34 | 12/19 |
| GCB-like | 19/54 | - | - | 8/14 | 11/40 | 13/34 | 6/20 | 12/35 | 7/19 | 12/34 | 7/19 |
| Parameter | IPI Score | MIR-34A | MIR-34B/C | MIR-203 | MIR-129-2 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0–2 | 3–5 | p-Value | M | U | p-Value | M | U | p-Value | M | U | p-Value | M | U | p-Value | |
| Remission rate Abs (%) | 24/29 | 28/44 | 0.078 | 11/20 (55) | 41/53 (77.4) | 0.060 | 33/45 | 19/28 | 0.616 | 35/48 | 17/25 | 0.660 | 36/49 | 16/24 | 0.547 |
| OS (%) | 65.5 | 43.2 | 0.043 | 40.0 | 56.6 | 0.162 | 53.3 | 50.0 | 0.699 | 54.2 | 48.0 | 0.590 | 57.1 | 41.7 | 0.269 |
| Method | Gene | Primer Sequences | Product Size (bp) | Ta (°C) | Reference |
|---|---|---|---|---|---|
| MS-HRM | MIR-34A | F 5/-tttttttttaggtggaggagatgt-3/ R 5/-ccaaacaaacccaaacaaaac-3/ | 155 | 64 | [17] |
| MIR-34B/C | F 5/-ttgttattaaaataaggtatagtatta-3/ R 5/-cgcttctcaaacatcttctct-3/ | 99 | 56 | ||
| MS-PCR | MIR-203 | MF 5/-gagtattttcggtttagacgagac-3/ MR 5/-ccttttatacgacgcaaccg-3/ UMF 5/-tttgagtatttttggtttagatgagat-3/ UMR 5/-aacaccttttatacaacacaacca-3/ | 287 | 60 | [22] |
| MIR-129-2 | MF 5/-gagttgggggatcgcggac-3/ MR 5/-atataccgacttcttcgattcgccg-3/ | 189 | 62 | [16] | |
| UMF 5/-gagttgggggattgtggat-3/ UMR 5/-aatataccaacttcttcaattcacca-3/ | 188 | 60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voropaeva, E.N.; Pospelova, T.I.; Orlov, Y.L.; Churkina, M.I.; Berezina, O.V.; Gurazheva, A.A.; Ageeva, T.A.; Seregina, O.B.; Maksimov, V.N. The Methylation of the p53 Targets the Genes MIR-203, MIR-129-2, MIR-34A and MIR-34B/C in the Tumor Tissue of Diffuse Large B-Cell Lymphoma. Genes 2022, 13, 1401. https://doi.org/10.3390/genes13081401
Voropaeva EN, Pospelova TI, Orlov YL, Churkina MI, Berezina OV, Gurazheva AA, Ageeva TA, Seregina OB, Maksimov VN. The Methylation of the p53 Targets the Genes MIR-203, MIR-129-2, MIR-34A and MIR-34B/C in the Tumor Tissue of Diffuse Large B-Cell Lymphoma. Genes. 2022; 13(8):1401. https://doi.org/10.3390/genes13081401
Chicago/Turabian StyleVoropaeva, Elena N., Tatjana I. Pospelova, Yuriy L. Orlov, Maria I. Churkina, Olga V. Berezina, Anna A. Gurazheva, Tatjana A. Ageeva, Olga B. Seregina, and Vladimir N. Maksimov. 2022. "The Methylation of the p53 Targets the Genes MIR-203, MIR-129-2, MIR-34A and MIR-34B/C in the Tumor Tissue of Diffuse Large B-Cell Lymphoma" Genes 13, no. 8: 1401. https://doi.org/10.3390/genes13081401