

An Attempt to Identify the Medaka Receptor for Somatolactin Alpha Using a Reverse Genetics Approach


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
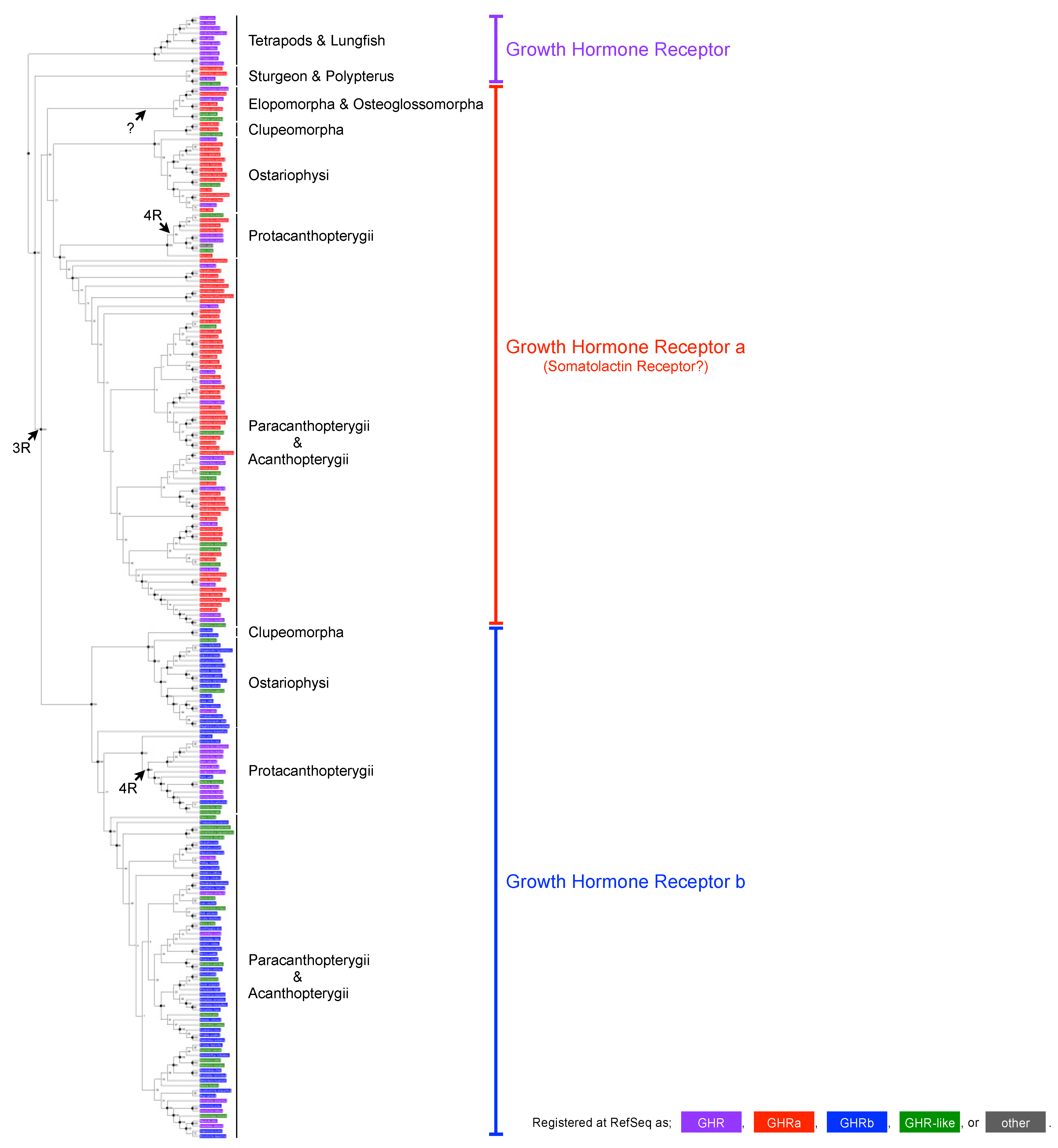
2.2. Phylogenetic Tree Reconstruction
2.3. Preparation and Microinjection of the gRNA and Cas9 mRNA
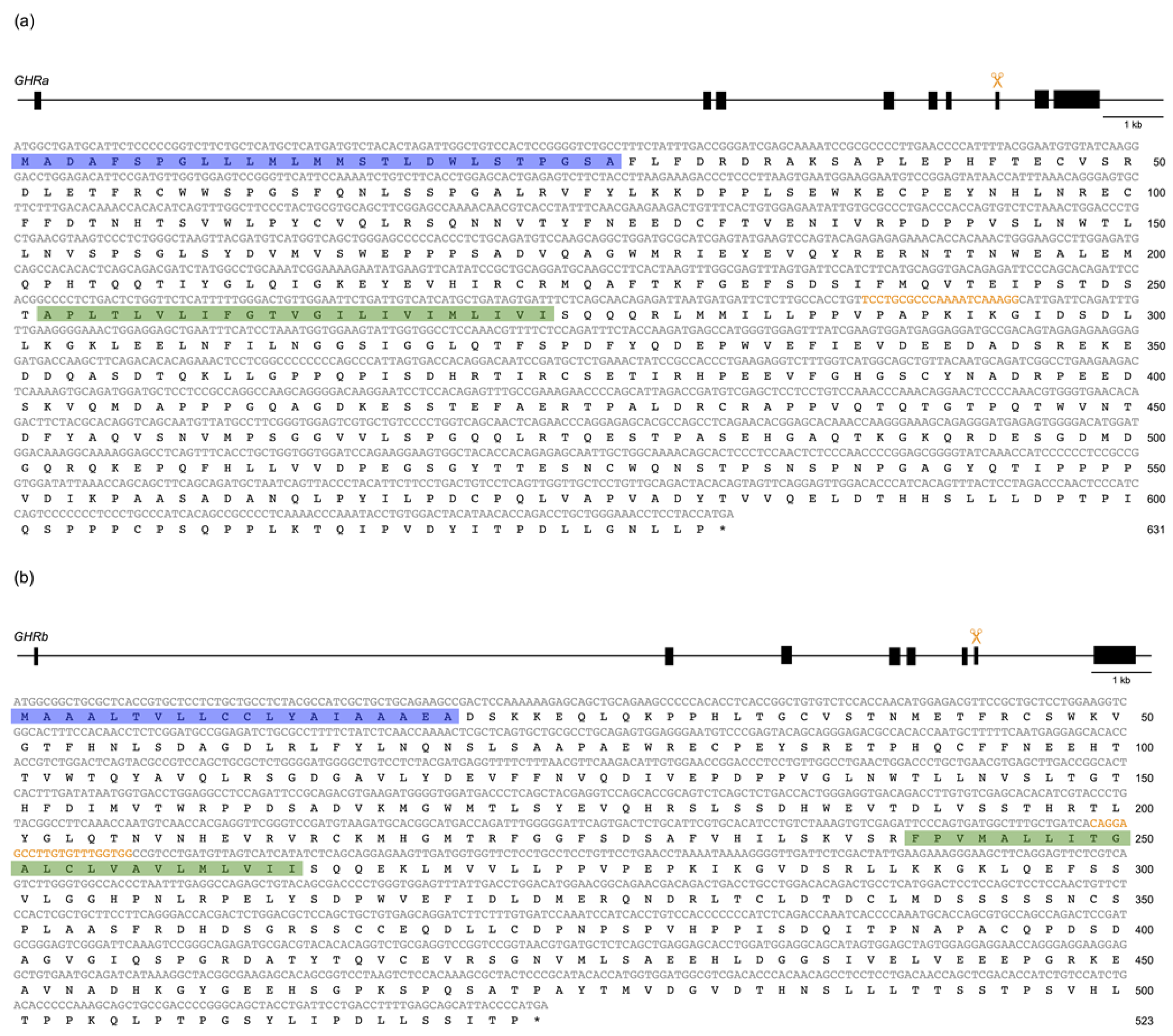
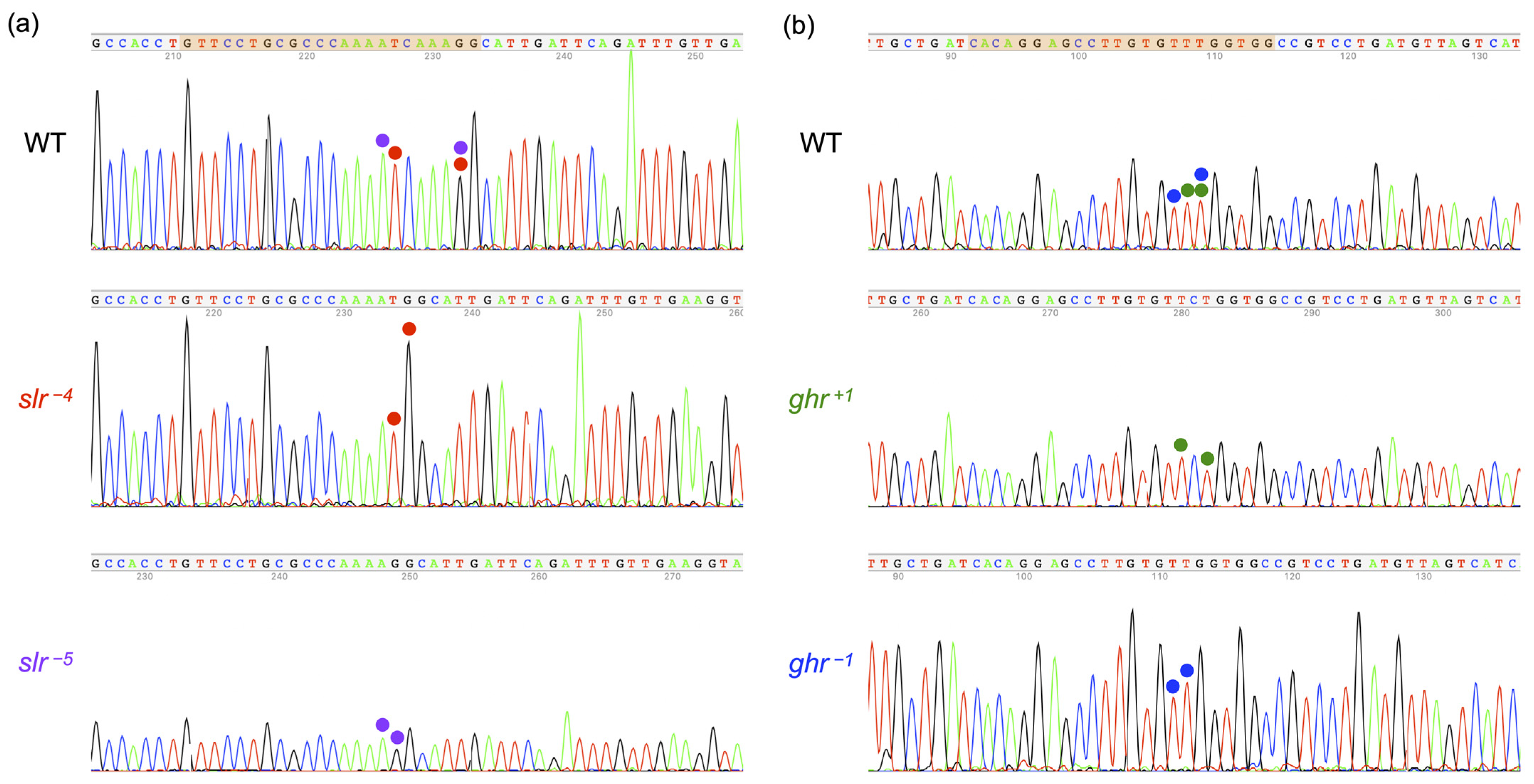
2.4. Identification of ins/del and Frameshift Mutations in SLR and GHR
2.5. Analysis of Viability
2.6. Analysis of Body Color and Body Length
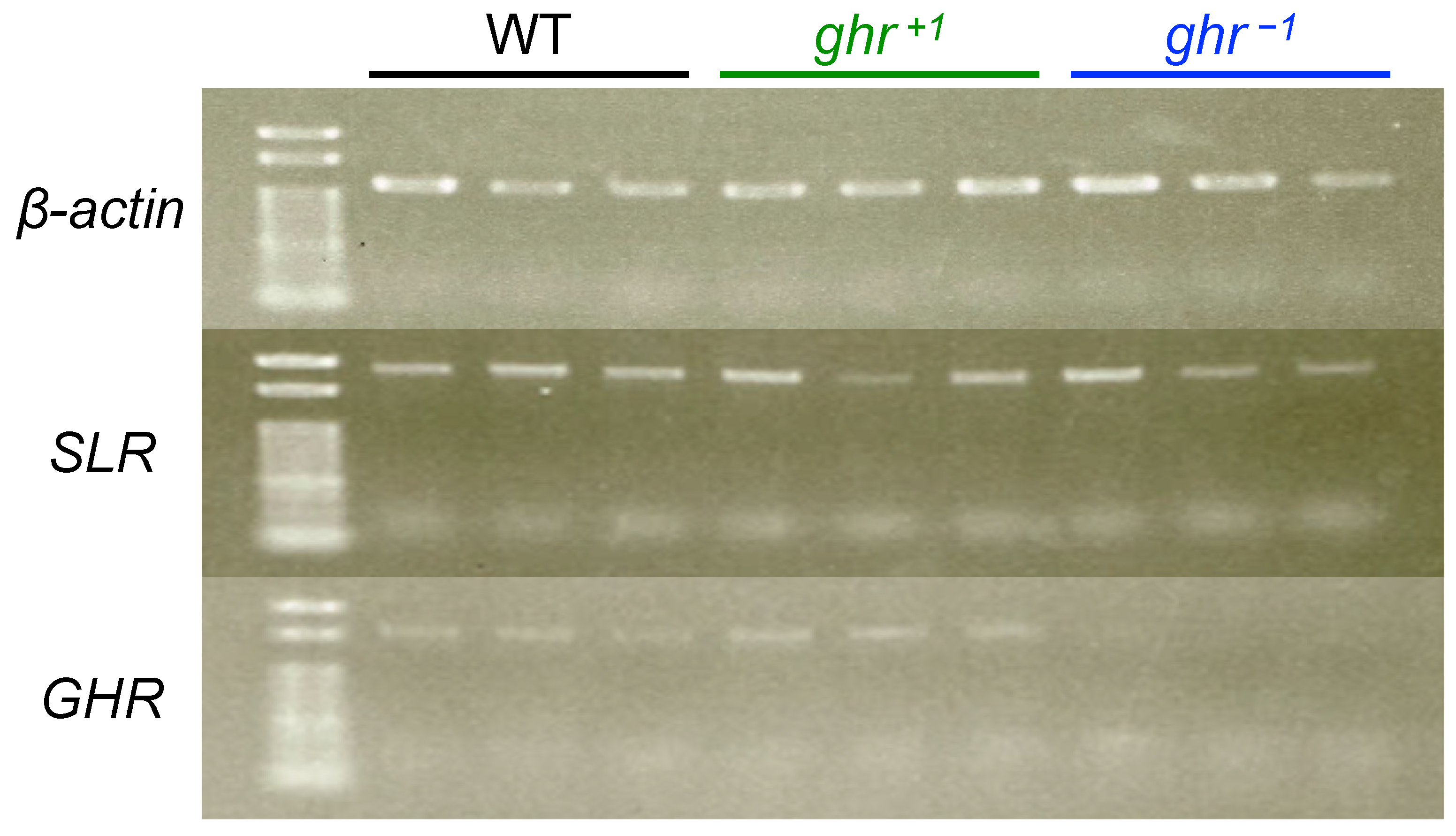
2.7. Reverse Transcription Polymerase Chain Reaction
2.8. Statistics
3. Results
3.1. Phylogenetic Reassessment of GHRa and GHRb in Teleosts
3.2. Mutagenesis Efficiency
3.3. Confirmation of Frameshift Mutations in slr and ghr Mutants
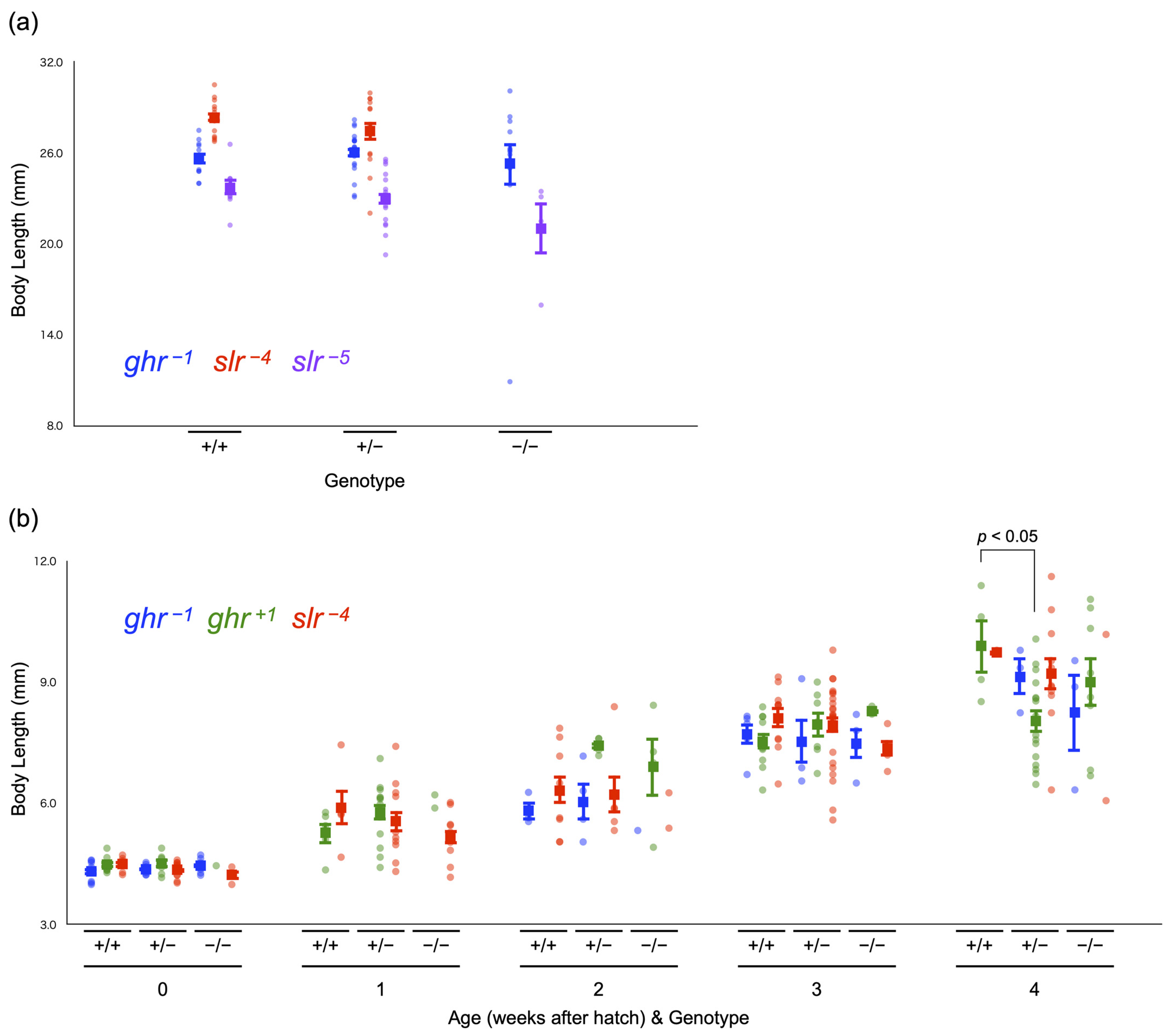
3.4. Viability and Body Size of Full Siblings Obtained by Crossing Heterozygous Mutants
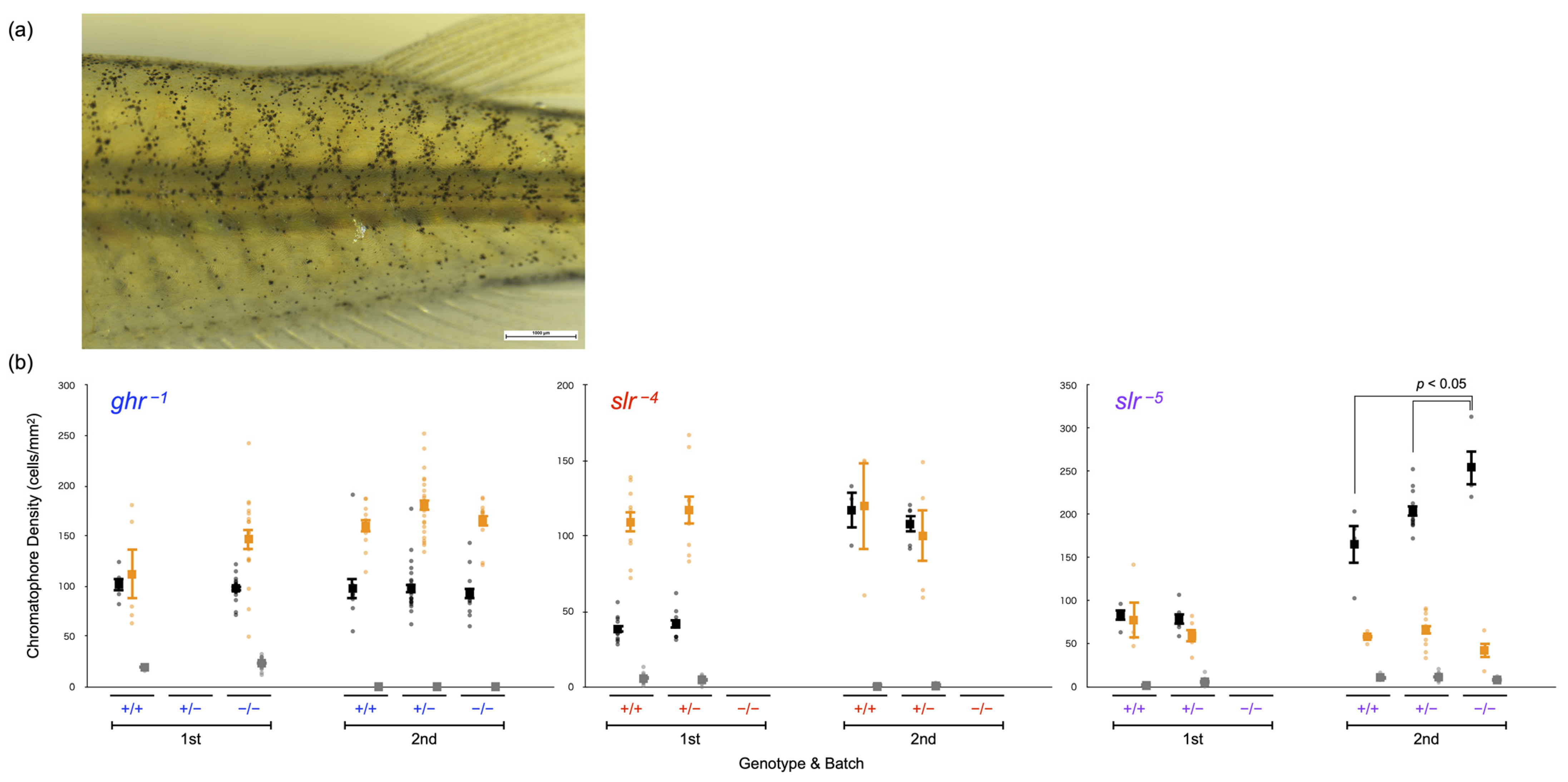
3.5. Body Color of Full Siblings Obtained by Crossing Heterozygous Mutant Fish
4. Discussion
4.1. Confirmation of Frameshift Mutations in ghr/slr Mutants
4.2. Estimation of Medaka SLR Functions
4.3. Estimation of Medaka GHR Functions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ono, M.; Takayama, Y.; Rand-Weaver, M.; Sakata, S.; Yasunaga, T.; Noso, T.; Kawauchi, H. cDNA cloning of somatolactin, a pituitary protein related to growth hormone and prolactin. Proc. Natl. Acad. Sci. USA 1990, 87, 4330–4334. [Google Scholar] [CrossRef] [Green Version]
- Fukamachi, S.; Sugimoto, M.; Mitani, H.; Shima, A. Somatolactin selectively regulates proliferation and morphogenesis of neural-crest derived pigment cells in medaka. Proc. Natl. Acad. Sci. USA 2004, 101, 10661–10666. [Google Scholar] [CrossRef] [Green Version]
- Fukamachi, S.; Yada, T.; Meyer, A.; Kinoshita, M. Effects of constitutive expression of somatolactin alpha on skin pigmentation in medaka. Gene 2009, 442, 81–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasano, Y.; Yoshimura, A.; Fukamachi, S. Reassessment of the function of somatolactin alpha in lipid metabolism using medaka mutant and transgenic strains. BMC Genet. 2012, 13, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimoto, M.; Uchida, N.; Hatayama, M. Apoptosis in skin pigment cells of the medaka, Oryzias latipes (Teleostei), during long-term chromatic adaptation: The role of sympathetic innervation. Cell Tissue Res. 2000, 301, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M. Morphological color changes in fish: Regulation of pigment cell density and morphology. Microsc. Res. Tech. 2002, 58, 496–503. [Google Scholar] [CrossRef]
- Reinecke, M.; Björnsson, B.T.; Dickhoff, W.W.; McCormick, S.D.; Navarro, I.; Power, D.M.; Gutiérrez, J. Growth hormone and insulin-like growth factors in fish: Where we are and where to go. Gen. Comp. Endocrinol. 2005, 142, 20–24. [Google Scholar] [CrossRef]
- Komine, R.; Nishimaki, T.; Kimura, T.; Oota, H.; Naruse, K.; Homma, N.; Fukamachi, S. Transgenic medaka that overexpress growth hormone have a skin color that does not indicate the activation or inhibition of somatolactin-a signal. Gene 2016, 584, 38–46. [Google Scholar] [CrossRef]
- Fukada, H.; Ozaki, Y.; Pierce, A.L.; Adachi, S.; Yamauchi, K.; Hara, A.; Swanson, P.; Dickhoff, W.W. Salmon growth hormone receptor: Molecular cloning, ligand specificity, and response to fasting. Gen. Comp. Endocrinol. 2004, 139, 61–71. [Google Scholar] [CrossRef]
- Fukada, H.; Ozaki, Y.; Pierce, A.L.; Adachi, S.; Yamauchi, K.; Hara, A.; Swanson, P.; Dickhoff, W.W. Identification of the salmon somatolactin receptor, a new member of the cytokine receptor family. Endocrinology 2005, 146, 2354–2361. [Google Scholar] [CrossRef]
- Chen, M.; Huang, X.; Yuen, D.S.; Cheng, C.H. A study on the functional interaction between the GH/PRL family of polypeptides with their receptors in zebrafish: Evidence against GHRI being the receptor for somatolactin. Mol. Cell. Endocrinol. 2011, 337, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, Y.; Fukada, H.; Kazero, Y.; Adachi, S.; Hara, A.; Yamauchi, K. Molecular cloning and characterization of growth hormone receptor and its homologue in the Japanese eel (Anguilla japonica). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2006, 143, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Ocampo Daza, D.; Larhammar, D. Evolution of the growth hormone, prolactin, prolactin 2 and somatolactin family. Gen. Comp. Endocrinol. 2018, 264, 94–112. [Google Scholar] [CrossRef]
- Ocampo Daza, D.; Larhammar, D. Evolution of the receptors for growth hormone, prolactin, erythropoietin and thrombopoietin in relation to the vertebrate tetraploidizations. Gen. Comp. Endocrinol. 2018, 257, 143–160. [Google Scholar] [CrossRef] [PubMed]
- Hwang, W.Y.; Fu, Y.; Reyon, D.; Maeder, M.L.; Tsai, S.Q.; Sander, J.D.; Peterson, R.T.; Yeh, J.R.; Joung, J.K. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat. Biotechnol. 2013, 3, 227–229. [Google Scholar] [CrossRef] [Green Version]
- Ansai, S.; Kinoshita, M. Targeted mutagenesis using CRISPR/Cas system in medaka. Biol. Open. 2014, 3, 362–371. [Google Scholar] [CrossRef] [Green Version]
- Lander, E.S. The heroes of CRISPR. Cell 2016, 164, 18–28. [Google Scholar] [CrossRef] [Green Version]
- Homma, N.; Harada, Y.; Uchikawa, T.; Kamei, Y.; Fukamachi, S. Protanopia (red color-blindness) in medaka: A simple system for producing color-blind fish and testing their spectral sensitivity. BMC Genet. 2017, 18, 10. [Google Scholar] [CrossRef] [Green Version]
- Kanazawa, N.; Goto, M.; Harada, Y.; Takimoto, C.; Sasaki, Y.; Uchikawa, T.; Kamei, Y.; Matsuo, M.; Fukamachi, S. Changes in a cone opsin repertoire affect color-dependent social behavior in medaka but not behavioral photosensitivity. Front. Genet. 2020, 11, 801. [Google Scholar] [CrossRef]
- Supek, F.; Lehner, B.; Lindeboom, R.G.H. To NMD or not to NMD: Nonsense-mediated mRNA decay in cancer and other genetic diseases. Trends Genet. 2021, 37, 657–668. [Google Scholar] [CrossRef]
- Delgadin, T.H.; Castañeda-Cortés, D.C.; Sacks, C.; Breccia, A.; Fernandino, J.I.; Vissio, P.G. Morphological colour adaptation during development in fish: Involvement of growth hormone receptor 1. J. Exp. Biol. 2020, 223, jeb230375. [Google Scholar] [CrossRef] [PubMed]
- Godowski, P.J.; Leung, D.W.; Meacham, L.R.; Galgani, J.P.; Hellmiss, R.; Keret, R.; Rotwein, P.S.; Parks, J.S.; Laron, Z.; Wood, W.I. Characterization of the human growth hormone receptor gene and demonstration of a partial gene deletion in two patients with Laron-type dwarfism. Proc. Natl. Acad. Sci. USA 1989, 20, 8083–8087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laron, Z. Laron syndrome (primary growth hormone resistance or insensitivity): The personal experience 1958–2003. J. Clin. Endocrinol. Metab. 2004, 89, 1031–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, D.; Li, F.; Li, Q.; Li, J.; Zhao, Y.; Hu, X.; Zhang, R.; Li, N. Generation of a miniature pig disease model for human Laron syndrome. Sci. Rep. 2015, 5, 15603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Xu, B.C.; Maheshwari, H.G.; He, L.; Reed, M.; Lozykowski, M.; Okada, S.; Cataldo, L.; Coschigamo, K.; Wagner, T.E.; et al. A mammalian model for Laron syndrome produced by targeted disruption of the mouse growth hormone receptor/binding protein gene (the Laron mouse). Proc. Natl. Acad. Sci. USA 1997, 94, 13215–13220. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| gRNA | Target Sequence (5′→3′) | Eggs Injected (N) | Survivors on the Day Following Injection (N) | Survivors on the Day Following Injection (%) | Normal Development (N) | Larvae Subjected to HMA (N) | Larvae Confirmed to Have a Band Shift (N) | Mutagenesis Efficiency (%) |
|---|---|---|---|---|---|---|---|---|
| SLR5 | CCAGAGTCAGAGGGGCCGTGG | 40 | 16 | 40.0 | 15 | 15 | 0 | 0 |
| SLR6 | TCTCATTTTTGGGACTGTTGG | ― | ― | ― | ― | ― | ― | ― |
| SLR7 | TTTGGGCGCAGGAACAGGTGG | ― | ― | ― | ― | ― | ― | ― |
| SLR8 | TCCTGCGCCCAAAATCAAAGG | 29 | 15 | 51.7 | 14 | 14 | 5 | 35.7 |
| GHR5 | GATCAGCAAAGCCATCACTGG | 143 | 35 | 24.5 | 11 | 11 | 0 | 0 |
| GHR6 | CAGGAGCCTTGTGTTTGGTGG | 127 | 48 | 37.8 | 17 | 16 | 4 | 25.0 |
| GHR7 | GGTTCAGGAACAGGAGGCAGG | 35 | 20 | 57.1 | 19 | 13 | 0 | 0 |
| GHR8 | TCCTGAACCTAAAATAAAAGG | 22 | 15 | 68.2 | 15 | 12 | 0 | 0 |
| gRNA | Potential Off-Target Sequence (5′→3′) | Chromosome |
|---|---|---|
| GHR6 | CAGGAGCCTTGTGTTTGGTGG | 12 (target) |
| CAGGAGACTTGTGTTTGTGGG | 1 | |
| CAGGAGCCATGTTTTTGGGGG | 12 | |
| GAGCAGCCTTGTGTTTGAGGG | 19 |
| Target Gene | G0 Individual Number | Mutation Type | Mutation | Sequence (5′→3′) | Number of F1 Larvae with Mutations (%) | Number of F1 Adults with Mutations (%) |
|---|---|---|---|---|---|---|
| SLR | - | WT | - | GCCCAAAATCAAAGGCATTGATTCAGATTTGTTGAAG | - | - |
| 1 | six-deletion | SLR-6 (1) | GCCCAAAATCA––––––TTGATTCAGATTTGTTGAAG | 3 (18.8) | 13 (13.1) | |
| 2 | one-deletion | SLR+3-4 | GCCCAAAA–CAGAGGCATTGATTCAGATTTGTTGAAG | 1 (6.3) | 0 (0) | |
| 4 × 7 | nine-insertion | SLR+9 | GCCCAAAAATCAATGAATCAAAGGCATTGATTCAGATTTGTTGAAG | 2 (12.5) | 0 (0) | |
| 4 × 7, 6 | five-deletion | SLR-5 | GCCCAAAA–––––GGCATTGATTCAGATTTGTTGAAG | 4 (18.8, 6.3) | 5 (3.9) | |
| 4 × 7, 8 | six-deletion | SLR-6 (2) | GCCCAAA––––––GGCATTGATTCAGATTTGTTGAAG | 2 (12.5) | 3 (1.7) | |
| 5, 6 | two-insertion | SLR+2 (1) | GCCCAAACAATCAAAGGCATTGATTCAGATTTGTTGAAG | 1 (6.3) | 1 (0.8) | |
| 6 | sixteen-insertion | SLR+17-1 | GCCCAAAAGAGCCTGAAACGATGTCCAAAGGCATTGATTCAGATTTGTTGAAG | 0 (0) | 3 (2.4) | |
| 6 | four-deletion | SLR-4 | GCCCAAAAT––––GGCATTGATTCAGATTTGTTGAAG | 1 (6.3) | 9 (7) | |
| 6 | three-deletion | SLR-3 | GCCCAAAA–––AAGGCATTGATTCAGATTTGTTGAAG | 0 (0) | 3 (2.3) | |
| 6 | two-insertion | SLR+2 (2) | GCCCAAAATGCCAAAGGCATTGATTCAGATTTGTTGAAG | 1 (6.3) | 9 (7) | |
| 6 | two-deletion | SLR-2 | GCCCAAAA––AAAGGCATTGATTCAGATTTGTTGAAG | 0 (0) | 2 (1.6) | |
| 8 | seven-deletion | SLR+6-13 | GCCCAAAGGTCTT–––––––ATTCAGATTTGTTGAAG | 0 (0) | 1 (0.6) | |
| 8 | six-insertion | SLR+7-1 | GCCCAAAAATGGGCCCAAAGGCATTGATTCAGATTTGTTGAAG | 2 (12.5) | 4 (2.3) | |
| 8 | one-insertion | SLR+1 | GCCCAAAATTCAAAGGCATTGATTCAGATTTGTTGAAG | 2 (12.5) | 29 (16.5) | |
| Total | 19 | 82 | ||||
| GHR | - | WT | - | GCCTTGTGTTTGGTGGCCATCCTGATGTTAGTCATCA | - | - |
| 1 | one-insertion | GHR+1 | GCCTTGTGTTCTGGTGGCCATCCTGATGTTAGTCATCA | 1 (6.3) | 4 (4) | |
| 4 × 7, 8 | one-deletion | GHR-1 | GCCTTGTGTT–GGTGGCCATCCTGATGTTAGTCATCA | 3 (12.5, 6.3) | 5 (2.8) | |
| 6 | six-insertion | GHR+6 | GCCTTGTGTGGTGCCTTGGTGGCCATCCTGATGTTAGTCATCA | 1 (6.3) | 7 (5.5) | |
| 8 | two-deletion | GHR-2 | GCCTTGTGT––GGTGGCCATCCTGATGTTAGTCATCA | 0 (0) | 2 (1.1) | |
| Total | 5 | 18 |
| Strain | Genotypes | ||
|---|---|---|---|
| +/+ | +/− | −/− | |
| slr–4 | 75 | 114 | 0 |
| 0.000 | |||
| slr–5 | 18 | 33 | 4 |
| 0.009 | |||
| ghr+1 | 12 | 24 | 9 |
| 0.741 | |||
| ghr–1 | 32 | 68 | 28 |
| 0.687 | |||
| Strain | Age (weeks) and Genotype | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | |||||||||||
| +/+ | +/− | −/− | +/+ | +/− | −/− | +/+ | +/− | −/− | +/+ | +/− | −/− | +/+ | +/− | −/− | |
| ghr+1 | 7 | 7 | 1 | 5 | 12 | 2 | 0 | 4 | 4 | 10 | 7 | 3 | 4 | 16 | 8 |
| 0.088 | 0.323 | 0.135 | 0.035 | 0.424 | |||||||||||
| ghr–1 | 7 | 7 | 7 | 0 | 0 | 0 | 3 | 4 | 1 | 5 | 4 | 4 | 0 | 3 | 3 |
| 0.311 | ― | 0.607 | 0.354 | 0.223 | |||||||||||
| slr–4 | 10 | 18 | 9 | 8 | 18 | 15 | 9 | 6 | 2 | 15 | 27 | 5 | 4 | 11 | 2 |
| 0.960 | 0.223 | 0.027 | 0.071 | 0.379 | |||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moroki, Y.; Komori, M.; Ogawa, Y.; Nagumo, E.; Ohno, H.; Fukamachi, S. An Attempt to Identify the Medaka Receptor for Somatolactin Alpha Using a Reverse Genetics Approach. Genes 2023, 14, 796. https://doi.org/10.3390/genes14040796
Moroki Y, Komori M, Ogawa Y, Nagumo E, Ohno H, Fukamachi S. An Attempt to Identify the Medaka Receptor for Somatolactin Alpha Using a Reverse Genetics Approach. Genes. 2023; 14(4):796. https://doi.org/10.3390/genes14040796
Chicago/Turabian StyleMoroki, Yuko, Mamiko Komori, Yuko Ogawa, Erina Nagumo, Haruna Ohno, and Shoji Fukamachi. 2023. "An Attempt to Identify the Medaka Receptor for Somatolactin Alpha Using a Reverse Genetics Approach" Genes 14, no. 4: 796. https://doi.org/10.3390/genes14040796
APA StyleMoroki, Y., Komori, M., Ogawa, Y., Nagumo, E., Ohno, H., & Fukamachi, S. (2023). An Attempt to Identify the Medaka Receptor for Somatolactin Alpha Using a Reverse Genetics Approach. Genes, 14(4), 796. https://doi.org/10.3390/genes14040796