
Genome-Wide Identification, and In-Silico Expression Analysis of YABBY Gene Family in Response to Biotic and Abiotic Stresses in Potato (Solanum tuberosum)

 , , , , , , and
, , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Database Search and Retrieval of Sequences
2.2. Determination of Physio-Chemical Properties of YABBY Proteins
2.3. Gene Structure Analysis
2.4. Comparative Phylogenetic Analysis
2.5. Gene Duplication and Synteny Analysis
2.6. Gene Ontology Enrichment Analysis
2.7. Expression Analysis
2.8. MiRNA Analysis
3. Results
3.1. Identification of the YABBY Genes in Potato
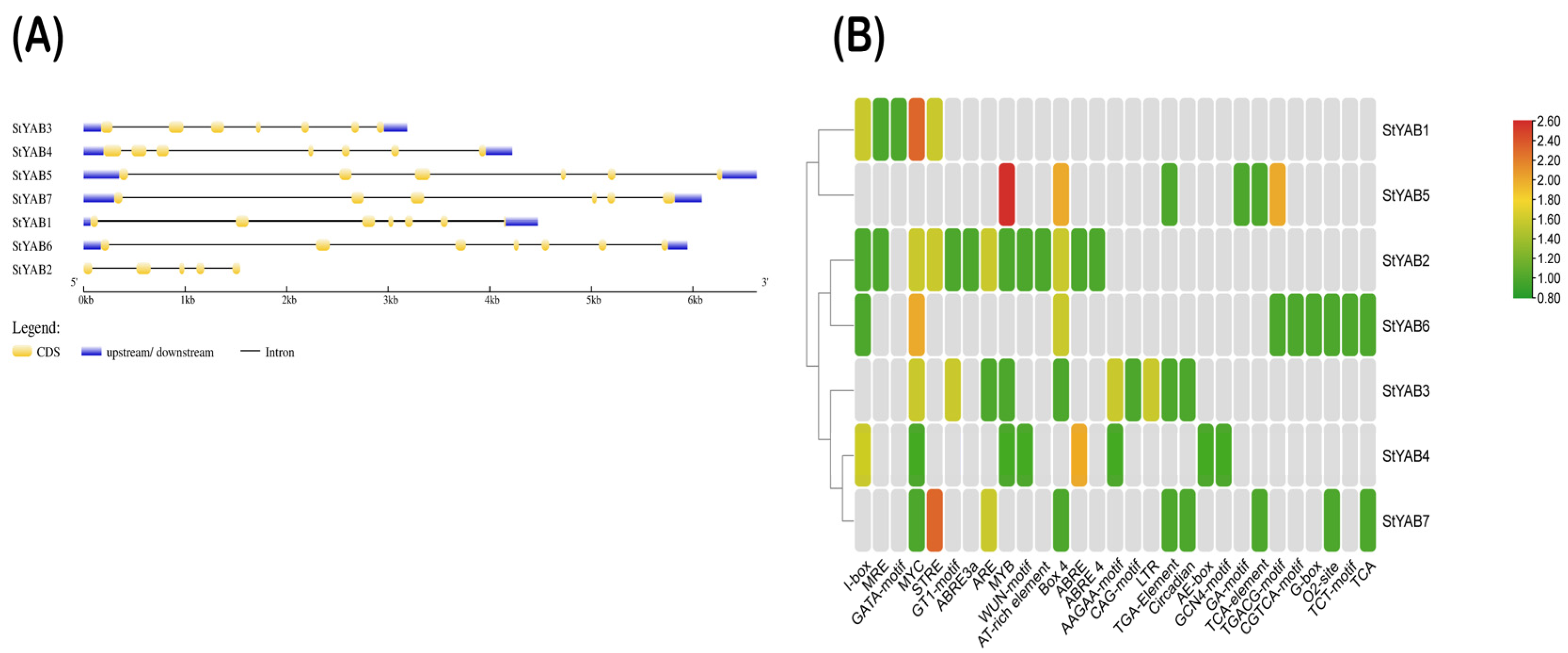
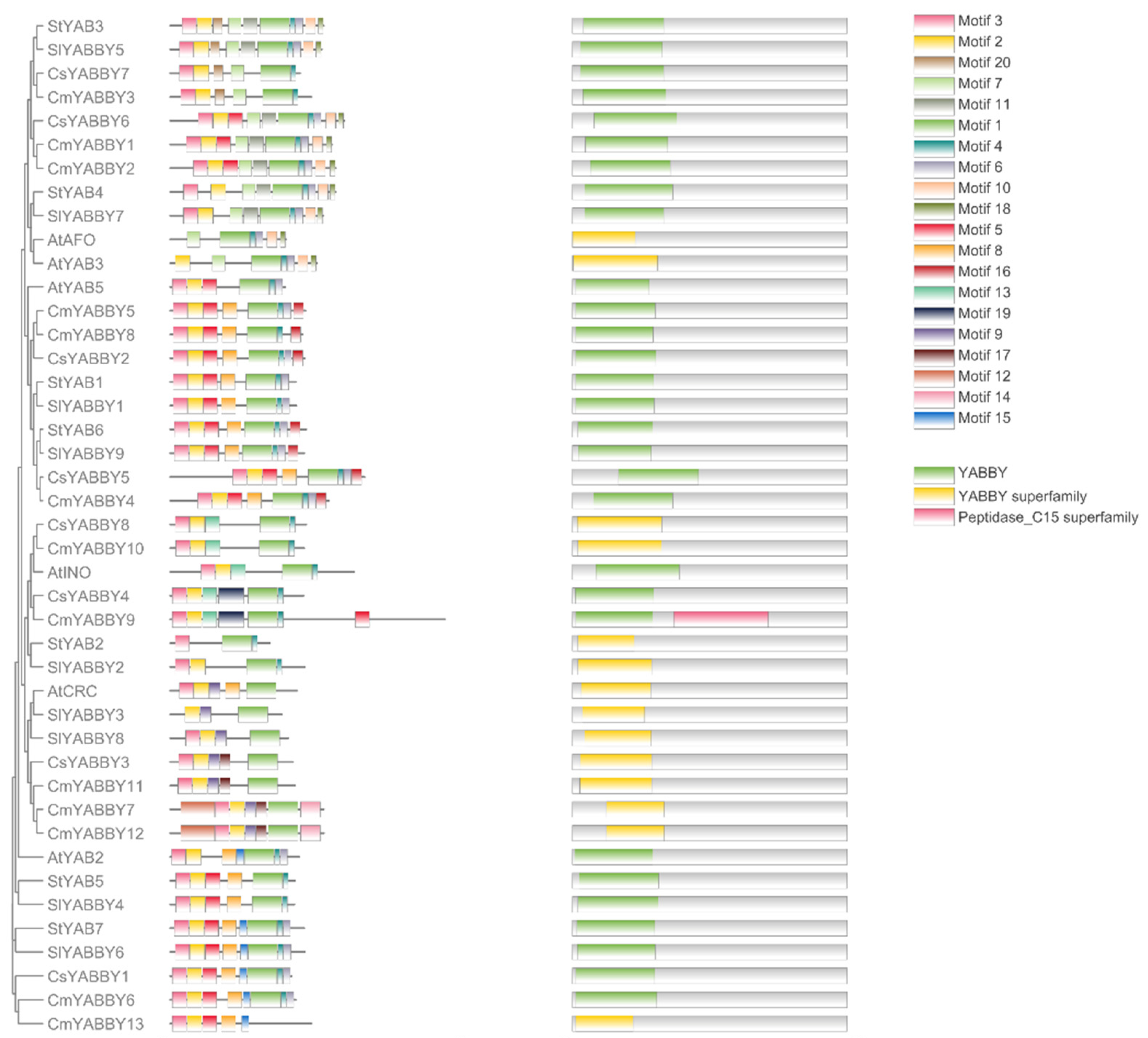
3.2. Gene Structures and Recognition of Conserved Cis-Elements and Motifs
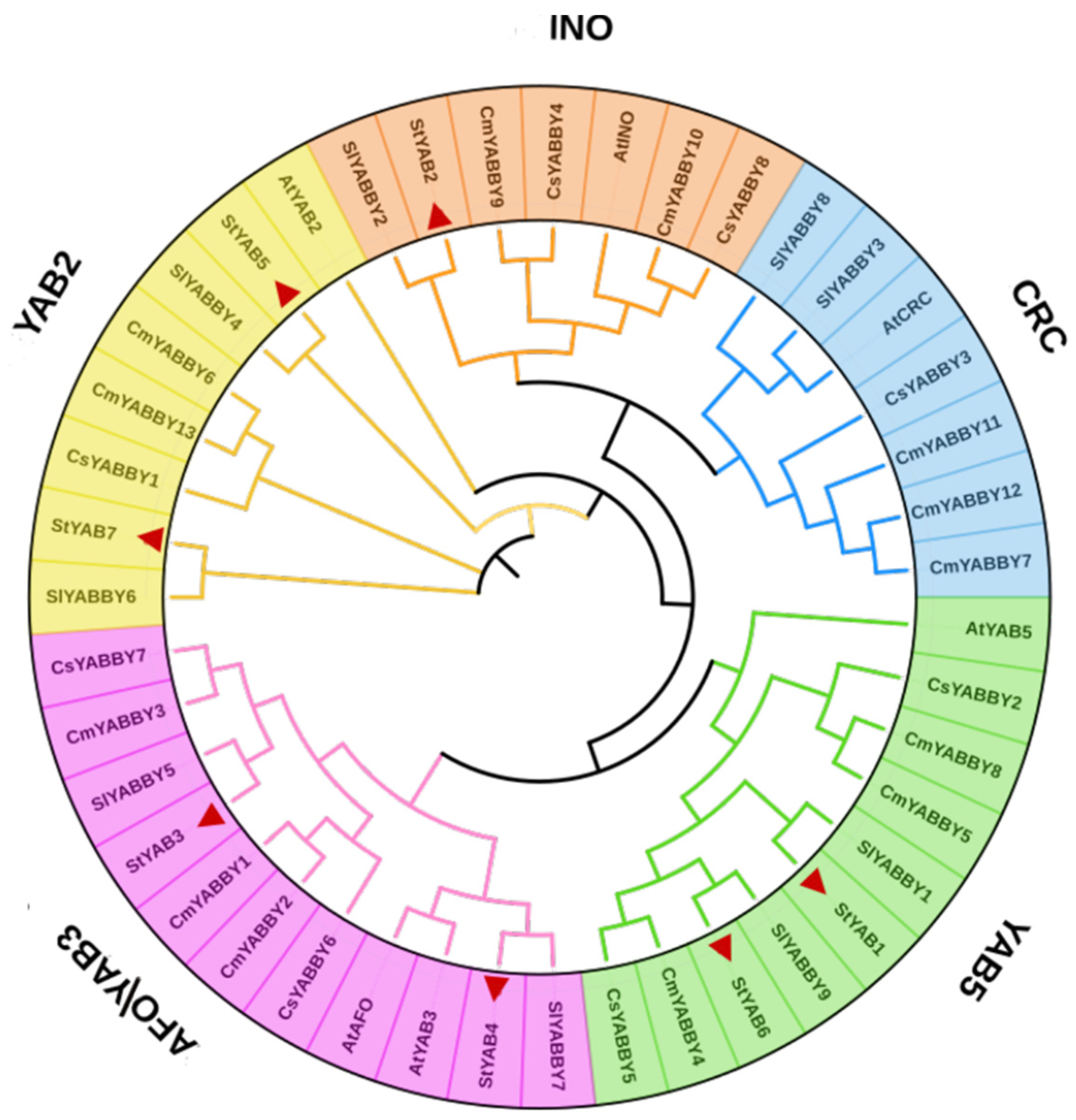
3.3. Phylogenetic Analysis of the YABBY Gene Family
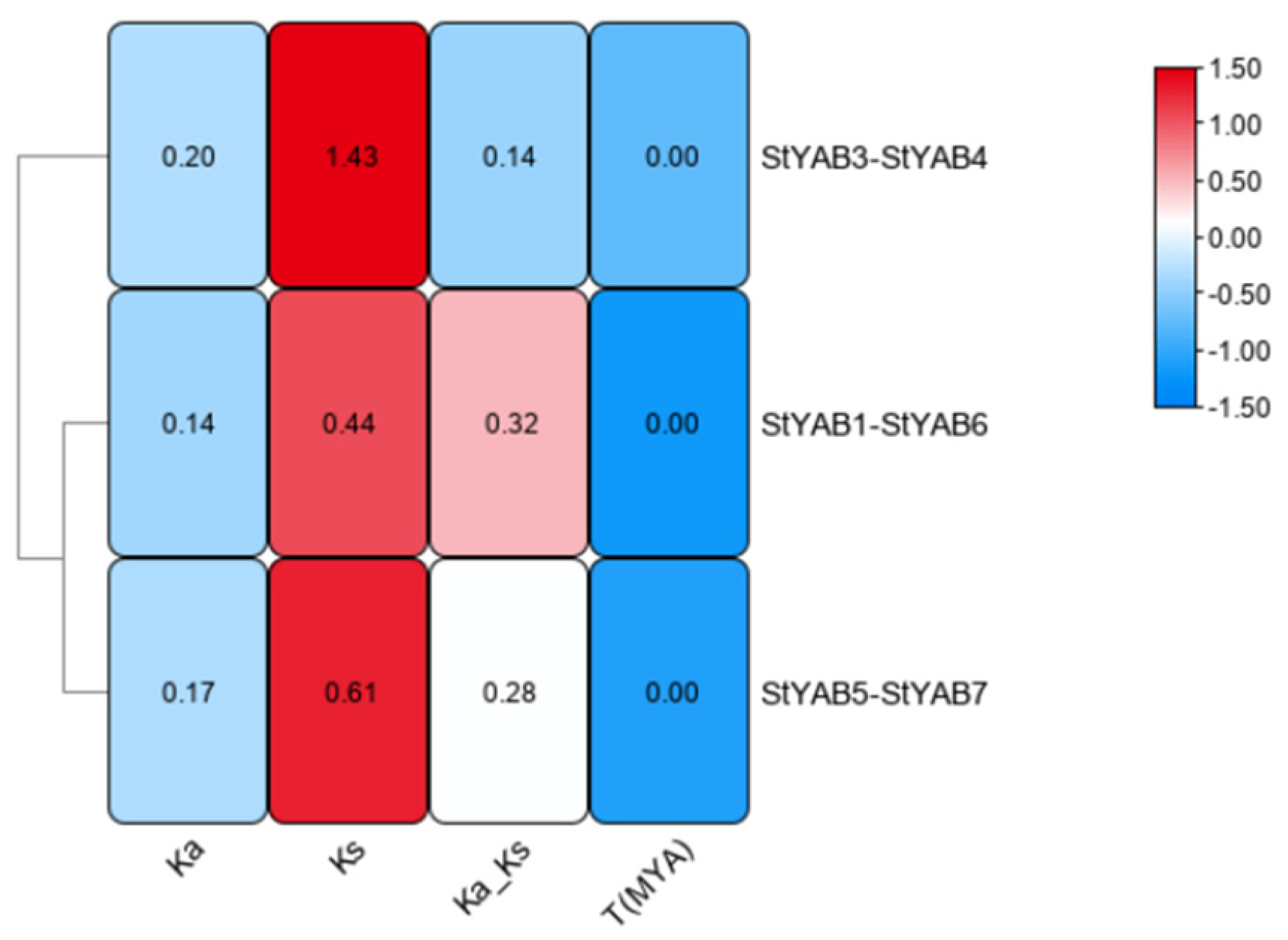
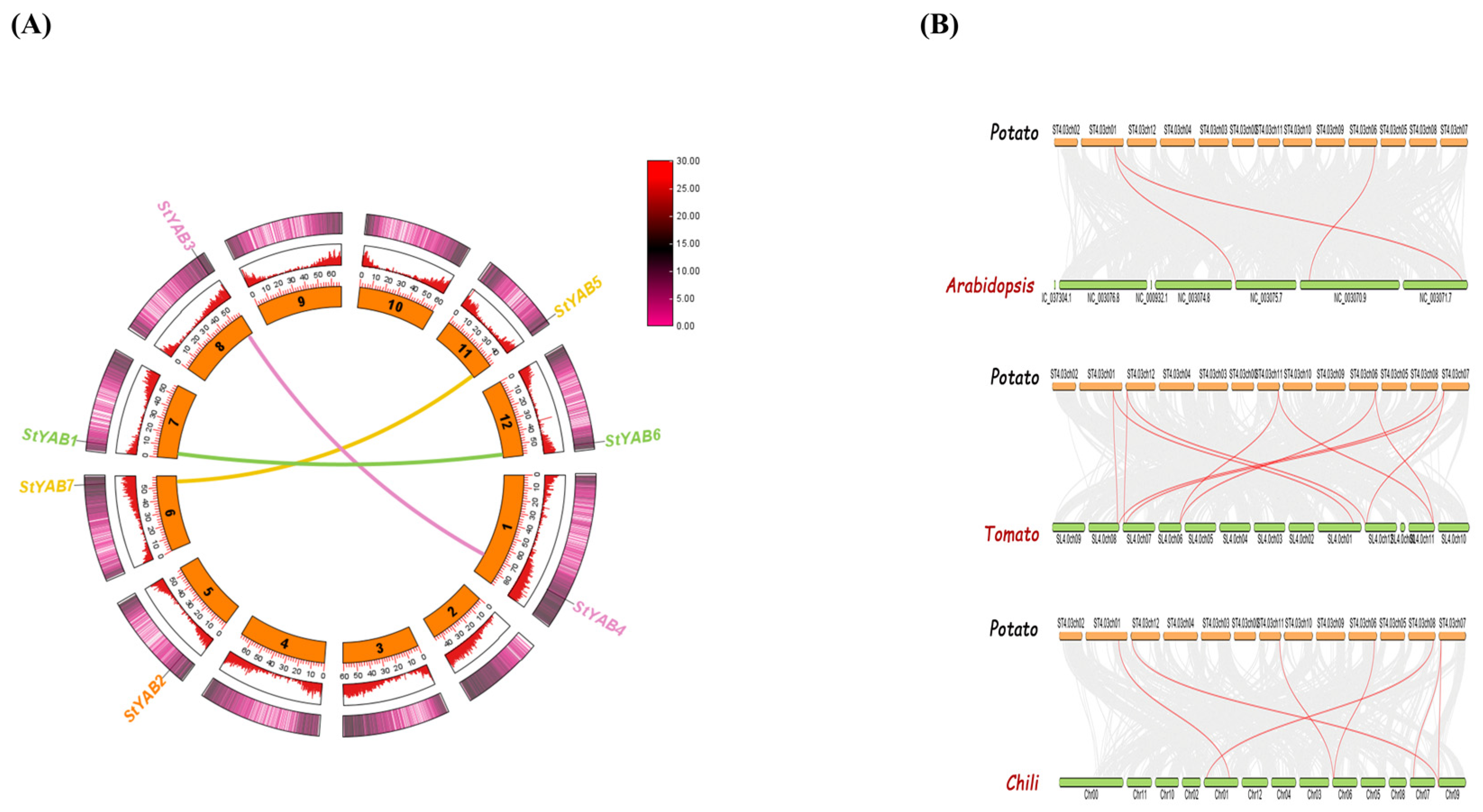
3.4. StYAB Gene Location on Chromosomes and Duplication Assessment
3.5. Enrichment Analysis and Ortholog Identification
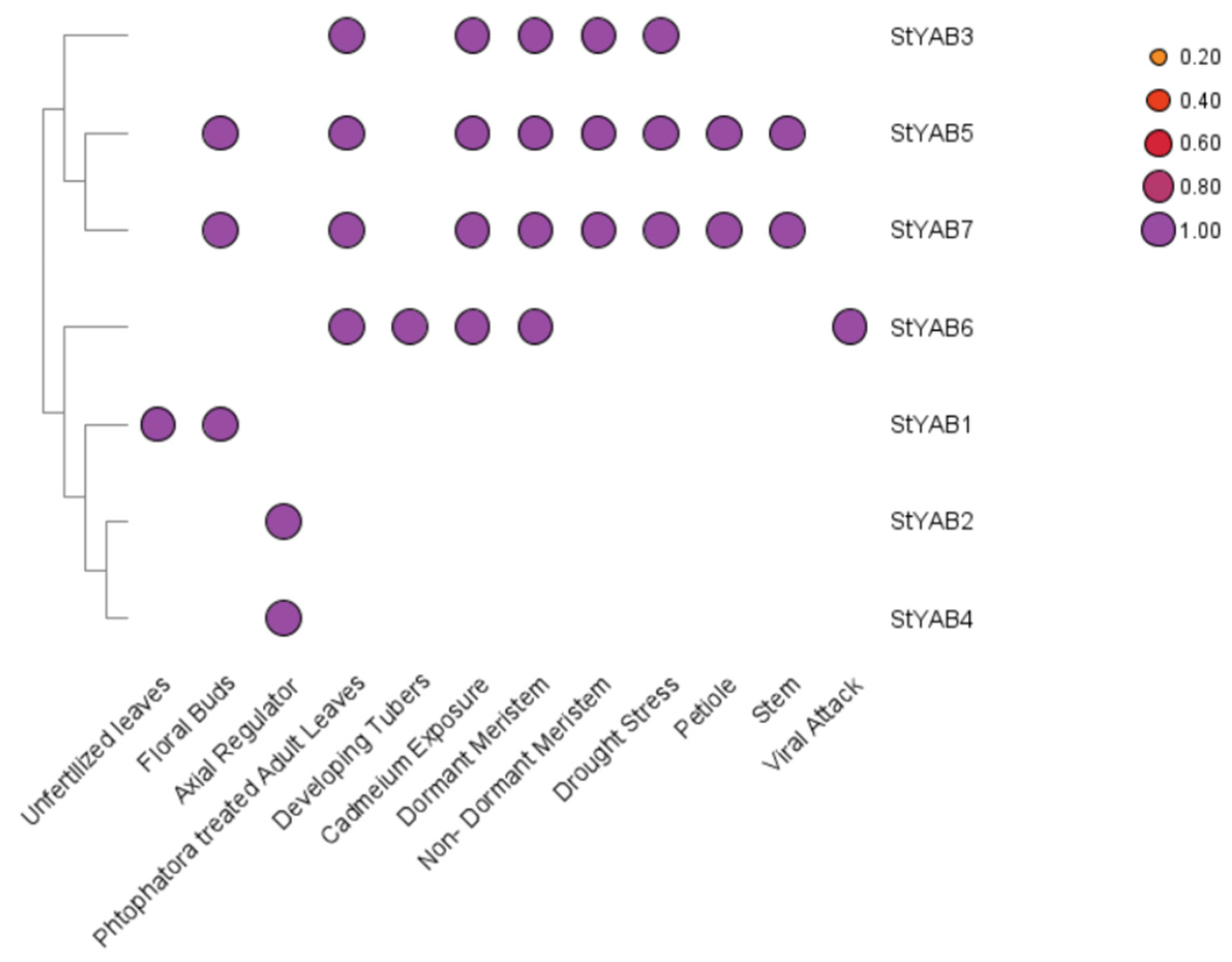
3.6. Expression Analysis of StYAB Gene
3.7. MiRNA Targets in POTATO
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hussain, Q.; Asim, M.; Zhang, R.; Khan, R.; Farooq, S.; Wu, J. Transcription Factors Interact with ABA through Gene Expression and Signaling Pathways to Mitigate Drought and Salinity Stress. Biomolecules 2021, 11, 1159. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Huang, B.; Huang, Z.; Zhang, Z. Genome-wide identification and analysis of the YABBY gene family in Moso Bamboo (Phyllostachys edulis (Carriere) J. Houz). PeerJ 2021, 9, e11780. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Gong, Q.; Wang, L.; Jin, Y.; Xi, J.; Li, Z.; Qin, W.; Yang, Z.; Lu, L.; Chen, Q.; et al. Genome-Wide Study of YABBY Genes in Upland Cotton and Their Expression Patterns under Different Stresses. Front. Genet. 2018, 9, 33. [Google Scholar] [CrossRef]
- Hussain, Q.; Zheng, M.; Ashraf, M.F.; Khan, R.; Yasir, M.; Farooq, S.; Zhang, R.; Wu, J. Genome-wide identification, characterization and expression analysis of the ABA receptor PYL gene family in response to ABA, photoperiod, and chilling in vegetative buds of Liriodendron chinense. Sci. Hortic. 2022, 303, 111200. [Google Scholar] [CrossRef]
- Eckardt, N.A. YABBY Genes and the Development and Origin of Seed Plant Leaves. Plant Cell 2010, 22, 2103. [Google Scholar] [CrossRef]
- Li, Z.; Li, G.; Cai, M.; Priyadarshani, S.V.G.N.; Aslam, M.; Zhou, Q.; Huang, X.; Wang, X.; Liu, Y.; Qin, Y. Genome-Wide Analysis of the YABBY Transcription Factor Family in Pineapple and Functional Identification of AcYABBY4 Involvement in Salt Stress. Int. J. Mol. Sci. 2019, 20, 5863. [Google Scholar] [CrossRef] [PubMed]
- Sarojam, R.; Sappl, P.G.; Goldshmidt, A.; Efroni, I.; Floyd, S.K.; Eshed, Y.; Bowman, J.L. Differentiating Arabidopsis Shoots from Leaves by Combined YABBY Activities. Plant Cell 2010, 22, 2113–2130. [Google Scholar] [CrossRef]
- Stahle, M.I.; Kuehlich, J.; Staron, L.; von Arnim, A.G.; Golz, J.F. YABBYs and the Transcriptional Corepressors LEUNIG and LEUNIG_HOMOLOG Maintain Leaf Polarity and Meristem Activity in Arabidopsis. Plant Cell 2009, 21, 3105–3118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-L.; Yang, Z.-P.; Zhang, J.; Zhang, L.-G. Ectopic Expression of BraYAB1-702, a Member of YABBY Gene Family in Chinese Cabbage, Causes Leaf Curling, Inhibition of Development of Shoot Apical Meristem and Flowering Stage Delaying in Arabidopsis thaliana. Int. J. Mol. Sci. 2013, 14, 14872–14891. [Google Scholar] [CrossRef]
- Lin, Z.; Li, X.; Shannon, L.M.; Yeh, C.-T.; Wang, M.L.; Bai, G.; Peng, Z.; Li, J.; Trick, H.N.; Clemente, T.E.; et al. Parallel domestication of the Shattering1 genes in cereals. Nat. Genet. 2012, 44, 720–724. [Google Scholar] [CrossRef]
- Zhao, S.P.; Lu, D.; Yu, T.F.; Ji, Y.J.; Zheng, W.J.; Zhang, S.X.; Chai, S.C.; Chen, Z.Y.; Cui, X.Y. Genome-wide analysis of the YABBY family in soybean and functional identification of GmYABBY10 involvement in high salt and drought stresses. Plant Physiol. Biochem. 2017, 119, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, L.; Sun, X.; Li, Y.; Yao, J.; van Nocker, S.; Wang, X. Genome-Wide Analysis of the YABBY Gene Family in Grapevine and Functional Characterization of VvYABBY4. Front. Plant Sci. 2019, 10, 1207. [Google Scholar] [CrossRef] [PubMed]
- Bartley, G.E.; Ishida, B.K. Ethylene-sensitive and insensitive regulation of transcription factor expression during in vitro tomato sepal ripening. J. Exp. Bot. 2007, 58, 2043–2051. [Google Scholar] [CrossRef]
- Peng, X.; Wu, Q.; Teng, L.; Tang, F.; Pi, Z.; Shen, S. Transcriptional regulation of the paper mulberry under cold stress as revealed by a comprehensive analysis of transcription factors. BMC Plant Biol. 2015, 15, 108. [Google Scholar] [CrossRef]
- Yuan, J.; Shen, C.; Xin, J.; Li, Z.; Li, X.; Zhou, J. Genome-wide Characterization and Expression Analysis of YABBY Gene Family in Three Cultivars of Cucurbita Linn. And Their Response of Salt Stress in Cucurbita Moschata. 2020. [Google Scholar] [CrossRef]
- Hou, H.; Wu, P.; Gao, L.; Zhang, C.; Hou, X. Characterization and expression profile analysis of YABBY family genes in Pak-choi (Brassica rapa ssp. chinensis) under abiotic stresses and hormone treatments. Plant Growth Regul. 2019, 87, 421–432. [Google Scholar] [CrossRef]
- Xia, J.; Wang, D.; Peng, Y.; Wang, W.; Wang, Q.; Xu, Y.; Li, T.; Zhang, K.; Li, J.; Xu, X. Genome-Wide Analysis of the YABBY Transcription Factor Family in Rapeseed (Brassica napus L.). Genes 2021, 12, 981. [Google Scholar] [CrossRef]
- Buttar, Z.A.; Yang, Y.; Sharif, R.; Nan Wu, S.; Xie, Y.; Wang, C. Genome Wide Identification, Characterization, and Expression Analysis of YABBY-Gene Family in WHEAT (Triticum aestivum L.). Agronomy 2020, 10, 1189. [Google Scholar] [CrossRef]
- Inal, B.; Buyuk, I.; Ilhan, E.; Aras, S. Genome-wide analysis of Phaseolus vulgaris C2C2-YABBY transcription factors under salt stress conditions. 3 Biotech 2017, 7, 302. [Google Scholar] [CrossRef]
- Huang, Z.; Van Houten, J.; Gonzalez, G.; Xiao, H.; van der Knaap, E. Genome-wide identification, phylogeny and expression analysis of SUN, OFP and YABBY gene family in tomato. Mol. Genet. Genom. 2013, 288, 111–129. [Google Scholar] [CrossRef]
- Tanaka, W.; Toriba, T.; Hirano, H.Y. Three TOB1-related YABBY genes are required to maintain proper function of the spikelet and branch meristems in rice. New Phytol. 2017, 215, 825–839. [Google Scholar] [CrossRef]
- Toriba, T.; Harada, K.; Takamura, A.; Nakamura, H.; Ichikawa, H.; Suzaki, T.; Hirano, H.Y. Molecular characterization the YABBY gene family in Oryza sativa and expression analysis of OsYABBY1. Mol. Genet Genom. 2007, 277, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Zhi-hong, L.; Lei, W. Characteristics and Expression Analysis of Transcription Factor YABBY Family in Maize. J. Agric. Sci. Technol. 2015, 17, 32–41. [Google Scholar]
- Luo, K.; Zhang, D.; Zhai, Z.; Liu, X.; Zhou, J.; Zhang, B.; Li, D. LsaYAB7, A Homologous Gene of FILAMENTOUS FLOWER, Participating In The Regulation of AdaxialAbaxial Polarity of Leaves In Lettuce. 2021. [Google Scholar] [CrossRef]
- Juarez, M.T.; Twigg, R.W.; Timmermans, M.C. Specification of adaxial cell fate during maize leaf development. Development 2004, 131, 4533–4544. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-Y.; Hsiao, Y.-Y.; Chang, S.-B.; Zhang, D.; Lan, S.-R.; Liu, Z.-J.; Tsai, W.-C. Genome-Wide Identification of YABBY Genes in Orchidaceae and Their Expression Patterns in Phalaenopsis Orchid. Genes 2020, 11, 955. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Li, S.; Gao, Y.; Bartholomew, E.S.; Wang, R.; Yang, H.; Liu, C.; Chen, X.; Wang, Y.; Liu, X.; et al. Genome-Wide Identification of YABBY Gene Family in Cucurbitaceae and Expression Analysis in Cucumber (Cucumis sativus L.). Genes 2022, 13, 467. [Google Scholar] [CrossRef]
- Tanaka, W.; Toriba, T.; Ohmori, Y.; Yoshida, A.; Kawai, A.; Mayama-Tsuchida, T.; Ichikawa, H.; Mitsuda, N.; Ohme-Takagi, M.; Hirano, H.-Y.J.T.P.C. The YABBY gene TONGARI-BOUSHI1 is involved in lateral organ development and maintenance of meristem organization in the rice spikelet. Plant Cell 2012, 24, 80–95. [Google Scholar] [CrossRef]
- Zhang, G.-Q.; Liu, K.-W.; Li, Z.; Lohaus, R.; Hsiao, Y.-Y.; Niu, S.-C.; Wang, J.-Y.; Lin, Y.-C.; Xu, Q.; Chen, L.-J.J.N. The Apostasia genome and the evolution of orchids. Nature 2017, 549, 379–383. [Google Scholar] [CrossRef]
- Ohmori, Y.; Tanaka, W.; Kojima, M.; Sakakibara, H.; Hirano, H.Y. WUSCHEL-RELATED HOMEOBOX4 is involved in meristem maintenance and is negatively regulated by the CLE gene FCP1 in rice. Plant Cell 2013, 25, 229–241. [Google Scholar] [CrossRef]
- Borrego-Benjumea, A.; Carter, A.; Tucker, J.R.; Yao, Z.; Xu, W.; Badea, A. Genome-Wide Analysis of Gene Expression Provides New Insights into Waterlogging Responses in Barley (Hordeum vulgare L.). Plants 2020, 9, 240. [Google Scholar] [CrossRef]
- Korpan, Y.I.; Nazarenko, E.A.; Skryshevskaya, I.V.; Martelet, C.; Jaffrezic-Renault, N.; El’skaya, A.V. Potato glycoalkaloids: True safety or false sense of security? Trends Biotechnol. 2004, 22, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Mondal, M.; Hossain, M.; Rasul, M.; Uddin, S. Genetic diversity in potato (Solanum tuberosum L.). Bangladesh J. Bot. 2008, 36, 121–125. [Google Scholar] [CrossRef]
- Koch, M.; Naumann, M.; Pawelzik, E.; Gransee, A.; Thiel, H. The Importance of Nutrient Management for Potato Production Part I: Plant Nutrition and Yield. Potato Res. 2019, 63, 97–119. [Google Scholar] [CrossRef]
- Goffart, J.-P.; Haverkort, A.; Storey, M.; Haase, N.; Martin, M.; Lebrun, P.; Ryckmans, D.; Florins, D.; Demeulemeester, K.J.P.R. Potato Production in Northwestern Europe (Germany, France, the Netherlands, United Kingdom, Belgium): Characteristics, Issues, Challenges and Opportunities. Potato Res. 2022, 65, 503–547. [Google Scholar] [CrossRef]
- Nasir, M.W.; Toth, Z.J.A. Effect of drought stress on potato production: A review. Agronomy 2022, 12, 635. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Hou, M.; Liu, Y.; Pei, Y.; Ye, M.; Zhou, Y.; Huang, C.; Zhao, Y.; Ma, H. Genome-wide identification, characterization and expression analysis of the non-specific lipid transfer proteins in potato. BMC Genom. 2019, 20, 375. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Tu, Z.J.; Millett, B.P.; Bradeen, J.M. Insights into organ-specific pathogen defense responses in plants: RNA-seq analysis of potato tuber-Phytophthora infestans interactions. BMC Genom. 2013, 14, 340. [Google Scholar] [CrossRef]
- Petek, M.; Rotter, A.; Kogovsek, P.; Baebler, S.; Mithofer, A.; Gruden, K. Potato virus Y infection hinders potato defence response and renders plants more vulnerable to Colorado potato beetle attack. Mol. Ecol. 2014, 23, 5378–5391. [Google Scholar] [CrossRef]
- Mengist, M.F.; Byrne, S.L.; Griffin, D.; Milbourne, D. A note on the early transcriptional response in leaves and root of potato plants to cadmium exposure. Ir. J. Agric. Food Res. 2021, 60, 27–32. [Google Scholar] [CrossRef]
- Sprenger, H.; Kurowsky, C.; Horn, R.; Erban, A.; Seddig, S.; Rudack, K.; Fischer, A.; Walther, D.; Zuther, E.; Kohl, K.; et al. The drought response of potato reference cultivars with contrasting tolerance. Plant Cell Environ. 2016, 39, 2370–2389. [Google Scholar] [CrossRef] [PubMed]
- Campbell, M.; Suttle, J.; Douches, D.S.; Buell, C.R. Treatment of potato tubers with the synthetic cytokinin 1-(alpha-ethylbenzyl)-3-nitroguanidine results in rapid termination of endodormancy and induction of transcripts associated with cell proliferation and growth. Funct. Integr. Genom. 2014, 14, 789–799. [Google Scholar] [CrossRef]
- Lin, X.; Kaul, S.; Rounsley, S.; Shea, T.P.; Benito, M.I.; Town, C.D.; Fujii, C.Y.; Mason, T.; Bowman, C.L.; Barnstead, M.; et al. Sequence and analysis of chromosome 2 of the plant Arabidopsis thaliana. Nature 1999, 402, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-Q.; Sveshnikov, D.; Zebarth, B.J.; Tai, H.; De Koeyer, D.; Millard, P.; Haroon, M.; Singh, M. Detection of Nitrogen Sufficiency in Potato Plants Using Gene Expression Markers. Am. J. Potato Res. 2009, 87, 50–59. [Google Scholar] [CrossRef]
- Zapata, L.; Ding, J.; Willing, E.M.; Hartwig, B.; Bezdan, D.; Jiao, W.B.; Patel, V.; Velikkakam James, G.; Koornneef, M.; Ossowski, S.; et al. Chromosome-level assembly of Arabidopsis thaliana Ler reveals the extent of translocation and inversion polymorphisms. Proc. Natl. Acad. Sci. USA 2016, 113, E4052–E4060. [Google Scholar] [CrossRef]
- Theologis, A.; Ecker, J.R.; Palm, C.J.; Federspiel, N.A.; Kaul, S.; White, O.; Alonso, J.; Altafi, H.; Araujo, R.; Bowman, C.L.; et al. Sequence and analysis of chromosome 1 of the plant Arabidopsis thaliana. Nature 2000, 408, 816–820. [Google Scholar] [CrossRef]
- Li, X.Q.; Griffiths, R.; De Koeyer, D.; Rothwell, C.; Gustafson, V.; Regan, S.; Flinn, B. Functional genomic resources for potato. Can. J. Plant Sci. 2008, 88, 573–581. [Google Scholar] [CrossRef]
- Flinn, B.; Rothwell, C.; Griffiths, R.; Lague, M.; DeKoeyer, D.; Sardana, R.; Audy, P.; Goyer, C.; Li, X.Q.; Wang-Pruski, G.; et al. Potato expressed sequence tag generation and analysis using standard and unique cDNA libraries. Plant Mol. Biol. 2005, 59, 407–433. [Google Scholar] [CrossRef]
- Shahzad, R.; Harlina, P.W.; Ayaad, M.; Ewas, M.; Nishawy, E.; Fahad, S.; Subthain, H.; Amar, M.H. Dynamic roles of microRNAs in nutrient acquisition and plant adaptation under nutrient stress: A review. Plant Omics 2018, 11, 58–79. [Google Scholar] [CrossRef]
- Tiwari, J.K.; Buckseth, T.; Zinta, R.; Saraswati, A.; Singh, R.K.; Rawat, S.; Chakrabarti, S.K. Genome-wide identification and characterization of microRNAs by small RNA sequencing for low nitrogen stress in potato. PLoS ONE 2020, 15, e0233076. [Google Scholar] [CrossRef] [PubMed]
- Sharif, y.; Zaynab, M.; Khan, K.A.; Li, S. Identification and Expression Analysis of DUF4228 Domain Containing (DDP) Genes in Potato Under Abiotic and Phytohormone Stress. Mol. Biol. Rep. 2021. [Google Scholar] [CrossRef]
- Iqbal, M.S.; Hafeez, M.N.; Wattoo, J.I.; Ali, A.; Sharif, M.N.; Rashid, B.; Tabassum, B.; Nasir, I.A. Prediction of Host-Derived miRNAs with the Potential to Target PVY in Potato Plants. Front. Genet. 2016, 7, 159. [Google Scholar] [CrossRef]
- Hou, X.; Du, Y.; Liu, X.; Zhang, H.; Liu, Y.; Yan, N.; Zhang, Z. Genome-Wide Analysis of Long Non-Coding RNAs in Potato and Their Potential Role in Tuber Sprouting Process. Int. J. Mol. Sci. 2017, 19, 101. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Lashbrook, C.C.; Cho, S.K.; Butler, N.M.; Sharma, P.; Muppirala, U.; Severin, A.J.; Hannapel, D.J. Transcriptional analysis of phloem-associated cells of potato. BMC Genom. 2015, 16, 665. [Google Scholar] [CrossRef]
- Lu, Y.; Feng, Z.; Liu, X.; Bian, L.; Xie, H.; Zhang, C.; Mysore, K.S.; Liang, J. MiR393 and miR390 synergistically regulate lateral root growth in rice under different conditions. BMC Plant Biol. 2018, 18, 261. [Google Scholar] [CrossRef]
- Hussain, Q.; Zheng, M.; Chang, W.; Ashraf, M.F.; Khan, R.; Asim, M.; Riaz, M.W.; Alwahibi, M.S.; Elshikh, M.S.; Zhang, R.; et al. Genome-Wide Identification and Expression Analysis of SnRK2 Gene Family in Dormant Vegetative Buds of Liriodendron chinense in Response to Abscisic Acid, Chilling, and Photoperiod. Genes 2022, 13, 1305. [Google Scholar] [CrossRef]
- Riaz, M.W.; Lu, J.; Shah, L.; Yang, L.; Chen, C.; Mei, X.D.; Xue, L.; Manzoor, M.A.; Abdullah, M.; Rehman, S.; et al. Expansion and Molecular Characterization of AP2/ERF Gene Family in Wheat (Triticum aestivum L.). Front. Genet. 2021, 12, 632155. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| YABBY Gene Name | Accession Number | Chromosome Number | Chromosome Location (bp) | Direction | No. of Amino Acids | Molecular Weight (kD) | Pi Value | No. of Introns | No. of Exons | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Phytozome V13 (6.1) | PLAZA Genome Database | Start | End | ||||||||
| StYAB 1 | Soltu.DM.07G003610 | PGSC0003DMT400079392 | 7 | 4307036 | 4311510 | F | 179 | 20.16703 | 8.77 | 6 | 7 |
| StYAB 2 | Soltu.DM.05G008980 | PGSC0003DMT400072893 | 5 | 9003101 | 9004644 | F | 142 | 16.15737 | 8.25 | 4 | 5 |
| StYAB 3 | Soltu.DM.08G025380 | PGSC0003DMT400012083 | 8 | 55208205 | 55211393 | R | 219 | 24.48169 | 7.22 | 6 | 7 |
| StYAB 4 | Soltu.DM.01G031660 | PGSC0003DMT400066793 | 1 | 71511821 | 71516043 | R | 218 | 24.16953 | 7.23 | 6 | 7 |
| StYAB 5 | Soltu.DM.11G024670 | PGSC0003DMT400007731 | 11 | 44742927 | 44749556 | R | 178 | 20.14184 | 9.24 | 5 | 6 |
| StYAB 6 | Soltu.DM.12G026680 | PGSC0003DMT400007474 | 12 | 56595139 | 56601086 | R | 194 | 22.09788 | 8.17 | 6 | 7 |
| StYAB 7 | Soltu.DM.06G029310 | PGSC0003DMT400015197 | 6 | 54473907 | 54479995 | F | 191 | 21.272 | 8.79 | 5 | 6 |
| Gene ID | GO Function | Sub-Cellular Localization | Gene Expression | Ortholog in Arabidopsis | References | ||
|---|---|---|---|---|---|---|---|
| Molecular Function | Biological Process | Gene Name | Function | ||||
| StYAB1 | Abaxial cell fate specification | Leaf development | Extra chromosomal | Unfertilized leaves and floral buds | YABBY 5 | Axial regulator | [45,46] |
| StYAB2 | Cell fate commitment, floral bud determinacy, and style development | Leaf and flower development | Nucleus | Axial regulator | INO | Outer integument growth | [47] |
| StYAB3 | Fruit development, polarity specification of abaxial-adaxial surface, regulation of shoot apical meristem, and leaf development | Leaf, flower, and fruit development | Nucleus | Phytophthora treated adult leaves, cadmium stress, dormant and non-dormant potato tubers, photoperiod effect, phloem associated cells | YABBY1/FIL | Axial regulator | [45] |
| StYAB4 | Fruit development, polarity specification of abaxial-adaxial surface, regulation of shoot apical meristem, and leaf development | Leaf, flower, and fruit development | Nucleus | Axial regulator | YABBY3 | Axial regulator | [47] |
| StYAB5 | Abaxial cell fate commitment | Leaf development | Chloroplast | Floral buds, cadmium stress, dormant and non-dormant potato tubers, photoperiod effect, phloem associated cells | YABBY2 | Putative axial regulator | [48,49] |
| StYAB6 | Abaxial cell fate commitment | Leaf development | Extra chromosomal | Developing tubers, cadmium stress, dormant and non-dormant potato tubers, photoperiod effect, phloem associated cells | YABBY5 | Axial regulator | [45,50] |
| StYAB7 | Abaxial cell fate commitment | Leaf development | Nucleus | Floral buds, cadmium stress, dormant and non-dormant potato tubers, photoperiod effect, phloem associated cells | YABBY2 | Putative axial regulator | [48,50] |
| Gene | Targeting miRNA | Function | Reference |
|---|---|---|---|
| StYAB3 | Stu-miR172d-5p | Enhance nodulation | [51] |
| StYAB4 | stu-miR5303g, stu-miR5303i, stu-miR5303h, stu-miR1886g-5p, stu-miR5303j, | Long Non-RNA development during tuber sprouting | [52] |
| StYAB5 | stu-miR8037 | No target site reported | [53] |
| StYAB6 | stu-miR7992-5p | Targets nuclear inclusion protein | [54] |
| StYAB7 | stu-miR390-3p, stu-miR1886g-5p | Promotes lateral growth | [55] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mazhar, H.S.-U.-D.; Shafiq, M.; Ali, H.; Ashfaq, M.; Anwar, A.; Tabassum, J.; Ali, Q.; Jilani, G.; Awais, M.; Sahu, R.; et al. Genome-Wide Identification, and In-Silico Expression Analysis of YABBY Gene Family in Response to Biotic and Abiotic Stresses in Potato (Solanum tuberosum). Genes 2023, 14, 824. https://doi.org/10.3390/genes14040824
Mazhar HS-U-D, Shafiq M, Ali H, Ashfaq M, Anwar A, Tabassum J, Ali Q, Jilani G, Awais M, Sahu R, et al. Genome-Wide Identification, and In-Silico Expression Analysis of YABBY Gene Family in Response to Biotic and Abiotic Stresses in Potato (Solanum tuberosum). Genes. 2023; 14(4):824. https://doi.org/10.3390/genes14040824
Chicago/Turabian StyleMazhar, Hafiz Sabah-Ud-Din, Muhammad Shafiq, Haider Ali, Muhammad Ashfaq, Alia Anwar, Javaria Tabassum, Qurban Ali, Ghulam Jilani, Muhammad Awais, Ravi Sahu, and et al. 2023. "Genome-Wide Identification, and In-Silico Expression Analysis of YABBY Gene Family in Response to Biotic and Abiotic Stresses in Potato (Solanum tuberosum)" Genes 14, no. 4: 824. https://doi.org/10.3390/genes14040824
APA StyleMazhar, H. S.-U.-D., Shafiq, M., Ali, H., Ashfaq, M., Anwar, A., Tabassum, J., Ali, Q., Jilani, G., Awais, M., Sahu, R., & Javed, M. A. (2023). Genome-Wide Identification, and In-Silico Expression Analysis of YABBY Gene Family in Response to Biotic and Abiotic Stresses in Potato (Solanum tuberosum). Genes, 14(4), 824. https://doi.org/10.3390/genes14040824