
Sequencing of the Pituitary Transcriptome after GnRH Treatment Uncovers the Involvement of lncRNA-m23b/miR-23b-3p/CAMK2D in FSH Synthesis and Secretion


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and GnRH Treatment
2.3. RNA Extraction
2.4. Library Construction
2.5. Sequencing, Clustering, Quality Control, and Inter-Sample Correlation Comparison
2.6. Identification of lncRNAs, mRNAs, and miRNAs
2.7. Differential Expression Analysis
2.8. Target Gene Prediction
2.9. Enrichment Analysis
2.10. Cell Culture
2.11. Cell Transfection
2.12. Real-Time Quantitative PCR Assay
2.13. Dual Luciferase Reporter Gene Assay
2.14. Western Blot Analysis
2.15. ELISA
2.16. Statistical Analysis
3. Results
3.1. Expression Analysis of mRNAs, lncRNAs, and miRNAs in the Rat Adenohypophysis
3.2. Analysis of Differential Gene Expression before and after GnRH Treatment
3.3. Interactions among lncRNAs, mRNAs, and miRNAs
3.4. GnRH Promotes the Target Binding of lncRNA-m23b to miR-23b-3p
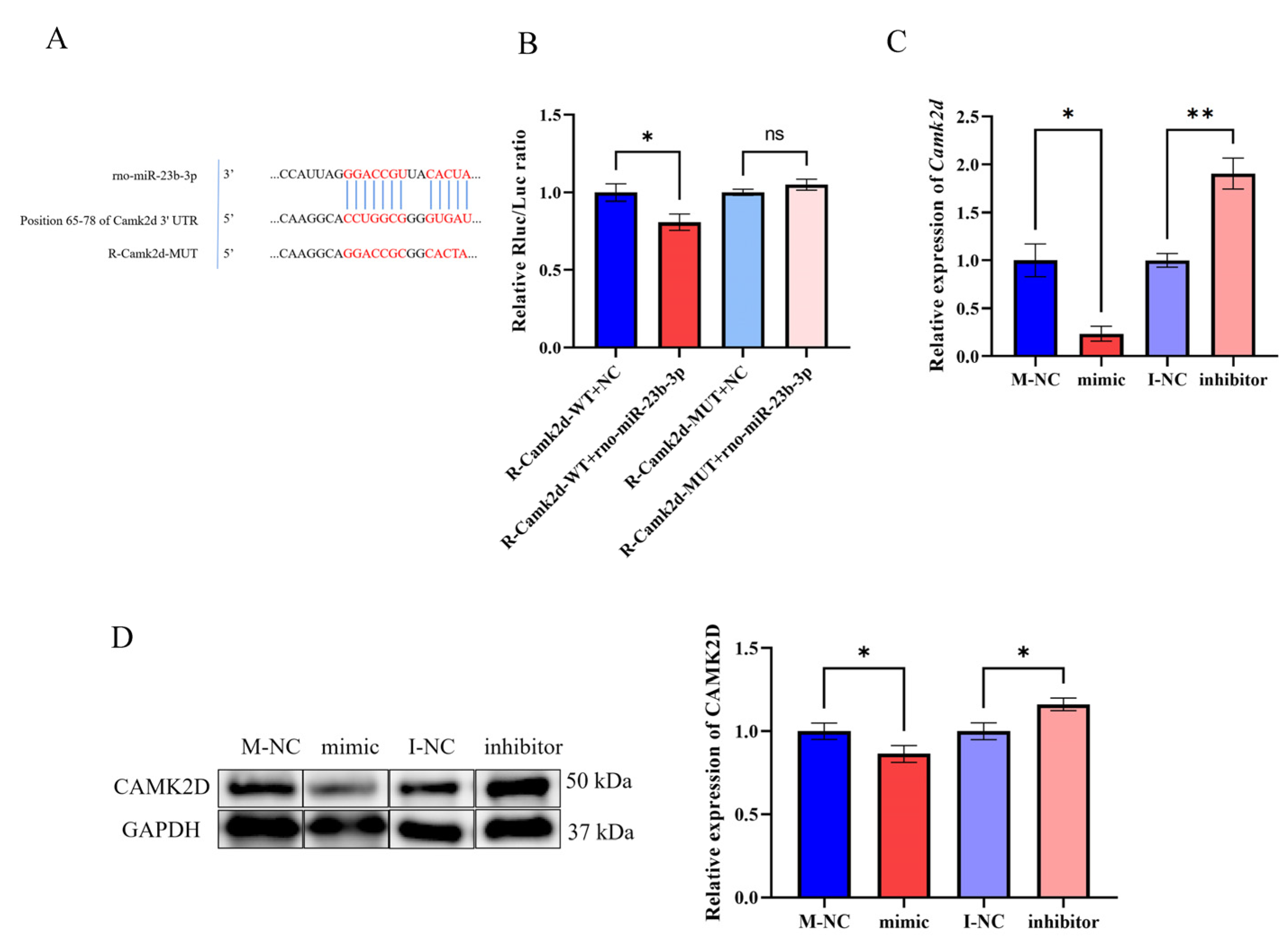
3.5. LncRNA-m23b Is Involved in the Regulation of the FSH Synthesis and Secretion by Targeting the Camk2d Expression through Competitive Binding with miR-23b-3p
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Amar, A.P.; Weiss, M.H. Pituitary anatomy and physiology. Neurosurg. Clin. N. Am. 2003, 14, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Brownstein, M.J.; Russell, J.T.; Gainer, H. Synthesis, transport, and release of posterior pituitary hormones. Science 1980, 207, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, S.A. The pituitary gland: A brief history. Pituitary 2007, 10, 323–325. [Google Scholar] [CrossRef]
- Edwards, W.; Raetzman, L.T. Complex integration of intrinsic and peripheral signaling is required for pituitary gland development. Biol. Reprod. 2018, 99, 504–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, E.; Kovacs, K. Fine structural cytology of the adenohypophysis in rat and man. J. Electron. Microsc. Tech. 1988, 8, 401–432. [Google Scholar] [CrossRef]
- Cohen, L.E.; Radovick, S. Molecular basis of combined pituitary hormone deficiencies. Endocr. Rev. 2002, 23, 431–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, J.; Duca, K.A.; Crowley, W.F., Jr. Gonadotropin-releasing hormone-induced stimulation and desensitization of free alpha-subunit secretion mirrors luteinizing hormone and follicle-stimulating hormone in perifused rat pituitary cells. Endocrinology 1990, 127, 2364–2371. [Google Scholar] [CrossRef]
- Ciccone, N.A.; Kaiser, U.B. The biology of gonadotroph regulation. Curr. Opin. Endocrinol. Diabetes Obes. 2009, 16, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Ye, R.S.; Xi, Q.Y.; Qi, Q.; Cheng, X.; Chen, T.; Li, H.; Kallon, S.; Shu, G.; Wang, S.B.; Jiang, Q.Y.; et al. Differentially expressed miRNAs after GnRH treatment and their potential roles in FSH regulation in porcine anterior pituitary cell. PLoS ONE 2013, 8, e57156. [Google Scholar] [CrossRef]
- Kanasaki, H.; Purwana, I.N.; Miyazaki, K. Possible role of PACAP and its PAC1 receptor in the differential regulation of pituitary LHbeta- and Fshbeta-subunit gene expression by pulsatile GnRH stimulation. Biol. Reprod. 2013, 88, 35. [Google Scholar] [CrossRef] [Green Version]
- Oleari, R.; Lettieri, A.; Paganoni, A.; Zanieri, L.; Cariboni, A. Semaphorin Signaling in GnRH Neurons: From Development to Disease. Neuroendocrinology 2019, 109, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Melamed, P.; Haj, M.; Yosefzon, Y.; Rudnizky, S.; Wijeweera, A.; Pnueli, L.; Kaplan, A. Multifaceted Targeting of the Chromatin Mediates Gonadotropin-Releasing Hormone Effects on Gene Expression in the Gonadotrope. Front. Endocrinol. 2018, 9, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shalev, D.; Melamed, P. The role of the hypothalamus and pituitary epigenomes in central activation of the reproductive axis at puberty. Mol. Cell. Endocrinol. 2020, 518, 111031. [Google Scholar] [CrossRef]
- Stamatiades, G.A.; Kaiser, U.B. Gonadotropin regulation by pulsatile GnRH: Signaling and gene expression. Mol. Cell. Endocrinol. 2018, 463, 131–141. [Google Scholar] [CrossRef]
- Dobkin-Bekman, M.; Naidich, M.; Pawson, A.J.; Millar, R.P.; Seger, R.; Naor, Z. Activation of mitogen-activated protein kinase (MAPK) by GnRH is cell-context dependent. Mol. Cell. Endocrinol. 2006, 252, 184–190. [Google Scholar] [CrossRef]
- Burger, L.L.; Haisenleder, D.J.; Aylor, K.W.; Marshall, J.C. Regulation of intracellular signaling cascades by GNRH pulse frequency in the rat pituitary: Roles for CaMK II, ERK, and JNK activation. Biol. Reprod. 2008, 79, 947–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, D.M.; Coss, D. PACAP induces Fshbeta gene expression via EPAC. Mol. Cell. Endocrinol. 2019, 492, 110438. [Google Scholar] [CrossRef]
- Tahir, M.S.; Porto-Neto, L.R.; Gondro, C.; Shittu, O.B.; Wockner, K.; Tan, A.W.L.; Smith, H.R.; Gouveia, G.C.; Kour, J.; Fortes, M.R.S. Meta-Analysis of Heifer Traits Identified Reproductive Pathways in Bos indicus Cattle. Genes 2021, 12, 768. [Google Scholar] [CrossRef]
- Haisenleder, D.J.; Burger, L.L.; Aylor, K.W.; Dalkin, A.C.; Marshall, J.C. Gonadotropin-releasing hormone stimulation of gonadotropin subunit transcription: Evidence for the involvement of calcium/calmodulin-dependent kinase II (Ca/CAMK II) activation in rat pituitaries. Endocrinology 2003, 144, 2768–2774. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Xu, C.; Liu, F.; Shen, X.; Meng, J.; Chen, H.; Yang, J.; Zhou, P.; Gao, R.; Gan, S. Effects of active immunization with newly modified GnRH peptides on spermatogenesis and production performance of Holstein bulls. Biol. Reprod. 2018, 99, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Fields, S.D.; Perry, B.L.; Perry, G.A. Effects of GnRH treatment on initiation of pulses of LH, LH release, and subsequent concentrations of progesterone. Domest. Anim. Endocrinol. 2009, 37, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.X.; Yuan, B.; Su, M.T.; Zheng, Y.; Zhang, J.Y.; Han, D.X.; Wang, H.Q.; Huang, Y.J.; Jiang, H.; Zhang, J.B. Identification of Circular RNAs in the Anterior Pituitary in Rats Treated with GnRH. Animals 2021, 11, 2557. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Q.; Wang, W.H.; Chen, C.Z.; Guo, H.X.; Jiang, H.; Yuan, B.; Zhang, J.B. Regulation of FSH Synthesis by Differentially Expressed miR-488 in Anterior Adenohypophyseal Cells. Animals 2021, 11, 3262. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Q.; Zhang, J.B.; Zheng, Y.; Zhang, W.D.; Guo, H.X.; Cong, S.; Ding, Y.; Yuan, B. Comprehensive analysis of differences in N6-methyladenosine RNA methylomes in the rat adenohypophysis after GnRH treatment. FASEB J. 2022, 36, e22204. [Google Scholar] [CrossRef]
- Kyritsi, K.; Meng, F.; Zhou, T.; Wu, N.; Venter, J.; Francis, H.; Kennedy, L.; Onori, P.; Franchitto, A.; Bernuzzi, F.; et al. Knockdown of Hepatic Gonadotropin-Releasing Hormone by Vivo-Morpholino Decreases Liver Fibrosis in Multidrug Resistance Gene 2 Knockout Mice by Down-Regulation of miR-200b. Am. J. Pathol. 2017, 187, 1551–1565. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Xu, S.; Ji, Z.; Sun, Y.; Cai, B.; Zhang, S.; Wang, P. The role of miR-7 as a potential switch in the mouse hypothalamus-pituitary-ovary axis through regulation of gonadotropins. Mol. Cell. Endocrinol. 2020, 518, 110969. [Google Scholar] [CrossRef]
- Yuen, T.; Ruf, F.; Chu, T.; Sealfon, S.C. Microtranscriptome regulation by gonadotropin-releasing hormone. Mol. Cell. Endocrinol. 2009, 302, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.J.; Gao, F.; Huang, Y.J.; Han, D.X.; Zheng, Y.; Wang, W.H.; Jiang, H.; Gao, Y.; Yuan, B.; Zhang, J.B. circAkap17b acts as a miR-7 family molecular sponge to regulate FSH secretion in rat pituitary cells. J. Mol. Endocrinol. 2020, 65, 135–148. [Google Scholar] [CrossRef]
- Huang, Y. The novel regulatory role of lncRNA-miRNA-mRNA axis in cardiovascular diseases. J. Cell. Mol. Med. 2018, 22, 5768–5775. [Google Scholar] [CrossRef]
- Han, D.X.; Sun, X.L.; Fu, Y.; Wang, C.J.; Liu, J.B.; Jiang, H.; Gao, Y.; Chen, C.Z.; Yuan, B.; Zhang, J.B. Identification of long non-coding RNAs in the immature and mature rat anterior pituitary. Sci. Rep. 2017, 7, 17780. [Google Scholar] [CrossRef] [Green Version]
- Han, D.X.; Sun, X.L.; Wang, C.J.; Yu, Z.W.; Zheng, Y.; Huang, Y.J.; Wang, W.H.; Jiang, H.; Gao, Y.; Yuan, B.; et al. Differentially expressed lncRNA-m433s1 regulates FSH secretion by functioning as a miRNA sponge in male rat anterior pituitary cellsdagger. Biol. Reprod. 2019, 101, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Ren, W.; Han, D.; Zhao, G.; Wang, H.; Guo, H.; Zheng, Y.; Ji, Z.; Gao, W.; Yuan, B. LncRNA-m18as1 competitively binds with miR-18a-5p to regulate follicle-stimulating hormone secretion through the Smad2/3 pathway in rat primary pituitary cells. J. Zhejiang Univ. Sci. B 2022, 23, 502–514. [Google Scholar] [CrossRef] [PubMed]
- Ewing, B.; Green, P. Base-calling of automated sequencer traces using phred. II. Error probabilities. Genome Res. 1998, 8, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Luo, H.; Bu, D.; Zhao, G.; Yu, K.; Zhang, C.; Liu, Y.; Chen, R.; Zhao, Y. Utilizing sequence intrinsic composition to classify protein-coding and long non-coding transcripts. Nucleic Acids Res. 2013, 41, e166. [Google Scholar] [CrossRef]
- Kong, L.; Zhang, Y.; Ye, Z.Q.; Liu, X.Q.; Zhao, S.Q.; Wei, L.; Gao, G. CPC: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res. 2007, 35, W345–W349. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Park, H.J.; Dasari, S.; Wang, S.; Kocher, J.P.; Li, W. CPAT: Coding-Potential Assessment Tool using an alignment-free logistic regression model. Nucleic Acids Res. 2013, 41, e74. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [Green Version]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Fahlgren, N.; Howell, M.D.; Kasschau, K.D.; Chapman, E.J.; Sullivan, C.M.; Cumbie, J.S.; Givan, S.A.; Law, T.F.; Grant, S.R.; Dangl, J.L.; et al. High-throughput sequencing of Arabidopsis microRNAs: Evidence for frequent birth and death of MIRNA genes. PLoS ONE 2007, 2, e219. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, D.X.; Sun, X.L.; Xu, M.Q.; Chen, C.Z.; Jiang, H.; Gao, Y.; Yuan, B.; Zhang, J.B. Roles of differential expression of microRNA-21-3p and microRNA-433 in FSH regulation in rat anterior pituitary cells. Oncotarget 2017, 8, 36553–36565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaprara, A.; Huhtaniemi, I.T. The hypothalamus-pituitary-gonad axis: Tales of mice and men. Metabolism 2018, 86, 3–17. [Google Scholar] [CrossRef]
- Burger, L.L.; Haisenleder, D.J.; Aylor, K.W.; Marshall, J.C. Regulation of Lhb and Egr1 gene expression by GNRH pulses in rat pituitaries is both c-Jun N-terminal kinase (JNK)- and extracellular signal-regulated kinase (ERK)-dependent. Biol. Reprod. 2009, 81, 1206–1215. [Google Scholar] [CrossRef] [Green Version]
- Harris, D.; Bonfil, D.; Chuderland, D.; Kraus, S.; Seger, R.; Naor, Z. Activation of MAPK cascades by GnRH: ERK and Jun N-terminal kinase are involved in basal and GnRH-stimulated activity of the glycoprotein hormone LHbeta-subunit promoter. Endocrinology 2002, 143, 1018–1025. [Google Scholar] [CrossRef]
- Okumu, L.A.; Braden, T.D.; Vail, K.; Simon, L.; Goyal, H.O. Low androgen induced penile maldevelopment involves altered gene expression of biomarkers of smooth muscle differentiation and a key enzyme regulating cavernous smooth muscle cell tone. J. Urol. 2014, 192, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Alen, F.; Gomez-Redondo, I.; Rivera, P.; Suarez, J.; Ramos-Ibeas, P.; Pericuesta, E.; Fernandez-Gonzalez, R.; Perez-Cerezales, S.; Horiuchi, K.; Orio, L.; et al. Sex-Dimorphic Behavioral Alterations and Altered Neurogenesis in U12 Intron Splicing-Defective Zrsr1 Mutant Mice. Int. J. Mol. Sci. 2019, 20, 3543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farshori, P.Q.; Shah, B.H.; Arora, K.K.; Martinez-Fuentes, A.; Catt, K.J. Activation and nuclear translocation of PKCdelta, Pyk2 and ERK1/2 by gonadotropin releasing hormone in HEK293 cells. J. Steroid Biochem. Mol. Biol. 2003, 85, 337–347. [Google Scholar] [CrossRef]
- Melamed, P.; Savulescu, D.; Lim, S.; Wijeweera, A.; Luo, Z.; Luo, M.; Pnueli, L. Gonadotrophin-releasing hormone signalling downstream of calmodulin. J. Neuroendocrinol. 2012, 24, 1463–1475. [Google Scholar] [CrossRef]
- Qian, X.; Lin, G.; Wang, J.; Zhang, S.; Ma, J.; Yu, B.; Wu, R.; Liu, M. CircRNA_01477 influences axonal growth via regulating miR-3075/FosB/Stat3 axis. Exp. Neurol. 2022, 347, 113905. [Google Scholar] [CrossRef]
- Chen, J.; An, B.S.; Cheng, L.; Hammond, G.L.; Leung, P.C. Gonadotropin-releasing hormone-mediated phosphorylation of estrogen receptor-alpha contributes to fosB expression in mouse gonadotrophs. Endocrinology 2009, 150, 4583–4593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.J.; Guo, H.X.; Han, D.X.; Yu, Z.W.; Zheng, Y.; Jiang, H.; Gao, Y.; Yuan, B.; Zhang, J.B. Pituitary tissue-specific miR-7a-5p regulates FSH expression in rat anterior adenohypophyseal cells. PeerJ 2019, 7, e6458. [Google Scholar] [CrossRef] [PubMed]
- Han, D.X.; Xiao, Y.; Wang, C.J.; Jiang, H.; Gao, Y.; Yuan, B.; Zhang, J.B. Regulation of FSH expression by differentially expressed miR-186-5p in rat anterior adenohypophyseal cells. PLoS ONE 2018, 13, e0194300. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Xu, J.; Luo, Y.; Zou, D.; Han, R.; Zhong, S.; Zhao, Q.; Mang, X.; Li, M.; Si, Y.; et al. Panoramic transcriptome analysis and functional screening of long noncoding RNAs in mouse spermatogenesis. Genome Res. 2021, 31, 13–26. [Google Scholar] [CrossRef]
- Wang, J.; Wang, L.; Feng, G.; Wang, Y.; Li, Y.; Li, X.; Liu, C.; Jiao, G.; Huang, C.; Shi, J.; et al. Asymmetric Expression of LincGET Biases Cell Fate in Two-Cell Mouse Embryos. Cell 2018, 175, 1887–1901.e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhang, X.; Dang, Y.; Li, D.; Lu, G.; Chan, W.Y.; Leung, P.C.K.; Zhao, S.; Qin, Y.; Chen, Z.J. Long noncoding RNA HCP5 participates in premature ovarian insufficiency by transcriptionally regulating MSH5 and DNA damage repair via YB1. Nucleic Acids Res. 2020, 48, 4480–4491. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhang, S.; Chen, X.; Li, N.; Li, J.; Jia, R.; Pan, Y.; Liang, H. CircNT5E Acts as a Sponge of miR-422a to Promote Glioblastoma Tumorigenesis. Cancer Res. 2018, 78, 4812–4825. [Google Scholar] [CrossRef] [Green Version]
- Liang, H.; Yu, T.; Han, Y.; Jiang, H.; Wang, C.; You, T.; Zhao, X.; Shan, H.; Yang, R.; Yang, L.; et al. LncRNA PTAR promotes EMT and invasion-metastasis in serous ovarian cancer by competitively binding miR-101-3p to regulate ZEB1 expression. Mol. Cancer 2018, 17, 119. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Hanson, P.I.; Schulman, H. Neuronal Ca2+/calmodulin-dependent protein kinases. Annu. Rev. Biochem. 1992, 61, 559–601. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.J.; Hidaka, H.; Dannies, P.S. KN-62, a calcium/calmodulin-dependent protein kinase II inhibitor, inhibits high potassium-stimulated prolactin secretion and intracellular calcium increases in anterior pituitary cells. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 1996, 1310, 343–347. [Google Scholar] [CrossRef] [Green Version]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Zhao, G.; Yu, S.; Zheng, Y.; Guo, H.; Wang, H.; Zhao, P.; Xie, W.; Ren, W.; Yuan, B. Sequencing of the Pituitary Transcriptome after GnRH Treatment Uncovers the Involvement of lncRNA-m23b/miR-23b-3p/CAMK2D in FSH Synthesis and Secretion. Genes 2023, 14, 846. https://doi.org/10.3390/genes14040846
Wang T, Zhao G, Yu S, Zheng Y, Guo H, Wang H, Zhao P, Xie W, Ren W, Yuan B. Sequencing of the Pituitary Transcriptome after GnRH Treatment Uncovers the Involvement of lncRNA-m23b/miR-23b-3p/CAMK2D in FSH Synthesis and Secretion. Genes. 2023; 14(4):846. https://doi.org/10.3390/genes14040846
Chicago/Turabian StyleWang, Tian, Guokun Zhao, Song Yu, Yi Zheng, Haixiang Guo, Haoqi Wang, Peisen Zhao, Wenyin Xie, Wenzhi Ren, and Bao Yuan. 2023. "Sequencing of the Pituitary Transcriptome after GnRH Treatment Uncovers the Involvement of lncRNA-m23b/miR-23b-3p/CAMK2D in FSH Synthesis and Secretion" Genes 14, no. 4: 846. https://doi.org/10.3390/genes14040846