
Vitrification with Dimethyl Sulfoxide Induces Transcriptomic Alteration of Gene and Transposable Element Expression in Immature Human Oocytes

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and Oocyte Collection
2.2. Oocyte Cryopreservation
2.3. Oocyte Thawing
2.4. RNA Sequencing and Gene Analysis
2.5. Statistical Analyses
2.6. RNA Sequencing
2.6.1. Quality Control Analysis
2.6.2. Gene Mapping
2.6.3. Gene Quantification
2.7. Transposable Element Expression Profiling
2.8. Real Time Quantitative PCR (RT-qPCR)
3. Results
3.1. Differential Expression Analysis
- Genes Upregulated After Vitrification
- Genes Downregulated After Vitrification
3.2. Functional Enrichment Analyses
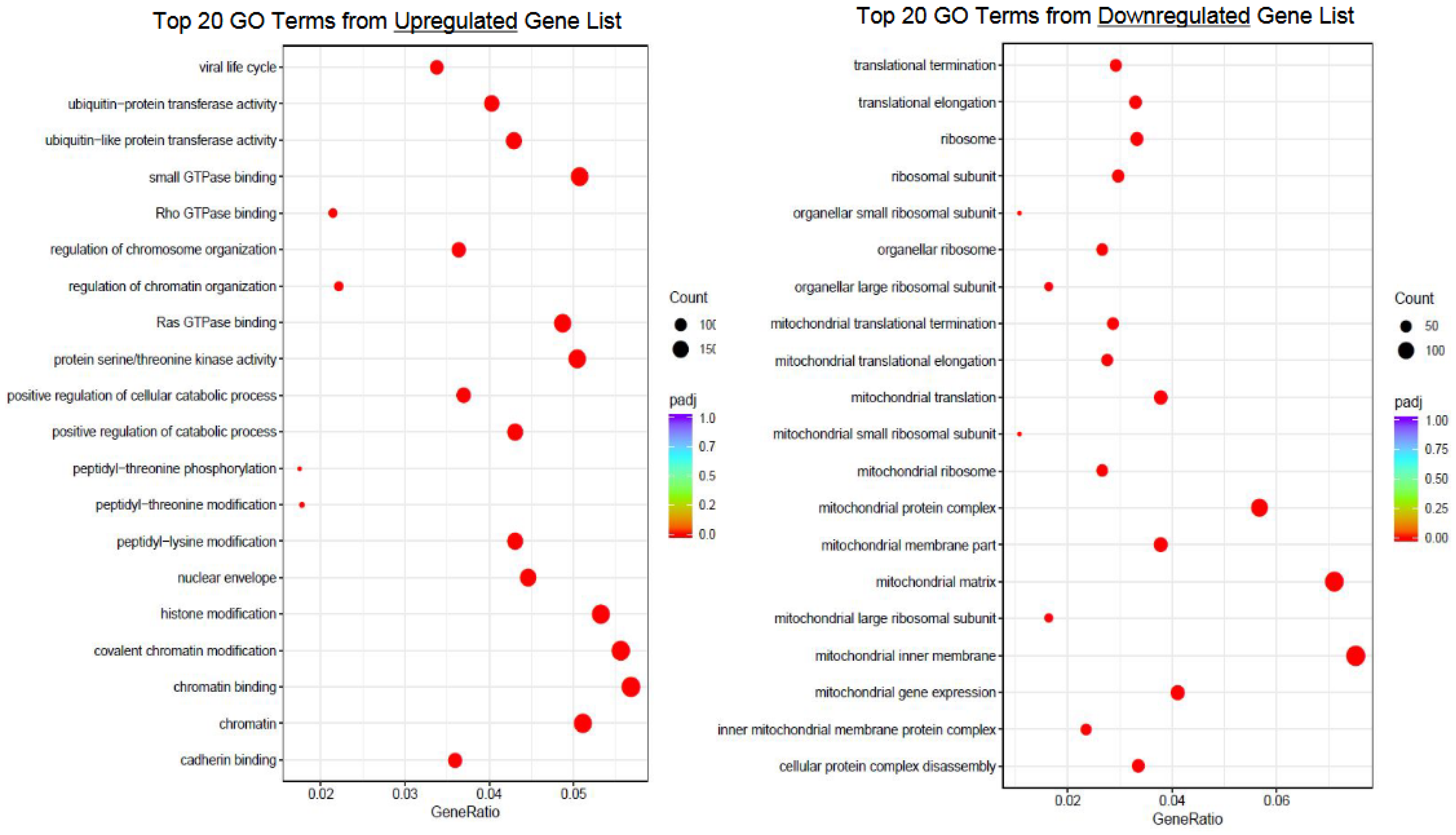
- Gene Ontology (GO)
- Kyoto Encyclopedia of Genes and Genomes (KEGG)
- Human Disease Ontology (HDO)
3.3. Transposable Element (TE) Expression
3.4. Real Time Quantitative PCR
4. Discussion
4.1. Differential Gene Expression and Gene Enrichment Analyses
4.2. Differential Transposable Element Expression
4.3. Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Practice Committees of the American Society for Reproductive Medicine and the Society for Assisted Reproductive Technology. Mature oocyte cryopreservation: A guideline. Fertil. Steril. 2013, 99, 37–43. [Google Scholar] [CrossRef]
- Barberet, J.; Barry, F.; Choux, C.; Guilleman, M.; Karoui, S.; Simonot, R.; Bruno, C.; Fauque, P. What impact does oocyte vitrification have on epigenetics and gene expression? Clin. Epigenetics 2020, 12, 121. [Google Scholar] [CrossRef]
- Forman, E.J.; Li, X.; Ferry, K.M.; Scott, K.; Treff, N.R.; Scott, R.T., Jr. Oocyte vitrification does not increase the risk of embryonic aneuploidy or diminish the implantation potential of blastocysts created after intracytoplasmic sperm injection: A novel, paired randomized controlled trial using DNA fingerprinting. Fertil. Steril. 2012, 98, 644–649. [Google Scholar] [CrossRef]
- Goldman, K.N.; Noyes, N.L.; Knopman, J.M.; McCaffrey, C.; Grifo, J.A. Oocyte efficiency: Does live birth rate differ when analyzing cryopreserved and fresh oocytes on a per-oocyte basis? Fertil. Steril. 2013, 100, 712–717. [Google Scholar] [CrossRef]
- Kushnir, V.A.; Darmon, S.K.; Barad, D.H.; Gleicher, N. New national outcome data on fresh versus cryopreserved donor oocytes. J. Ovarian Res. 2018, 11, 2. [Google Scholar] [CrossRef]
- Spector, L.G.; Brown, M.B.; Wantman, E.; Letterie, G.S.; Toner, J.P.; Doody, K.; Ginsburg, E.; Williams, M.; Koch, L.; Schymura, M.J.; et al. Association of In Vitro Fertilization With Childhood Cancer in the United States. JAMA Pediatr. 2019, 173, e190392. [Google Scholar] [CrossRef] [PubMed]
- Hargreave, M.; Jensen, A.; Hansen, M.K.; Dehlendorff, C.; Winther, J.F.; Schmiegelow, K.; Kjær, S.K. Association Between Fertility Treatment and Cancer Risk in Children. JAMA 2019, 322, 2203–2210. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Chen, L.; Yang, T.; Wang, L.; Zhao, L.; Zhang, S.; Ye, Z.; Chen, L.; Zheng, Z.; Qin, J. Cancer risk among children conceived by fertility treatment. Int. J. Cancer 2019, 144, 3001–3013. [Google Scholar] [CrossRef] [PubMed]
- Berntsen, S.; Pinborg, A. Large for gestational age and macrosomia in single-tons born after frozen/thawed embryo transfer (FET) in assisted reproductivetechnology (ART). Birth Defects Res. 2018, 110, 630–643. [Google Scholar] [CrossRef]
- Cantone, I.; Fisher, A.G. Epigenetic programming and reprogramming during development. Nat. Struct. Mol. Biol. 2013, 20, 282–289. [Google Scholar] [CrossRef]
- Smallwood, S.A.; Kelsey, G. De novo DNA methylation: A germ cell perspective. Trends Genet. 2012, 28, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Wyse, B.A.; Fuchs Weizman, N.; Kadish, S.; Balakier, H.; Sangaralingam, M.; Librach, C.L. Transcriptomics of cumulus cells—A window into oocyte maturation in humans. J. Ovarian Res. 2020, 13, 93. [Google Scholar] [CrossRef]
- Whaley, D.; Damyar, K.; Witek, R.P.; Mendoza, A.; Alexander, M.; Lakey, J.R. Cryopreservation: An Overview of Principles and Cell-Specific Considerations. Cell Transplant. 2021, 30, 963689721999617. [Google Scholar] [CrossRef]
- Choi, T. Dimethyl sulfoxide inhibits spontaneous oocyte fragmentation and delays inactivation of maturation promoting factor (MPF) during the prolonged culture of ovulated murine oocytes in vitro. Cytotechnology 2011, 63, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Alsalim, H.; Jafarpour, F.; Ghazvini Zadegan, F.; Nasr-Esfahani, M.H.; Niasari-Naslaji, A. Epigenotoxic Effect of Dimethyl Sulfoxide on Buffalo Somatic Cells and Buffalo-Bovine Interspecies Somatic Cell Nuclear Transfer Embryos. Cell J. 2019, 20, 544–551. [Google Scholar] [CrossRef]
- Zhou, D.; Shen, X.; Gu, Y.; Zhang, N.; Li, T.; Wu, X.; Lei, L. Effects of dimethyl sulfoxide on asymmetric division and cytokinesis in mouse oocytes. BMC Dev. Biol. 2014, 14, 28. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Wang, Y.K.; Song, Z.Q.; Du, Z.Q.; Yang, C.X. Dimethyl Sulfoxide Perturbs Cell Cycle Progression and Spindle Organization in Porcine Meiotic Oocytes. PLoS ONE 2016, 11, e0158074. [Google Scholar] [CrossRef] [Green Version]
- Verheijen, M.; Lienhard, M.; Schrooders, Y.; Clayton, O.; Nudischer, R.; Boerno, S.; Timmermann, B.; Selevsek, N.; Schlapbach, R.; Gmuender, H.; et al. DMSO induces drastic changes in human cellular processes and epigenetic landscape in vitro. Sci. Rep. 2019, 9, 4641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaler, R.; Spitzer, S.; Karlic, H.; Klaushofer, K.; Varga, F. DMSO is a strong inducer of DNA hydroxymethylation in pre-osteoblastic MC3T3-E1 cells. Epigenetics 2012, 7, 635–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monzo, C.; Haouzi, D.; Roman, K.; Assou, S.; Dechaud, H.; Hamamah, S. Slow freezing and vitrification differentially modify the gene expression profile of human metaphase ii oocytes. Hum. Reprod. 2012, 27, 2160–2168. [Google Scholar] [CrossRef]
- Chamayou, S.; Bonaventura, G.; Alecci, C.; Tibullo, D.; Di Raimondo, F.; Guglielmino, A.; Barcellona, M. Consequences of metaphase ii oocyte cryopreservation on Mrna content. Cryobiology 2011, 62, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Tunbak, H.; Enriquez-Gasca, R.; Tie, C.H.C.; Gould, P.A.; Mlcochova, P.; Gupta, R.K.; Fernandes, L.; Holt, J.; van der Veen, A.G.; Giampazolias, E.; et al. The HUSH complex is a gatekeeper of type I interferon through epigenetic regulation of LINE-1s. Nat. Commun. 2020, 11, 5387. [Google Scholar] [CrossRef]
- Pray, L. Transposons: The Jumping Genes. Nat. Educ. 2008, 1, 204. Available online: https://www.nature.com/scitable/topicpage/transposons-the-jumping-genes-518/ (accessed on 1 January 2022).
- Ardeljan, D.; Steranka, J.P.; Liu, C.; Li, Z.; Taylor, M.S.; Payer, L.M.; Gorbounov, M.; Sarnecki, J.S.; Deshpande, V.; Hruban, R.H.; et al. Cell fitness screens reveal a conflict between LINE-1 retrotransposition and DNA replication. Nat. Struct. Mol. Biol. 2020, 27, 168–178. [Google Scholar] [CrossRef]
- De Cecco, M.; Ito, T.; Petrashen, A.P.; Elias, A.E.; Skvir, N.J.; Criscione, S.W.; Caligiana, A.; Brocculi, G.; Adney, E.M.; Boeke, J.D.; et al. L1 drives IFN in senescent cells and promotes age-associated inflammation. Nature 2019, 566, 73–78, Erratum in Nature 2019, 572, E5. [Google Scholar] [CrossRef] [PubMed]
- Torres-Padilla, M.E. On transposons and totipotency. Philos. Trans. R Soc. Lond B Biol. Sci. 2020, 375, 20190339. [Google Scholar] [CrossRef] [PubMed]
- Virant-Klun, I.; Knez, K.; Tomazevic, T.; Skutella, T. Gene expression profiling of human oocytes developed and matured in vivo or in vitro. Biomed. Res. Int. 2013, 2013, 879489. [Google Scholar] [CrossRef] [Green Version]
- Hasuwa, H.; Iwasaki, Y.W.; Au Yeung, W.K.; Ishino, K.; Masuda, H.; Sasaki, H.; Siomi, H. Production of functional oocytes requires maternally expressed PIWI genes and piRNAs in golden hamsters. Nat. Cell Biol. 2021, 23, 1002–1012. [Google Scholar] [CrossRef]
- Cantatore, C.; George, J.S.; Depalo, R.; D’Amato, G.; Moravek, M.; Smith, G.D. Mouse oocyte vitrification with and without dimethyl sulfoxide: Influence on cryo-survival, development, and maternal imprinted gene expression. J. Assist. Reprod. Genet. 2021, 38, 2129–2138. [Google Scholar] [CrossRef]
- Habara, O.; Logan, C.Y.; Kanai-Azuma, M.; Nusse, R.; Takase, H.M. WNT signaling in pre-granulosa cells is required for ovarian folliculogenesis and female fertility. Development 2021, 148, dev198846. [Google Scholar] [CrossRef]
- Boyer, A.; Goff, A.K.; Boerboom, D. WNT signaling in ovarian follicle biology and tumorigenesis. Trends Endocrinol. Metab. 2010, 21, 25–32. [Google Scholar] [CrossRef]
- Theka, I.; Sottile, F.; Cammisa, M.; Bonnin, S.; Sanchez-Delgado, M.; Di Vicino, U.; Neguembor, M.V.; Arumugam, K.; Aulicino, F.; Monk, D.; et al. Wnt/β-catenin signaling pathway safeguards epigenetic stability and homeostasis of mouse embryonic stem cells. Sci. Rep. 2019, 9, 948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laskowski, D.; Sjunnesson, Y.; Humblot, P.; Andersson, G.; Gustafsson, H.; Båge, R. The functional role of insulin in fertility and embryonic development-What can we learn from the bovine model? Theriogenology 2016, 86, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Templeman, N.M.; Luo, S.; Kaletsky, R.; Shi, C.; Ashraf, J.; Keyes, W.; Murphy, C.T. Insulin Signaling Regulates Oocyte Quality Maintenance with Age via Cathepsin B Activity. Curr. Biol. 2018, 28, 753–760.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, J.; Scaramuzzi, R.J. Insulin signalling and glucose transport in the ovary and ovarian function during the ovarian cycle. Biochem. J. 2016, 473, 1483–1501. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Yu, Q. Role of mTOR Signaling in Female Reproduction. Front. Endocrinol. 2019, 10, 692. [Google Scholar] [CrossRef]
- Hwang, M.; Perez, C.A.; Moretti, L.; Lu, B. The mTOR signaling network: Insights from its role during embryonic development. Curr. Med. Chem. 2008, 15, 1192–1208. [Google Scholar] [CrossRef]
- Goldman, K.N.; Chenette, D.; Arju, R.; Duncan, F.E.; Keefe, D.L.; Grifo, J.A.; Schneider, R.J. mTORC1/2 inhibition preserves ovarian function and fertility during genotoxic chemotherapy. Proc. Natl. Acad. Sci. USA 2017, 114, 3186–3191. [Google Scholar] [CrossRef] [Green Version]
- Papageorgiou, K.; Mastora, E.; Zikopoulos, A.; Grigoriou, M.E.; Georgiou, I.; Michaelidis, T.M. Interplay Between mTOR and Hippo Signaling in the Ovary: Clinical Choice Guidance Between Different Gonadotropin Preparations for Better IVF. Front. Endocrinol. 2021, 12, 702446. [Google Scholar] [CrossRef]
- Wu, Z.; Guan, K.L. Hippo Signaling in Embryogenesis and Development. Trends Biochem. Sci. 2021, 46, 51–63. [Google Scholar] [CrossRef]
- Madan, P.; Calder, M.D.; Watson, A.J. Mitogen-activated protein kinase (MAPK) blockade of bovine preimplantation embryogenesis requires inhibition of both p38 and extracellular signal-regulated kinase (ERK) pathways. Reproduction 2005, 130, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, R.M.; Stein, P.; Svoboda, P. The oocyte-to-embryo transition in mouse: Past, present, and future. Biol. Reprod. 2018, 99, 160–174. [Google Scholar] [CrossRef]
- Medvedev, S.; Pan, H.; Schultz, R.M. Absence of MSY2 in mouse oocytes perturbs oocyte growth and maturation, RNA stability, and the transcriptome. Biol. Reprod. 2011, 85, 575–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Yan, Z.; Qin, Q.; Nisenblat, V.; Chang, H.M.; Yu, Y.; Wang, T.; Lu, C.; Yang, M.; Yang, S.; et al. Transcriptome Landscape of Human Folliculogenesis Reveals Oocyte and Granulosa Cell Interactions. Mol. Cell. 2018, 72, 1021–1034.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De La Fuente, R.; Eppig, J.J. Transcriptional activity of the mouse oocyte genome: Companion granulosa cells modulate transcription and chromatin remodeling. Dev. Biol. 2001, 229, 224–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, S.; Tyler, J.K. Epigenetics and aging. Sci. Adv. 2016, 2, e1600584. [Google Scholar] [CrossRef] [Green Version]
- Gorbunova, V.; Seluanov, A.; Mita, P.; McKerrow, W.; Fenyö, D.; Boeke, J.D.; Linker, S.B.; Gage, F.H.; Kreiling, J.A.; Petrashen, A.P.; et al. The role of retrotransposable elements in ageing and age-associated diseases. Nature 2021, 596, 43–53. [Google Scholar] [CrossRef]
- Doyle, J.O.; Richter, K.S.; Lim, J.; Stillman, R.J.; Graham, J.R.; Tucker, M.J. Successful elective and medically indicated oocyte vitrification and warming for autologous in vitro fertilization, with predicted birth probabilities for fertility preservation according to number of cryopreserved oocytes and age at retrieval. Fertil. Steril. 2016, 105, 459–466.e2. [Google Scholar] [CrossRef] [Green Version]
- Nagy, Z.P.; Anderson, R.E.; Feinberg, E.C.; Hayward, B.; Mahony, M.C. The Human Oocyte Preservation Experience (HOPE) Registry: Evaluation of cryopreservation techniques and oocyte source on outcomes. Reprod. Biol. Endocrinol. 2017, 15, 10. [Google Scholar] [CrossRef] [Green Version]
- Steuerwald, N.M.; Bermúdez, M.G.; Wells, D.; Munné, S.; Cohen, J. Maternal age-related differential global expression profiles observed in human oocytes. Reprod. Biomed. Online 2007, 14, 700–708. [Google Scholar] [CrossRef]
- Deininger, P. Alu elements: Know the SINEs. Genome Biol. 2011, 12, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegenhain, C.; Vieth, B.; Parekh, S.; Reinius, B.; Guillaumet-Adkins, A.; Smets, M.; Leonhardt, H.; Heyn, H.; Hellmann, I.; Enard, W. Comparative Analysis of Single-Cell RNA Sequencing Methods. Mol. Cell. 2017, 65, 631–643.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whiteley, G.E.; Martini, A.E.; Jahandideh, S.; Devine, K.; Hill, M.J.; DeCherney, A.H.; Doyle, J.; Kallen, C. The Impact of duration of oocyte cryopreservation on live birth outcomes in IVF cycles using autologous thawed oocytes. Fertil. Steril. 2021, 116, e9. [Google Scholar] [CrossRef]
- Parmegiani, L.; Garello, C.; Granella, F.; Guidetti, D.; Bernardi, S.; Cognigni, G.E.; Revelli, A.; Filicori, M. Long-term cryostorage does not adversely affect the outcome of oocyte thawing cycles. Reprod. Biomed. Online 2009, 19, 374–379. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient Clinical and Cycle Characteristics | ||||||||
|---|---|---|---|---|---|---|---|---|
| Subject |
Age (Years) |
BMI (kg/m²) |
AMH (ng/mL) |
FSH (mIU/mL) |
E2 on Trigger Day/1 Day Post Trigger (pg/mL) | Days of Stimulation | Oocytes Retrieved/MII | Vitrified/Non-Vitrified |
| 1 | 30 | 21 | 2.6 | 6.5 | 2149/2516 | 9 | 10/5 | 2/2 |
| 2 | 35 | 21 | 3.3 | 4.4 | 4188/4765 | 11 | 29/12 | 4/4 |
| 3 | 27 | 31 | 8.2 | 4.8 | 4936/6659 | 12 | 39/32 | 3/3 |
| 4 | 34 | 21 | 8.6 | 7.6 | 2832/4486 | 10 | 32/22 | 3/3 |
| TE Correlation to PIWIL2, DNMT3A, DNTM3B, and Age | ||
|---|---|---|
| Positive Correlation | Negative Correlation | |
| PIWIL2 | Ancient HERVs: LTR41B, MER51A SINE: MamSINE1 DNA TEs: MER102a, Tigger11a, Tigger3d | |
| DNMT3A | Ancient(mammalian) L1s: L1MB8, L1MC2:L1:LINE, L1ME4b:L1:LINE, LTR41B:ERVL:LT MamSINE1, MIR ERV1 LTRs: MER51A DNATEs: Tigger11a, Tigger3d | |
| DNMT3B | Ancient (mammalian) L1s: L1M2a1, L1M5 A DNA TE: MER63C | MLT1J |
| Age | Ancient LINE-1: HAL1ME | HERVs: LTR14B, LTR1C3, LTR7C, MLT1O |
| Primer Name | Sequence |
|---|---|
| ERV1-1-F | TGG ACC TCT CAC AAC ACA AAC T |
| ERV1-1-R | AGG GGA ATT CCA GTG GGT CT |
| ERV1-2-F | ACGCTTTACAGCCCTAGACC |
| ERV1-2-R | GTCGGGAGCAGATTGGGTAA |
| ERVK-1-F | GGC CAT CAG AGT CTA AAC CAC G |
| ERVK-1-R | CTG ACT TTC TGG GGG TGG CCG |
| ERVK-2-F | GGG TAC CTG GCC CCA TAG AT |
| ERVK-2-R | CAT CAT CCC TTC TTC CTC AGG TT |
| GAPDH-R | CCC TTT TGG CTC CAC CCT |
| GAPDH-F | TTC ACC ACC ATG GAG AAG GC |
| DNMT3A-F | CAA GGA GGA GCG CCA AGA |
| DNMT3A-R | ACT TGG AGA TCA CCG CAG G |
| DNMT3B-F | CCC AGC TCT TAC CTT ACC ATC G |
| DNMT3B-R | GGTCCCCTATTCCAAACTCCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiltshire, A.; Schaal, R.; Wang, F.; Tsou, T.; McKerrow, W.; Keefe, D. Vitrification with Dimethyl Sulfoxide Induces Transcriptomic Alteration of Gene and Transposable Element Expression in Immature Human Oocytes. Genes 2023, 14, 1232. https://doi.org/10.3390/genes14061232
Wiltshire A, Schaal R, Wang F, Tsou T, McKerrow W, Keefe D. Vitrification with Dimethyl Sulfoxide Induces Transcriptomic Alteration of Gene and Transposable Element Expression in Immature Human Oocytes. Genes. 2023; 14(6):1232. https://doi.org/10.3390/genes14061232
Chicago/Turabian StyleWiltshire, Ashley, Renata Schaal, Fang Wang, Tiffany Tsou, Wilson McKerrow, and David Keefe. 2023. "Vitrification with Dimethyl Sulfoxide Induces Transcriptomic Alteration of Gene and Transposable Element Expression in Immature Human Oocytes" Genes 14, no. 6: 1232. https://doi.org/10.3390/genes14061232
APA StyleWiltshire, A., Schaal, R., Wang, F., Tsou, T., McKerrow, W., & Keefe, D. (2023). Vitrification with Dimethyl Sulfoxide Induces Transcriptomic Alteration of Gene and Transposable Element Expression in Immature Human Oocytes. Genes, 14(6), 1232. https://doi.org/10.3390/genes14061232