
Genetic Diversity of Common Olive (Olea europaea L.) Cultivars from Nikita Botanical Gardens Collection Revealed Using RAD-Seq Method

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and DNA Extraction
2.2. RAD Libraries Preparation and Sequencing
2.3. Data Analysis
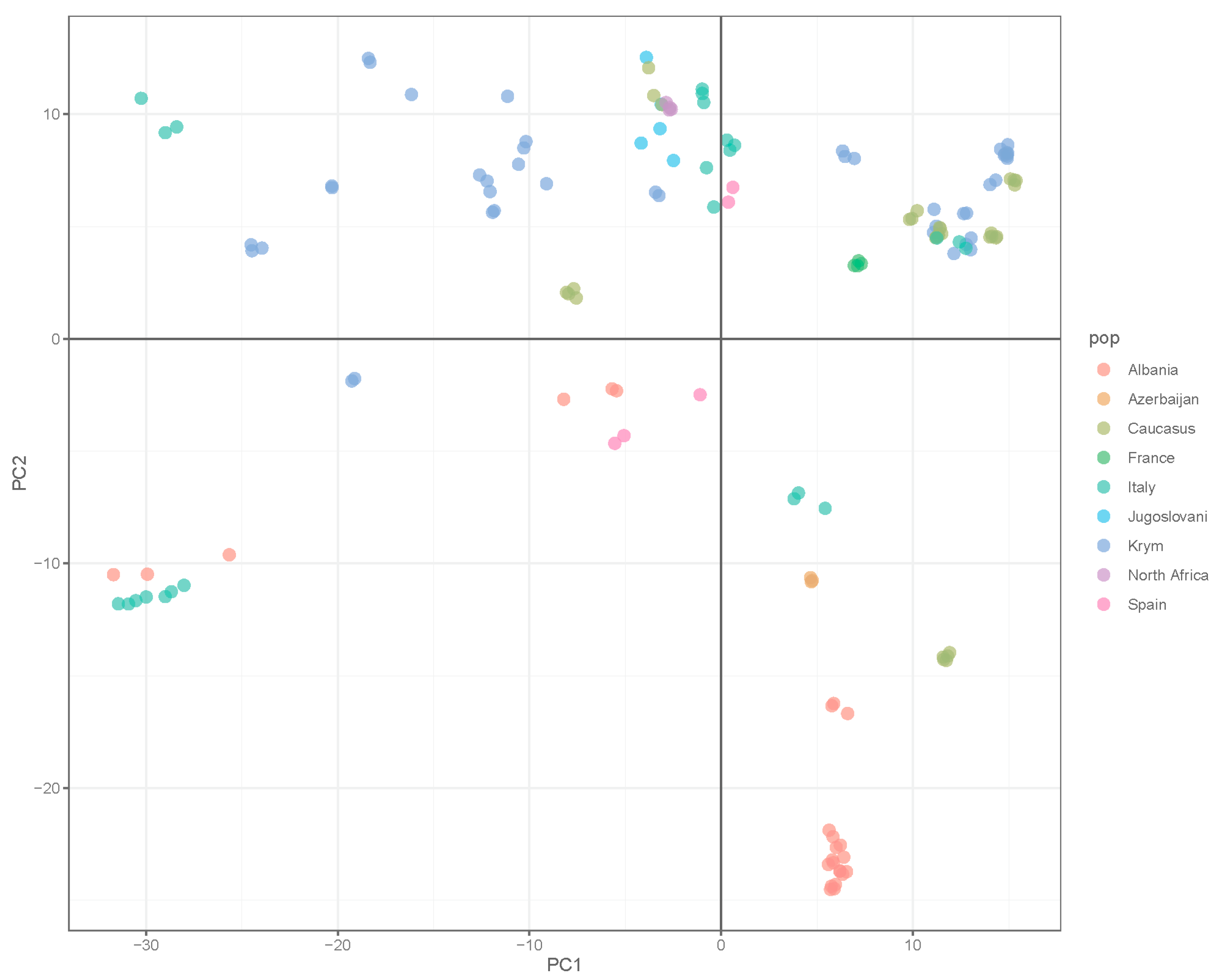
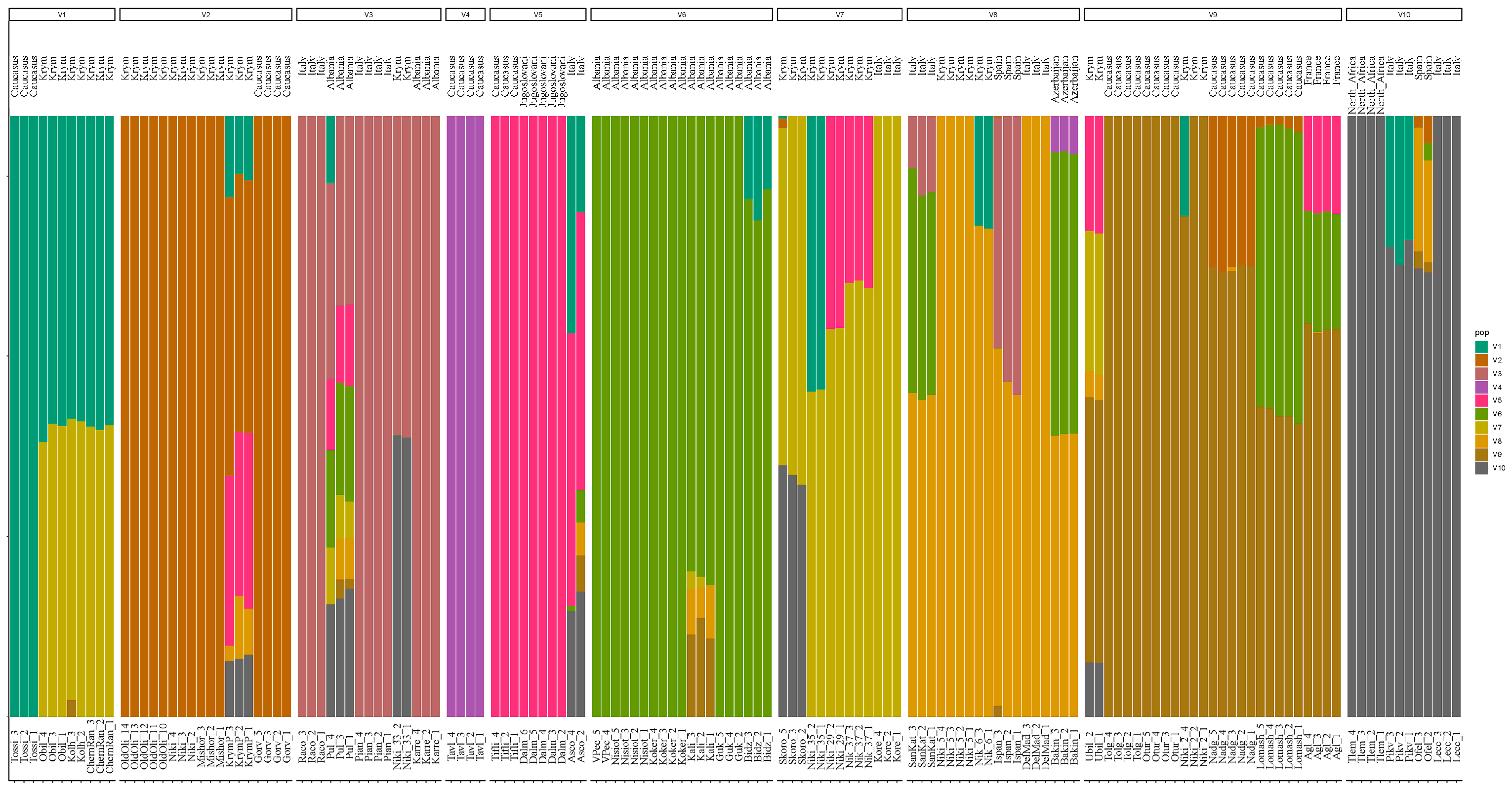
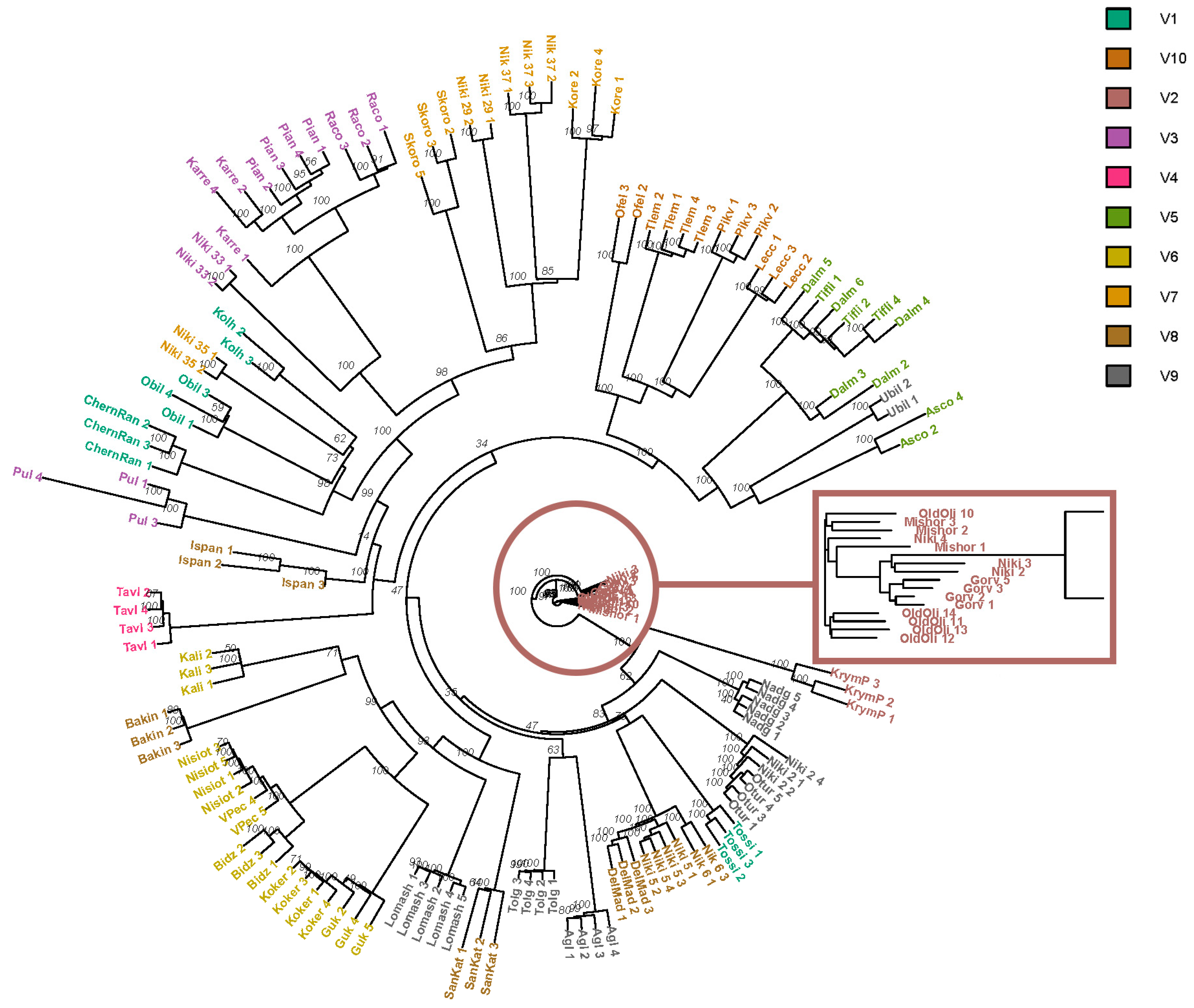
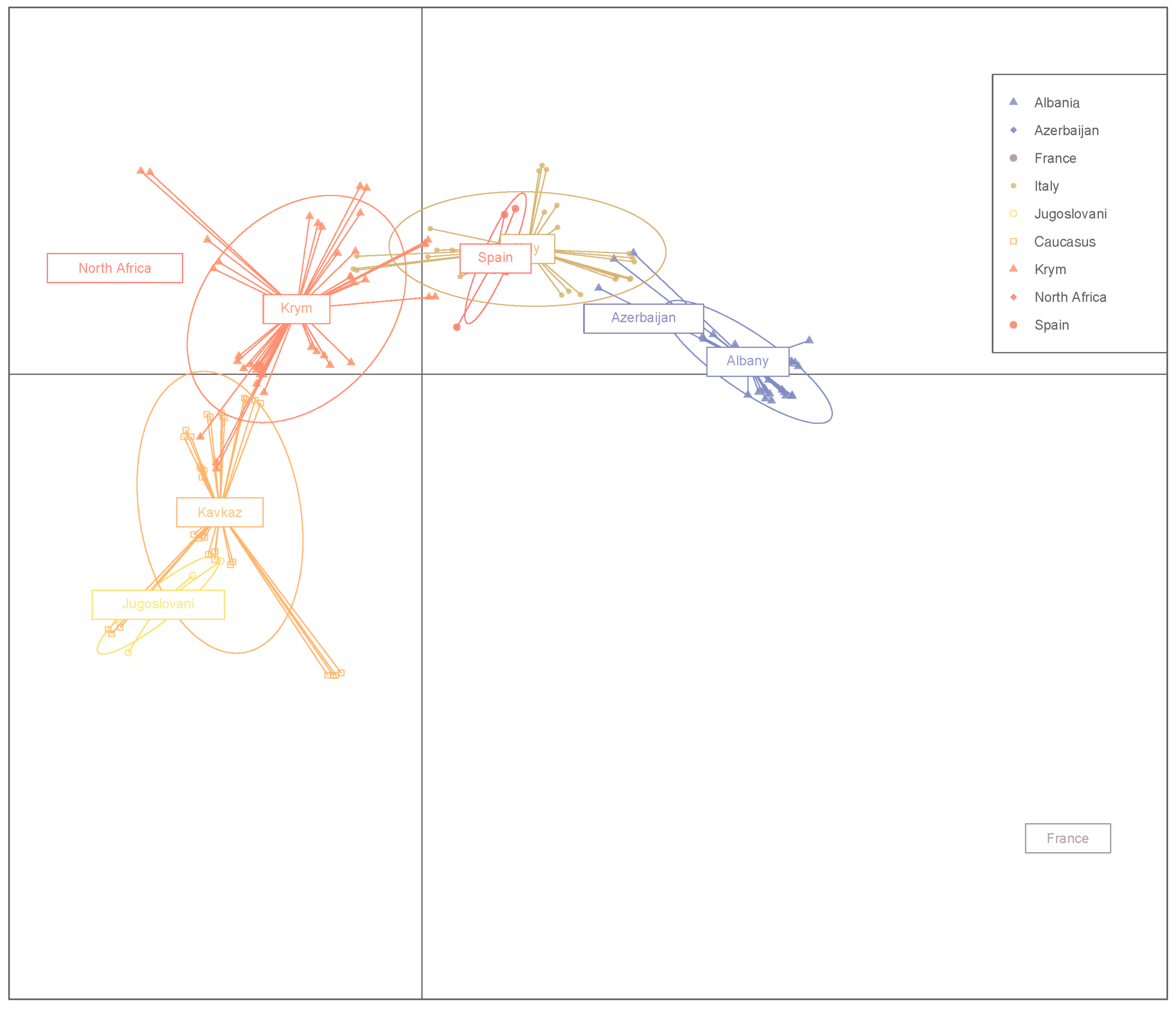
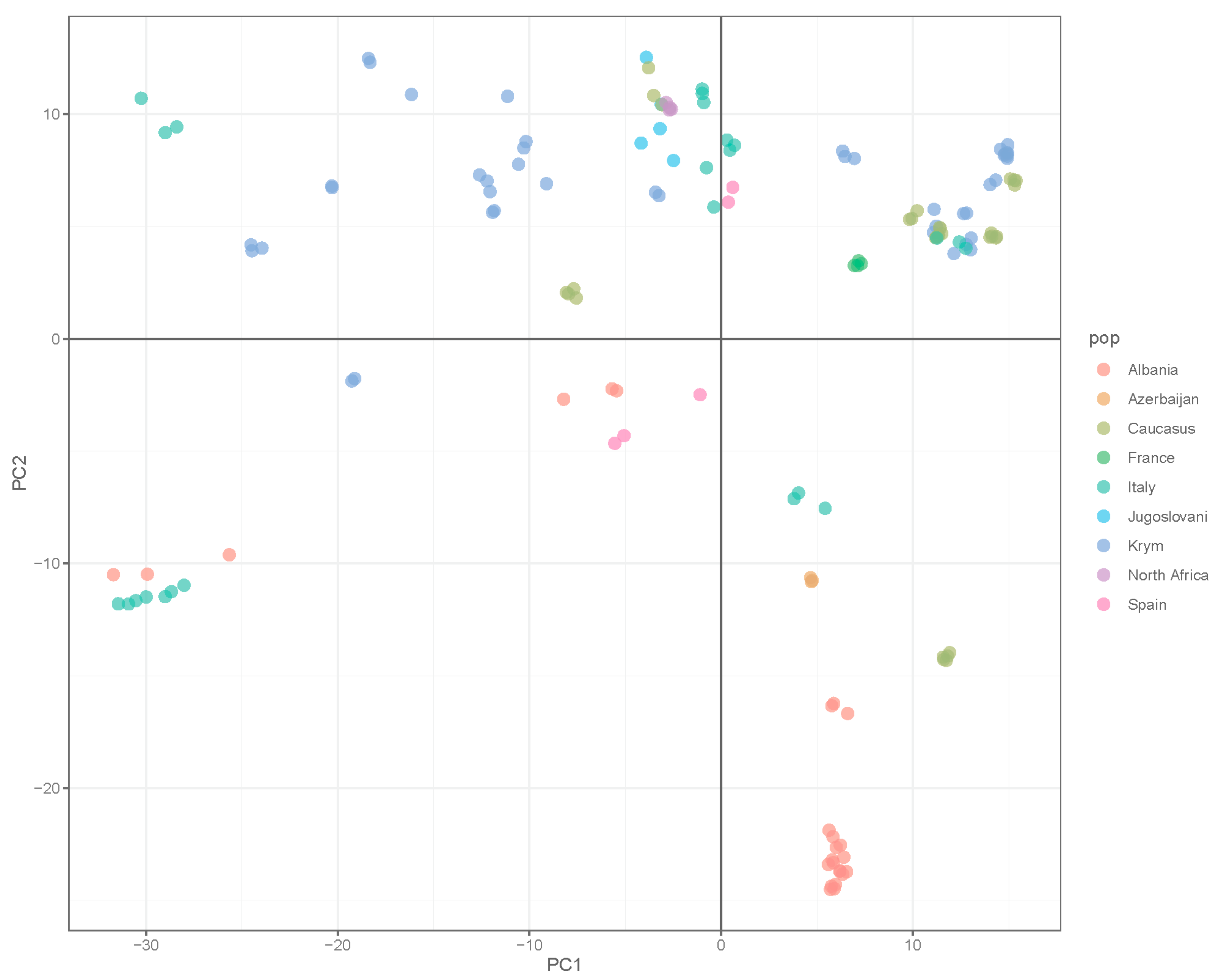
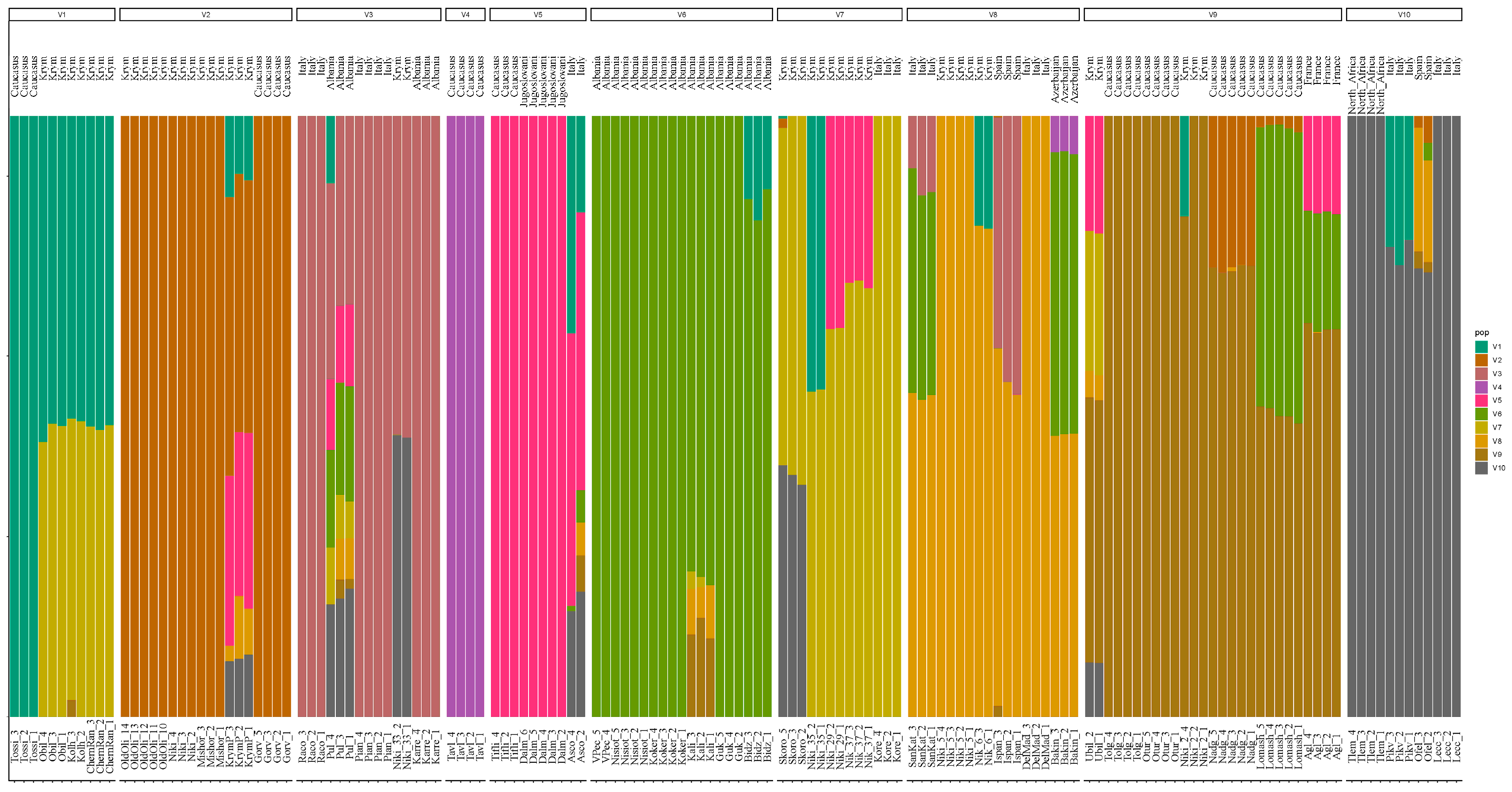
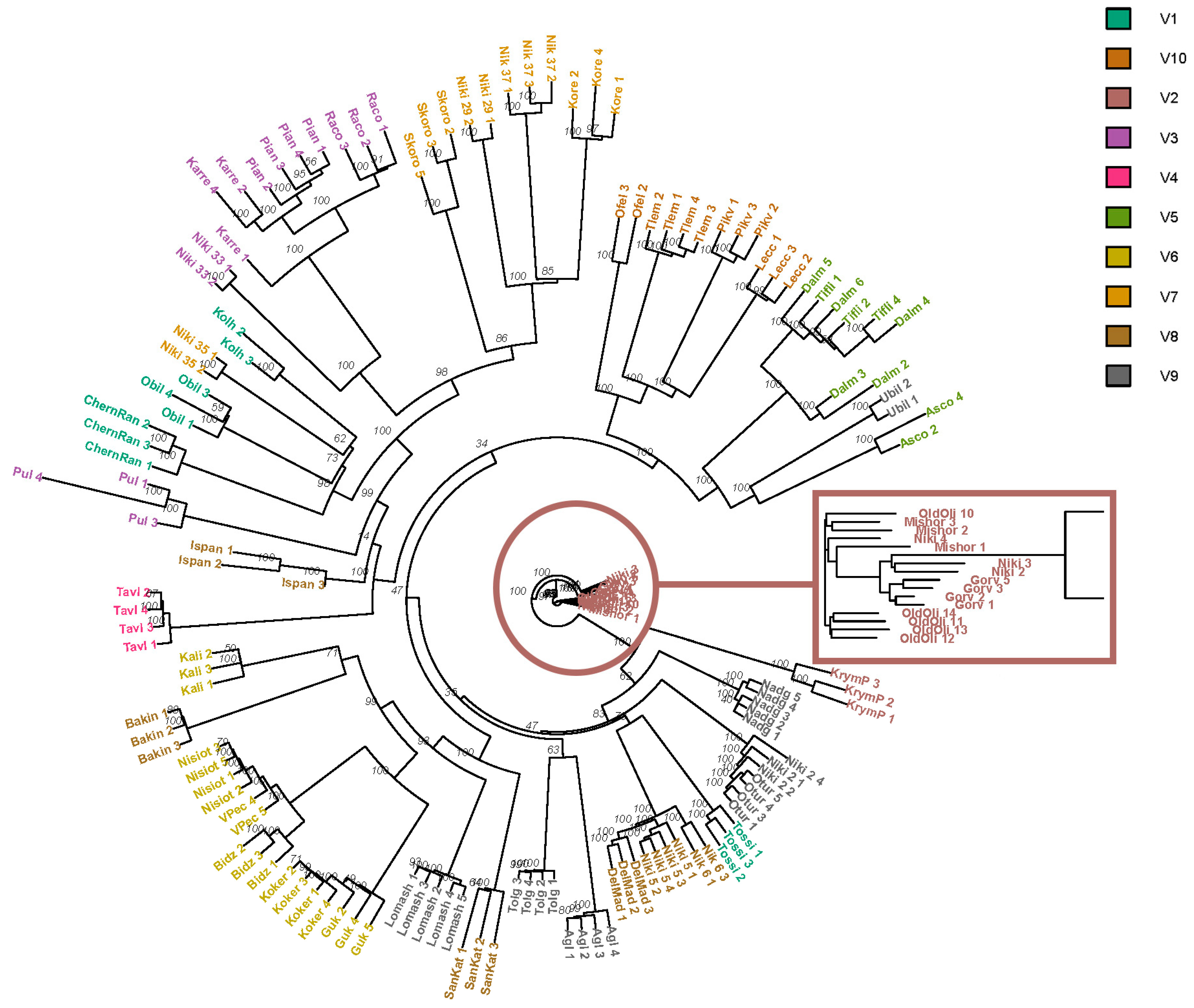
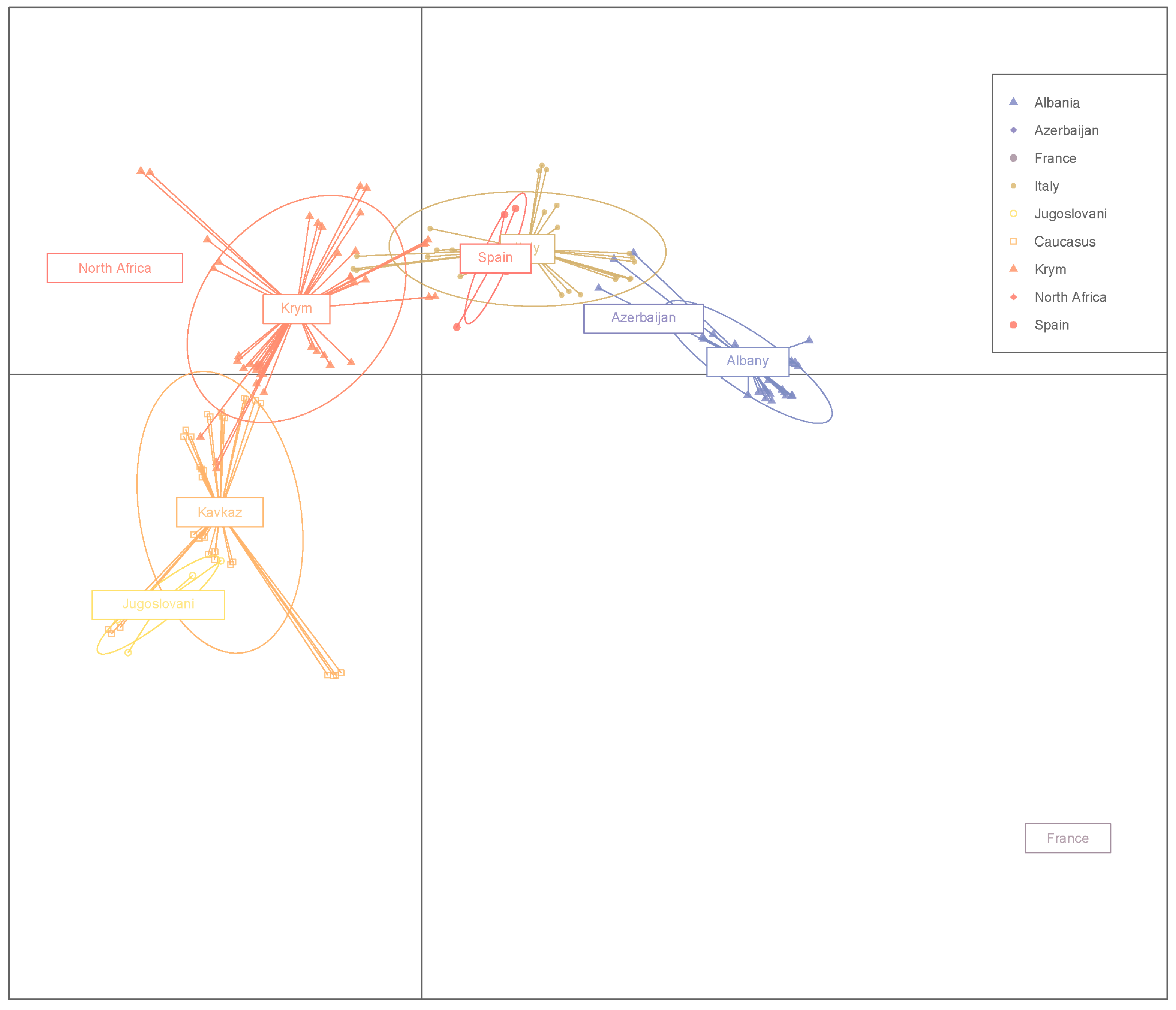
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaniewski, D.; Van Campo, E.; Boiy, T.; Terral, J.-F.; Khadari, B.; Besnard, G. Primary Domestication and Early Uses of the Emblematic Olive Tree: Palaeobotanical, Historical and Molecular Evidence from the Middle East. Biol. Rev. 2012, 87, 885–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langgut, D.; Garfinkel, Y. 7000-Year-Old Evidence of Fruit Tree Cultivation in the Jordan Valley, Israel. Sci. Rep. 2022, 12, 7463. [Google Scholar] [CrossRef] [PubMed]
- Barazani, O.; Dag, A.; Dunseth, Z. The History of Olive Cultivation in the Southern Levant. Front. Plant Sci. 2023, 14, 1131557. [Google Scholar] [CrossRef] [PubMed]
- Breton, C.; Terral, J.-F.; Pinatel, C.; Médail, F.; Bonhomme, F.; Bervillé, A. The Origins of the Domestication of the Olive Tree. Comptes Rendus Biol. 2009, 332, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Besnard, G.; Khadari, B.; Navascués, M.; Fernández-Mazuecos, M.; El Bakkali, A.; Arrigo, N.; Baali-Cherif, D.; Brunini-Bronzini de Caraffa, V.; Santoni, S.; Vargas, P.; et al. The Complex History of the Olive Tree: From Late Quaternary Diversification of Mediterranean Lineages to Primary Domestication in the Northern Levant. Proc. R. Soc. B Biol. Sci. 2013, 280, 2012–2833. [Google Scholar] [CrossRef] [Green Version]
- Diez, C.M.; Trujillo, I.; Martinez-Urdiroz, N.; Barranco, D.; Rallo, L.; Marfil, P.; Gaut, B.S. Olive Domestication and Diversification in the Mediterranean Basin. New Phytol. 2015, 206, 436–447. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations Statistical Dataset Food and Agriculture Data 2021. Available online: https://www.fao.org/faostat (accessed on 24 March 2023).
- Kaya, H.B.; Akdemir, D.; Lozano, R.; Cetin, O.; Sozer Kaya, H.; Sahin, M.; Smith, J.L.; Tanyolac, B.; Jannink, J.-L. Genome Wide Association Study of 5 Agronomic Traits in Olive (Olea europaea L.). Sci. Rep. 2019, 9, 18764. [Google Scholar] [CrossRef] [Green Version]
- Sygouni, V.; Pantziaros, A.G.; Iakovides, I.C.; Sfetsa, E.; Bogdou, P.I.; Christoforou, E.A.; Paraskeva, C.A. Treatment of Two-Phase Olive Mill Wastewater and Recovery of Phenolic Compounds Using Membrane Technology. Membranes 2019, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- The International Olive Council. Available online: https://www.internationaloliveoil.org/ (accessed on 24 March 2023).
- Petruccelli, R.; Bartolini, G.; Ganino, T.; Zelasco, S.; Lombardo, L.; Perri, E.; Durante, M.; Bernardi, R. Cold Stress, Freezing Adaptation, Varietal Susceptibility of Olea europaea L.: A Review. Plants 2022, 11, 1367. [Google Scholar] [CrossRef]
- OLEA Databases; FAO/Plant Production and Protection: Rome, Italy. 2005. Available online: www.oleadb.it (accessed on 24 February 2022).
- Ghanbari, R.; Anwar, F.; Alkharfy, K.M.; Gilani, A.-H.; Saari, N. Valuable Nutrients and Functional Bioactives in Different Parts of Olive (Olea europaea L.)—A Review. Int. J. Mol. Sci. 2012, 13, 3291–3340. [Google Scholar] [CrossRef]
- Duran, S.T.; Aghayeva, S.; Akparov, Z.; Mammadov, A.; Asgarova, R.; Uslu, O.Y.; Kirikoglu, O.; Duran, U.T.; Ipek, M.; Barut, E.; et al. Genetic Variation and Relationships between Azerbaijani and Turkish Olive Genetic Resources. Mol. Biol. Rep. 2022, 49, 5209–5217. [Google Scholar] [CrossRef]
- Bartolini, G.; Petruccelli, R. Classification, Origin, Diffusion and History of the Olive; Food and Agriculture Organization of the United Nations: Rome, Italy, 2002. [Google Scholar]
- Fiorino, P.; Lombardo, N.; Marone, E. Il germoplasma olivicolo: Un patrimonio da valorizzare. Italus Hortus 2005, 12, 7–18. [Google Scholar]
- Barranco Navero, D.; Cimato, A.; Fiorino, P.; Rallo Romero, L.; Touzani, A.; Castañeda, C.; Serafini, E.; Trujillo Navas, I. World Catalogue of Olive Varieties; Mundi-Prensa: Madrid, Spain, 2000. [Google Scholar]
- Cantini, C.; Cimato, A.; Autino, A.; Redi, A.; Cresti, M. Assessment of the Tuscan Olive Germplasm by Microsatellite Markers Reveals Genetic Identities and Different Discrimination Capacity among and within Cultivars. J. Am. Soc. Hortic. Sci. 2008, 133, 598–604. [Google Scholar] [CrossRef] [Green Version]
- Corrado, G.; La Mura, M.; Ambrosino, O.; Pugliano, G.; Varricchio, P.; Rao, R. Relationships of Campanian Olive Cultivars: Comparative Analysis of Molecular and Phenotypic Data. Genome 2009, 52, 692–700. [Google Scholar] [CrossRef] [Green Version]
- Díez, C.M.; Imperato, A.; Rallo, L.; Barranco, D.; Trujillo, I. Worldwide Core Collection of Olive Cultivars Based on Simple Sequence Repeat and Morphological Markers. Crop Sci. 2012, 52, 211–221. [Google Scholar] [CrossRef]
- Trujillo, I.; Ojeda, M.A.; Urdiroz, N.M.; Potter, D.; Barranco, D.; Rallo, L.; Diez, C.M. Identification of the Worldwide Olive Germplasm Bank of Córdoba (Spain) Using SSR and Morphological Markers. Tree Genet. Genomes 2014, 10, 141–155. [Google Scholar] [CrossRef]
- Belaj, A.; Ninot, A.; Gómez-Gálvez, F.J.; El Riachy, M.; Gurbuz-Veral, M.; Torres, M.; Lazaj, A.; Klepo, T.; Paz, S.; Ugarte, J.; et al. Utility of EST-SNP Markers for Improving Management and Use of Olive Genetic Resources: A Case Study at the Worldwide Olive Germplasm Bank of Córdoba. Plants 2022, 11, 921. [Google Scholar] [CrossRef]
- Valeri, M.C.; Mifsud, D.; Sammut, C.; Pandolfi, S.; Lilli, E.; Bufacchi, M.; Stanzione, V.; Passeri, V.; Baldoni, L.; Mariotti, R.; et al. Exploring Olive Genetic Diversity in the Maltese Islands. Sustainability 2022, 14, 10684. [Google Scholar] [CrossRef]
- Said, E.M.; Hassan, M.E. DNA Barcodes in Egyptian Olive Cultivars (Olea europaea L.) Using the RbcL and MatK Coding Sequences. J. Crop Sci. Biotechnol. 2023. [Google Scholar] [CrossRef]
- Debbabi, O.S.; Amar, F.B.; Rahmani, S.M.; Taranto, F.; Montemurro, C.; Miazzi, M.M. The Status of Genetic Resources and Olive Breeding in Tunisia. Plants 2022, 11, 1759. [Google Scholar] [CrossRef]
- Khabou, W.; Ben Amar, F.; Gharsallaoui, M.; Ayadi, M. Agronomic and technological performances of olive tree varieties (Olea europaea L.) in collection in the region of Sfax (Tunisia). In Proceedings of the Second International Seminar Olivebioteq, Marsala, Italy, 5–10 November 2006; Campo Artigrafiche: Alcamo, Italy, 2006; Volume 1, pp. 433–436. [Google Scholar]
- Laaribi, I.; Gouta, H.; Mezghani Ayachi, M.; Labidi, F.; Mars, M. Combination of Morphological and Molecular Markers for the Characterization of Ancient Native Olive Accessions in Central-Eastern Tunisia. Comptes Rendus Biol. 2017, 340, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Batrinou, A.; Strati, I.F.; Houhoula, D.; Tsaknis, J.; Sinanoglou, V.J. Authentication of Olive Oil Based on DNA Analysis. Grasas Aceites 2020, 71, 366. [Google Scholar] [CrossRef]
- Grati-Kamoun, N.; Mahmoud, F.L.; Rebaï, A.; Gargouri, A.; Panaud, O.; Saar, A. Genetic Diversity of Tunisian Olive Tree (Olea europaea L.) Cultivars Assessed by AFLP Markers. Genet. Resour. Crop Evol. 2006, 53, 265–275. [Google Scholar] [CrossRef]
- Ben Ali, S.B.; Mohamed, M.B.; Guasmi, F.; Triki, T.; Kammoun, N.G.; Kamel, N. Genetic Variability of Tunisian Olive Cultivars by ISSR and SSR Markers. J. New Sci. Agric. Biotechnol. 2015, 24, 1098–1107. [Google Scholar]
- Rekik, I.; Salimonti, A.; GratiKamoun, N.; Muzzalupo, I.; Lepais, O.; Gerber, S.; Perri, E.; Rebai, A. Characterization and Identification of Tunisian Olive Tree Varieties by Microsatellite Markers. HortScience 2008, 43, 1371–1376. [Google Scholar]
- Fendri, M.; Trujillo, I.; Trigui, A.; Rodríguez-García, M.I.; de Dios Alché Ramírez, J. Simple Sequence Repeat Identification and Endocarp Characterization of Olive Tree Accessions in a Tunisian Germplasm Collection. HortScience 2010, 45, 1429–1436. [Google Scholar] [CrossRef] [Green Version]
- Ben Mohamed, M.; Zelasco, S.; Ben Ali, S.; Guasmi, F.; Triki, T.; Conforti, F.L.; Naziha, K. Research Article Exploring Olive Trees Genetic Variability in the South East of Tunisia. Genet. Mol. Res. 2017, 16, 16039850. [Google Scholar] [CrossRef]
- Abdelhamid, S.; Omri, A.; Grati-Kamoun, N.; Paolo Marra, F.; Caruso, T. Molecular Characterization and Genetic Relationships of Cultivated Tunisian Olive Varieties (Olea europaea L.) Using SSR Markers. J. New Sci. Agric. Biotechnol. 2017, 40, 2175–2185. [Google Scholar]
- Ben Ayed, R.; Kallel, I.; Ben Hassen, H.; Rebai, A. SNP Marker Analysis for Validating the Authenticity of Tunisian Olive Oil. J. Genet. 2015, 94, 148–154. [Google Scholar] [CrossRef]
- Ben Ayed, R.; Rebai, A. Tunisian Table Olive Oil Traceability and Quality Using SNP Genotyping and Bioinformatics Tools. BioMed Res. Int. 2019, 2019, 8291341. [Google Scholar] [CrossRef]
- Antyufeev, V. Agroclimatic potencial of subtropical gardenic in Crimea. Proc. Orenbg. State Agrar. Univ. 2015, 4, 185–188. [Google Scholar]
- Wulf, E.; Maleev, V. Trees and Shrubs; Proc. of the V.M. Molotov State Nikitsky Botanical Gardens: Yalta, Ukraine, 1948; Volume 22, p. 212. [Google Scholar]
- Franchini, P.; Monné Parera, D.; Kautt, A.F.; Meyer, A. QuaddRAD: A New High-Multiplexing and PCR Duplicate Removal DdRAD Protocol Produces Novel Evolutionary Insights in a Nonradiating Cichlid Lineage. Mol. Ecol. 2017, 26, 2783–2795. [Google Scholar] [CrossRef] [Green Version]
- Korsakova, S.P.; Korsakov, P.B. Climate References of the 2019 Seasons in the Southern Coast of Crimea. Sci. Notes Cape Martyan Nat. Reserv. 2020, 11, 6–22. [Google Scholar] [CrossRef]
- Lo Piccolo, S.; Alfonzo, A.; Conigliaro, G.; Moschetti1, G.; Burruano, S.; Barone, A. A Simple and Rapid DNA Extraction Method from Leaves of Grapevine Suitable for Polymerase Chain Reaction Analysis. AFRICAN J. Biotechnol. 2012, 11, 10305–10309. [Google Scholar] [CrossRef] [Green Version]
- Shalgimbayeva, G.; Volkov, A.; Slobodova, N.; Sharko, F.; Tsygankova, S.; Boulygina, E.; Nguyen, V.Q.; Pham, T.T.; Nguyen, D.T.; Assylbekova, S.Z.; et al. Genetic Investigation of Aral Wild Common Carp Populations (Cyprinus Carpio) Using DdRAD Sequencing. Diversity 2021, 13, 295. [Google Scholar] [CrossRef]
- Rochette, N.C.; Rivera-Colón, A.G.; Catchen, J.M. Stacks 2: Analytical Methods for Paired-end Sequencing Improve RADseq-based Population Genomics. Mol. Ecol. 2019, 28, 4737–4754. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Knaus, B.J.; Grünwald, N.J. vcfr: A Package to Manipulate and Visualize Variant Call Format Data in R. Mol. Ecol. Resour. 2017, 17, 44–53. [Google Scholar] [CrossRef]
- Kamvar, Z.N.; Brooks, J.C.; Granwald, N.J. Novel R Tools for Analysis of Genome-Wide Population Genetic Data with Emphasis on Clonality. Front. Genet. 2015, 6, 208. [Google Scholar] [CrossRef] [Green Version]
- Mijangos, J.L.; Gruber, B.; Berry, O.; Pacioni, C.; Georges, A. DartR v2: An Accessible Genetic Analysis Platform for Conservation, Ecology and Agriculture. Methods Ecol. Evol. 2022, 13, 2150–2158. [Google Scholar] [CrossRef]
- Jombart, T.; Ahmed, I. Adegenet 1.3-1: New Tools for the Analysis of Genome-Wide SNP Data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef] [Green Version]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast Model-Based Estimation of Ancestry in Unrelated Individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [Green Version]
- François, O.; Durand, E. Spatially Explicit Bayesian Clustering Models in Population Genetics. Mol. Ecol. Resour. 2010, 10, 773–784. [Google Scholar] [CrossRef] [PubMed]
- Hartigan, J.A.; Wong, M.A. Algorithm AS 136: A K-Means Clustering Algorithm. Appl. Stat. 1979, 28, 100. [Google Scholar] [CrossRef]
- Sholokhova, V.; Cherkasova, K. Results of Variety Study of Eastern Persimmon and Olive on the Southern Coast of Crimea; Sbornik nauchnykh trudov Gosudarstvennogo Nikitskogo botanicheskogo cada: Kharkiv, Ukraine, 1970; Volume 47. [Google Scholar]
- D’Agostino, N.; Taranto, F.; Camposeo, S.; Mangini, G.; Fanelli, V.; Gadaleta, S.; Miazzi, M.M.; Pavan, S.; di Rienzo, V.; Sabetta, W.; et al. GBS-Derived SNP Catalogue Unveiled Wide Genetic Variability and Geographical Relationships of Italian Olive Cultivars. Sci. Rep. 2018, 8, 15877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sholokhova, V. Catalog of Promising Varieties and Forms of Olives from the Collection of the State Nikitsky Botanical Garden; Proc. of the V.M. Molotov State Nikitsky Botanical Gardens: Yalta, Ukraine, 1973; p. 36. [Google Scholar]
- Sholokhova, V. Guidelines for the Cultivation of Zoned and Promising Varieties and Forms of Olive; Proc. of the V.M. Molotov State Nikitsky Botanical Gardens: Yalta, Ukraine, 1980; p. 30. [Google Scholar]
- Kazas, A.N.; Litvinova, T.V.; Myazina, L.F.; Sinko, L.T.; Khokhlov, S.Y.; Chernobay, I.G.; Shishkina, E.L.; Sholokhova, V.A.; Yadrov, A.A. Subtropical Fruit and Nut Crops: Scientific Reference Edition; IT “ARIAL”: Simferopol, Ukraine, 2012. [Google Scholar]
- Muzzalupo, I.; Vendramin, G.G.; Chiappetta, A. Genetic Biodiversity of Italian Olives (Olea europaea) Germplasm Analyzed by SSR Markers. Sci. World J. 2014, 2014, 296590. [Google Scholar] [CrossRef] [Green Version]
- Las Casas, G.; Scollo, F.; Distefano, G.; Continella, A.; Gentile, A.; La Malfa, S. Molecular Characterization of Olive (Olea europaea L.) Sicilian Cultivars Using SSR Markers. Biochem. Syst. Ecol. 2014, 57, 15–19. [Google Scholar] [CrossRef]
- Richter, A.A. (Ed.) Catalog of Species, Varieties and Hybrid Forms of Subtropical Fruit Crops Growing in the State Nikitsky Botanical Garden; Proc. of the V.M. Molotov State Nikitsky Botanical Gardens: Yalta, Ukraine, 1975; p. 48. [Google Scholar]
- Lauri, P.; Costes, E.; Regnard, J.; Brun, L.; Simon, S.; Monney, P.; Sinoquet, H. Does Knowledge on Fruit Tree Architecture and Its Implications for Orchard Management Improve Horticultural Sustainability? An Overview of Recent Advances in the Apple. ActaHortic 2009, 817, 243–250. [Google Scholar] [CrossRef]
- Cherbiy-Hoffmann, S.U.; Searles, P.S.; Hall, A.J.; Rousseaux, M.C. Influence of Light Environment on Yield Determinants and Components in Large Olive Hedgerows Following Mechanical Pruning in the Subtropics of the Southern Hemisphere. Sci. Hortic. 2012, 137, 36–42. [Google Scholar] [CrossRef]
- Gregoriou, K.; Pontikis, K.; Vemmos, S. Effects of Reduced Irradiance on Leaf Morphology, Photosynthetic Capacity, and Fruit Yield in Olive (Olea europaea L.). Photosynthetica 2007, 45, 172–181. [Google Scholar] [CrossRef]
- Dervishi, A.; Jakše, J.; Ismaili, H.; Javornik, B.; Štajner, N. Genetic Structure and Core Collection of Olive Germplasm from Albania Revealed by Microsatellite Markers. Genes 2021, 12, 256. [Google Scholar] [CrossRef]
- Belaj, A.; Trujillo, I.; de la Rosa, R.; Rallo, L.; Giménez, M.J. Polymorphism and Discrimination Capacity of Randomly Amplified Polymorphic Markers in an Olive Germplasm Bank. J. Am. Soc. Hortic. Sci. 2001, 126, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Besnard, G.; Baradat, P.; Bervillé, A. Genetic Relationships in the Olive (Olea europaea L.) Reflect Multilocal Selection of Cultivars. Theor. Appl. Genet. 2001, 102, 251–258. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
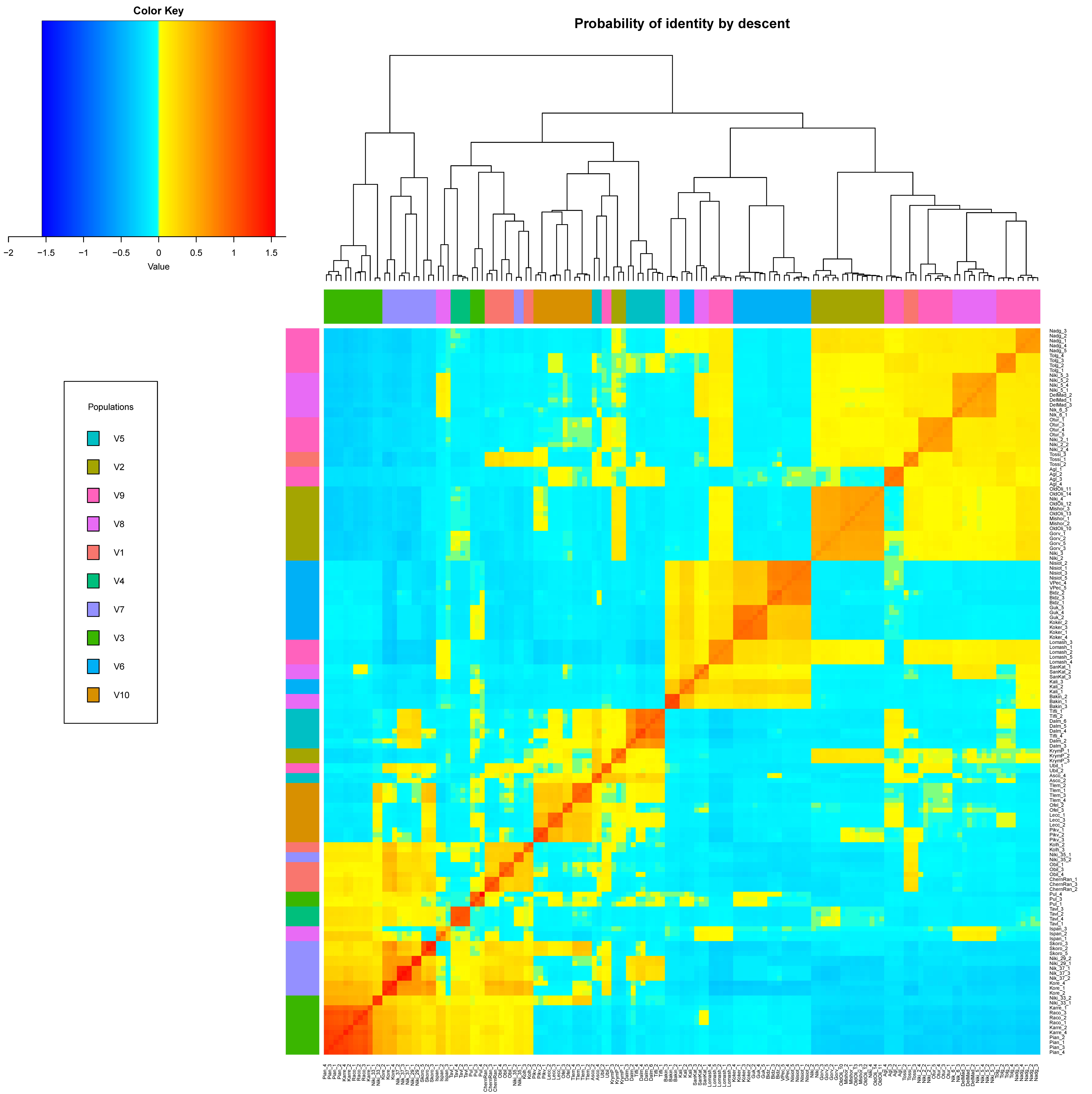{kind=link}
| V5 | V2 | V9 | V8 | V1 | V4 | V7 | V3 | V6 | V10 | |
|---|---|---|---|---|---|---|---|---|---|---|
| V5 | 0 | |||||||||
| V2 | 0.311 | 0 | ||||||||
| V9 | 0.223 | 0.155 | 0 | |||||||
| V8 | 0.237 | 0.196 | 0.09 | 0 | ||||||
| V1 | 0.237 | 0.248 | 0.155 | 0.167 | 0 | |||||
| V4 | 0.374 | 0.372 | 0.285 | 0.295 | 0.303 | 0 | ||||
| V7 | 0.235 | 0.312 | 0.242 | 0.238 | 0.144 | 0.265 | 0 | |||
| V3 | 0.285 | 0.334 | 0.262 | 0.241 | 0.216 | 0.265 | 0.167 | 0 | ||
| V6 | 0.32 | 0.303 | 0.19 | 0.2 | 0.278 | 0.391 | 0.31 | 0.283 | 0 | |
| V10 | 0.229 | 0.253 | 0.188 | 0.202 | 0.205 | 0.314 | 0.209 | 0.253 | 0.278 | 0 |
| Cultivar1 | Cultivar2 | Fst | IBS |
|---|---|---|---|
| Nikitskaya 2 | Otur | 0.011 | 0.974 |
| Nikitskaya 6 | Nikitskaya 5 | 0.005 | 0.973 |
| Nikitskaya 5 | Della Madonna | 0.027 | 0.980 |
| Nikitskaya 6 | Della Madonna | 0.051 | 0.967 |
| Nikitskaya | Miskhorskaya | 0.019 | 0.969 |
| Nikitskaya | Gorvala | 0.045 | 0.983 |
| “Old Olive Tree” | Miskhorskaya | 0.006 | 0.976 |
| “Old Olive Tree” | Gorvala | 0.045 | 0.973 |
| Gorvala | Miskhorskaya | 0.049 | 0.971 |
| Razzo | Piangente | 0.073 | 0.962 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slobodova, N.; Sharko, F.; Gladysheva-Azgari, M.; Petrova, K.; Tsiupka, S.; Tsiupka, V.; Boulygina, E.; Rastorguev, S.; Tsygankova, S. Genetic Diversity of Common Olive (Olea europaea L.) Cultivars from Nikita Botanical Gardens Collection Revealed Using RAD-Seq Method. Genes 2023, 14, 1323. https://doi.org/10.3390/genes14071323
Slobodova N, Sharko F, Gladysheva-Azgari M, Petrova K, Tsiupka S, Tsiupka V, Boulygina E, Rastorguev S, Tsygankova S. Genetic Diversity of Common Olive (Olea europaea L.) Cultivars from Nikita Botanical Gardens Collection Revealed Using RAD-Seq Method. Genes. 2023; 14(7):1323. https://doi.org/10.3390/genes14071323
Chicago/Turabian StyleSlobodova, Natalia, Fedor Sharko, Maria Gladysheva-Azgari, Kristina Petrova, Sergey Tsiupka, Valentina Tsiupka, Eugenia Boulygina, Sergey Rastorguev, and Svetlana Tsygankova. 2023. "Genetic Diversity of Common Olive (Olea europaea L.) Cultivars from Nikita Botanical Gardens Collection Revealed Using RAD-Seq Method" Genes 14, no. 7: 1323. https://doi.org/10.3390/genes14071323
APA StyleSlobodova, N., Sharko, F., Gladysheva-Azgari, M., Petrova, K., Tsiupka, S., Tsiupka, V., Boulygina, E., Rastorguev, S., & Tsygankova, S. (2023). Genetic Diversity of Common Olive (Olea europaea L.) Cultivars from Nikita Botanical Gardens Collection Revealed Using RAD-Seq Method. Genes, 14(7), 1323. https://doi.org/10.3390/genes14071323