
Isoflurane Anesthesia’s Impact on Gene Expression Patterns of Rat Brains in an Ischemic Stroke Model

 , ,
, ,  and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. tMCAO Model in Rats
2.2.1. tMCAO under Long-Term Isoflurane (ISO) Anesthesia
2.2.2. tMCAO Model in Rats under Short-Term ISO Anesthesia
2.2.3. Magnetic Resonance Imaging (MRI)
2.3. RNA Isolation
2.4. cDNA Synthesis and Reverse-Transcription Polymerase Chain Reaction (RT–PCR) in Real Time
2.5. Data Analysis and Statistics
2.5.1. Real-Time RT-PCR Data
2.5.2. RNA-Seq Data
2.5.3. Functional Annotation and Cluster Analysis
2.6. Availability of Data and Material
3. Results
3.1. MRI Study of the Ischemic Injury of Rat Brains at 24 h after tMCAO under Long-Term and Short-Term ISO Anesthesia
3.2. RNA-Seq Analysis of the Effect of IR on the mRNA Level in the Subcortical Structures of the IH Related to SO Rats under Long-Term and Short-Term Anesthesia at 24 h after tMCAO
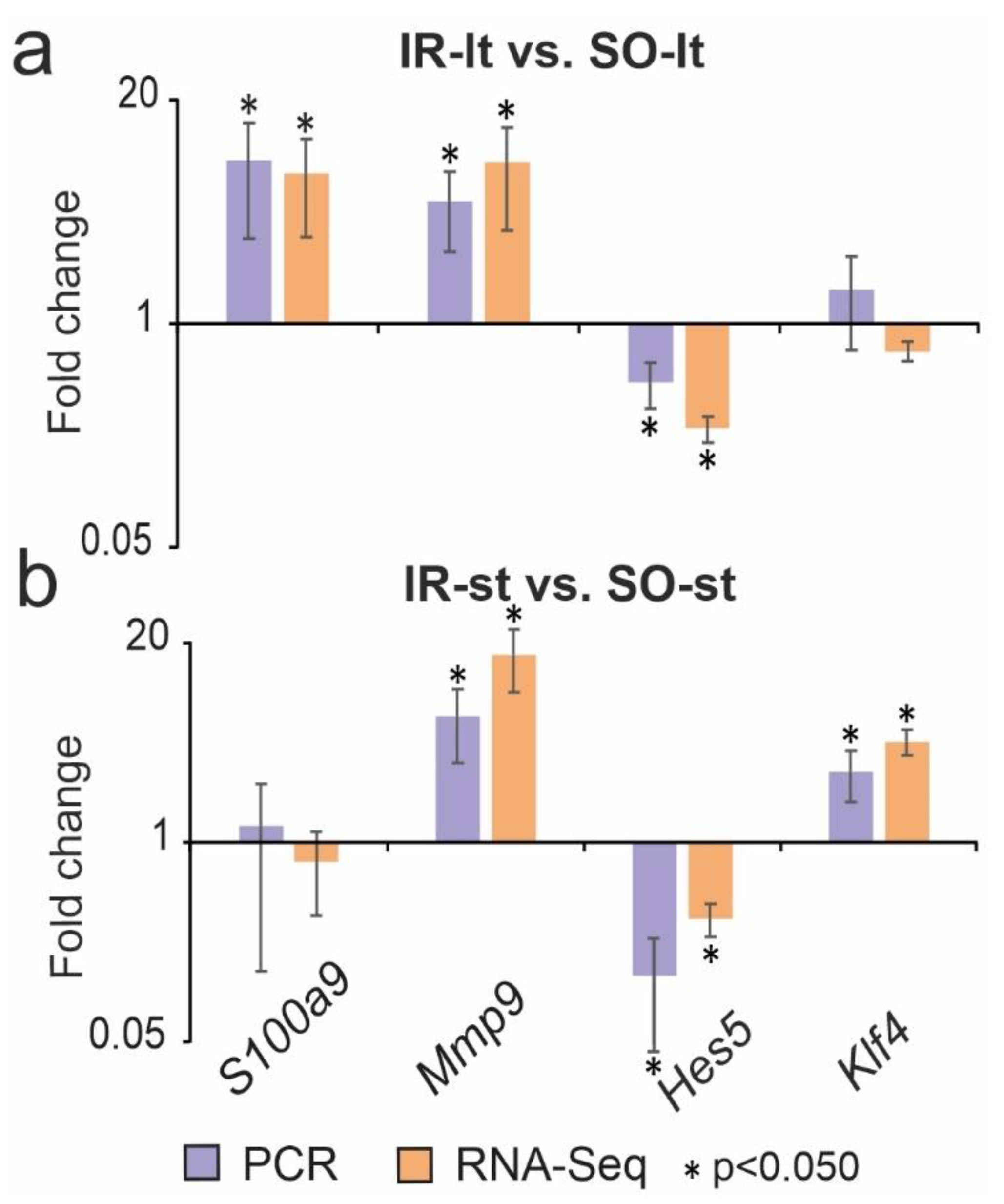
3.3. Reverse-Transcription Polymerase Chain Reaction (RT-PCR) Validation of the RNA-Seq Results at 24 h after tMCAO under Long-Term and Short-Term Anesthesia
3.4. Divergent Changes in the Expression of Genes in Rat Brains under Long-Term and Short-Term Anesthesia 24 h after tMCAO
3.5. Functional Annotation of DEGs in the Subcortical Structures of Brain after tMCAO in Rats under Different Durations of Anesthesia
3.6. Functional Annotation of DEGs That Reversed Their Expression Levels in Rat Brains after tMCAO under Different Durations of Anesthesia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Filippenkov, I.B.; Remizova, J.A.; Denisova, A.E.; Stavchansky, V.V.; Golovina, K.D.; Gubsky, L.V.; Limborska, S.A.; Dergunova, L.V. Comparative Use of Contralateral and Sham-Operated Controls Reveals Traces of a Bilateral Genetic Response in the Rat Brain after Focal Stroke. Int. J. Mol. Sci. 2022, 23, 7308. [Google Scholar] [CrossRef] [PubMed]
- Donkor, E.S. Stroke in the 21st Century: A Snapshot of the Burden, Epidemiology, and Quality of Life. Stroke Res. Treat. 2018, 2018, 30598741. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Diano, M.; Battaglia, S. Editorial: Insights into structural and functional organization of the brain: Evidence from neuroimaging and non-invasive brain stimulation techniques. Front. Psychiatry 2023, 14, 37377471. [Google Scholar] [CrossRef] [PubMed]
- Shimura, A.; Masuya, J.; Yokoi, K.; Morishita, C.; Kikkawa, M.; Nakajima, K.; Chen, C.; Nakagawa, S.; Inoue, T. Too much is too little: Estimating the optimal physical activity level for a healthy mental state. Front. Psychol. 2023, 13, 36710801. [Google Scholar] [CrossRef]
- Polyák, H.; Galla, Z.; Nánási, N.; Cseh, E.K.; Rajda, C.; Veres, G.; Spekker, E.; Szabó, Á.; Klivényi, P.; Tanaka, M.; et al. The Tryptophan-Kynurenine Metabolic System Is Suppressed in Cuprizone-Induced Model of Demyelination Simulating Progressive Multiple Sclerosis. Biomedicines 2023, 11, 945. [Google Scholar] [CrossRef]
- Kuriakose, D.; Xiao, Z. Pathophysiology and treatment of stroke: Present status and future perspectives. Int. J. Mol. Sci. 2020, 21, 7609. [Google Scholar] [CrossRef]
- Durukan, A.; Tatlisumak, T. Acute ischemic stroke: Overview of major experimental rodent models, pathophysiology, and therapy of focal cerebral ischemia. Pharmacol. Biochem. Behav. 2007, 87, 179–197. [Google Scholar] [CrossRef]
- Fluri, F.; Schuhmann, M.K.; Kleinschnitz, C. Animal models of ischemic stroke and their application in clinical research. Drug Des. Devel. Ther. 2015, 9, 3445–3454. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Liu, M.; Pan, Y.; Bai, B.; Chen, J. Global gene expression profile of cerebral ischemia-reperfusion injury in rat MCAO model. Oncotarget 2017, 8, 74607–74622. [Google Scholar] [CrossRef] [Green Version]
- Ford, G.; Xu, Z.; Gates, A.; Jiang, J.; Ford, B.D. Expression Analysis Systematic Explorer (EASE) analysis reveals differential gene expression in permanent and transient focal stroke rat models. Brain Res. 2006, 1071, 226–236. [Google Scholar] [CrossRef]
- Mollet, I.; Marto, J.P.; Mendonça, M.; Baptista, M.V.; Vieira, H.L.A. Remote but not Distant: A Review on Experimental Models and Clinical Trials in Remote Ischemic Conditioning as Potential Therapy in Ischemic Stroke. Mol. Neurobiol. 2022, 59, 294–325. [Google Scholar] [CrossRef]
- Maida, C.D.; Norrito, R.L.; Daidone, M.; Tuttolomondo, A.; Pinto, A. Neuroinflammatory Mechanisms in Ischemic Stroke: Focus on Cardioembolic Stroke, Background, and Therapeutic Approaches. Int. J. Mol. Sci. 2020, 21, 6454. [Google Scholar] [CrossRef]
- Iadecola, C.; Anrather, J. The immunology of stroke: From mechanisms to translation. Nat. Med. 2011, 17, 796. [Google Scholar] [CrossRef]
- Zhu, H.; Zhang, Y.; Zhong, Y.; Ye, Y.; Hu, X.; Gu, L.; Xiong, X. Inflammation-Mediated Angiogenesis in Ischemic Stroke. Front. Cell. Neurosci. 2021, 15, 33967696. [Google Scholar] [CrossRef]
- Peng, L.; Yin, J.; Ge, M.; Wang, S.; Xie, L.; Li, Y.; Si, J.Q.; Ma, K. Isoflurane post-conditioning ameliorates cerebral ischemia/reperfusion injury by enhancing angiogenesis through activating the Shh/Gli signaling pathway in rats. Front. Neurosci. 2019, 13, 31024240. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.N.; Zhang, C.T.; Liu, L.F.; Wu, X.L.; Hu, H.B.; Chen, Y.; Iqbal, M.; Ma, Y.B.; Zhou, J.S.; Xiao, X.L.; et al. Identification of microRNA/target gene in the dentate gyrus of 7-day-old mice following isoflurane exposure. Acta Neurobiol. Exp. 2022, 82, 35451427. [Google Scholar] [CrossRef]
- Zhang, Q.; Yin, J.; Xu, F.; Zhai, J.; Yin, J.; Ge, M.; Zhou, W.; Li, N.; Qin, X.; Li, Y.; et al. Isoflurane post-conditioning contributes to anti-apoptotic effect after cerebral ischaemia in rats through the ERK5/MEF2D signaling pathway. J. Cell. Mol. Med. 2021, 25, 3803–3815. [Google Scholar] [CrossRef]
- Yuan, M.; Ge, M.; Yin, J.; Dai, Z.; Xie, L.; Li, Y.; Liu, X.; Peng, L.; Zhang, G.; Si, J.; et al. Isoflurane post-conditioning down-regulates expression of aquaporin 4 in rats with cerebral ischemia/reperfusion injury and is possibly related to bone morphogenetic protein 4/Smad1/5/8 signaling pathway. Biomed. Pharmacother. 2018, 97, 429–438. [Google Scholar] [CrossRef]
- Koizumi, J.; Yoshida, Y.; Nakazawa, T.; Ooneda, G. Experimental studies of ischemic brain edema. Nosotchu 1986, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Dergunova, L.V.; Filippenkov, I.B.; Stavchansky, V.V.; Denisova, A.E.; Yuzhakov, V.V.; Mozerov, S.A.; Gubsky, L.V.; Limborska, S.A. Genome-wide transcriptome analysis using RNA-Seq reveals a large number of differentially expressed genes in a transient MCAO rat model. BMC Genom. 2018, 19, 655. [Google Scholar] [CrossRef] [Green Version]
- Filippenkov, I.B.; Stavchansky, V.V.; Glazova, N.Y.; Sebentsova, E.A.; Remizova, J.A.; Valieva, L.V.; Levitskaya, N.G.; Myasoedov, N.F.; Limborska, S.A.; Dergunova, L.V. Antistress action of melanocortin derivatives associated with correction of gene expression patterns in the hippocampus of male rats following acute stress. Int. J. Mol. Sci. 2021, 22, 10054. [Google Scholar] [CrossRef] [PubMed]
- Filippenkov, I.B.; Remizova, J.A.; Denisova, A.E.; Stavchansky, V.V.; Golovina, K.D.; Gubsky, L.V.; Limborska, S.A.; Dergunova, L.V. Differential gene expression in the contralateral hemisphere of the rat brain after focal ischemia. Sci. Rep. 2023, 13, 573. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Huang, X.; Muruganujan, A.; Tang, H.; Mills, C.; Kang, D.; Thomas, P.D. PANTHER version 11: Expanded annotation data from Gene Ontology and Reactome pathways, and data analysis tool enhancements. Nucleic Acids Res. 2017, 45, D183–D189. [Google Scholar] [CrossRef] [Green Version]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software Environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/Traces/study/?acc=SRP148632 (accessed on 6 February 2019).
- National Center for Biotechnology Information. Available online: http://www.ncbi.nlm.nih.gov/bioproject/803984 (accessed on 18 April 2022).
- Jiang, M.; Sun, L.; Feng, D.X.; Yu, Z.Q.; Gao, R.; Sun, Y.Z.; Chen, G. Neuroprotection provided by isoflurane pre-conditioning and post-conditioning. Med. Gas Res. 2017, 7, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Ludders, J.W. Advantages and guidelines for using isoflurane. Vet. Clin. North Am.-Small Anim. Pract. 1992, 22, 328–331. [Google Scholar] [CrossRef]
- Li, X.; Li, G.; Jin, Y.; Yao, Q.; Li, R.; Wang, H. Long non-coding RNA maternally expressed 3 (MEG3) regulates isoflurane-induced cognitive dysfunction by targeting miR-7-5p. Toxicol. Mech. Methods 2022, 32, 453–462. [Google Scholar] [CrossRef]
- Bae, S.H.; Yoo, M.R.; Kim, Y.Y.; Hong, I.K.; Kim, M.H.; Lee, S.H.; Kim, D.Y. Brain-derived neurotrophic factor mediates macrophage migration inhibitory factor to protect neurons against oxygen-glucose deprivation. Neural Regen. Res. 2020, 15, 1483–1489. [Google Scholar] [CrossRef]
- Yan, W.; Chen, Z.; Chen, J.; Chen, H. Isoflurane preconditioning protects rat brain from ischemia reperfusion injury via up-regulating the HIF-1α expression through Akt/mTOR/s6K activation. Cell. Mol. Biol. 2016, 62, 38–44. [Google Scholar] [CrossRef]
- Zuo, C.; Ma, J.; Pan, Y.; Zheng, D.; Chen, C.; Ruan, N.; Su, Y.; Nan, H.; Lian, Q.; Lin, H. Isoflurane and Sevoflurane Induce Cognitive Impairment in Neonatal Rats by Inhibiting Neural Stem Cell Development Through Microglial Activation, Neuroinflammation, and Suppression of VEGFR2 Signaling Pathway. Neurotox. Res. 2022, 40, 775–790. [Google Scholar] [CrossRef]
- Demirgan, S.; Akyol, O.; Temel, Z.; Şengelen, A.; Pekmez, M.; Ulaş, O.; Sevdi, M.S.; Erkalp, K.; Selcan, A. Intranasal levosimendan prevents cognitive dysfunction and apoptotic response induced by repeated isoflurane exposure in newborn rats. Naunyn. Schmiedebergs. Arch. Pharmacol. 2021, 394, 1553–1567. [Google Scholar] [CrossRef]
- Yılmaz, H.; Şengelen, A.; Demirgan, S.; Paşaoğlu, H.E.; Çağatay, M.; Erman, İ.E.; Bay, M.; Güneyli, H.C.; Önay-Uçar, E. Acutely increased aquaporin-4 exhibits more potent protective effects in the cortex against single and repeated isoflurane-induced neurotoxicity in the developing rat brain. Toxicol. Mech. Methods 2022, 33, 36127839. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Ye, Y.; Zhu, H.; Zhang, J.; Wang, H.; Lei, J.; Gu, L.; Zhan, L. NLRP3 inflammasome deficiency attenuates cerebral ischemia-reperfusion injury by inhibiting ferroptosis. Brain Res. Bull. 2023, 193, 37–46. [Google Scholar] [CrossRef]
- Lu, Z.; Lu, Q.; Li, Z. Research progress of nuclear factor E 2-related factor 2 signaling pathway in neuroprotective mechanism of cerebral ischemia/reperfusion injury. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2022, 34, 325–328. [Google Scholar] [CrossRef]
- Rosenberg, G.A.; Estrada, E.Y.; Dencoff, J.E. Matrix metalloproteinases and TIMPs are associated with blood-brain barrier opening after reperfusion in rat brain. Stroke 1998, 29, 2189–2195. [Google Scholar] [CrossRef] [Green Version]
- Zhong, C.; Yang, J.; Xu, T.; Xu, T.; Peng, Y.; Wang, A.; Wang, J.; Peng, H.; Li, Q.; Ju, Z.; et al. Serum matrix metalloproteinase-9 levels and prognosis of acute ischemic stroke. Neurology 2017, 89, 805–812. [Google Scholar] [CrossRef]
- Sudarkina, O.Y.; Filippenkov, I.B.; Stavchansky, V.V.; Denisova, A.E.; Yuzhakov, V.V.; Sevan’kaeva, L.E.; Valieva, L.V.; Remizova, J.A.; Dmitrieva, V.G.; Gubsky, L.V.; et al. Brain Protein Expression Profile Confirms the Protective Effect of the ACTH(4–7)PGP Peptide (Semax) in a Rat Model of Cerebral Ischemia–Reperfusion. Int. J. Mol. Sci. 2021, 22, 6179. [Google Scholar] [CrossRef]
- Xu, Z.X.; Xu, L.; Wang, J.Q.; Mang, J.; Yang, L.; He, J.T. Expression changes of the notch signaling pathway of PC12 cells after oxygen glucose deprivation. Int. J. Biol. Macromol. 2018, 118, 1984–1988. [Google Scholar] [CrossRef]
- Yu, H.C.; Qin, H.Y.; He, F.; Wang, L.; Fu, W.; Liu, D.; Guo, F.C.; Liang, L.; Dou, K.F.; Han, H. Canonical notch pathway protects hepatocytes from ischemia/reperfusion injury in mice by repressing reactive oxygen species production through JAK2/STAT3 signaling. Hepatology 2011, 54, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.C.; Bai, L.; Yue, S.Q.; Wang, D.S.; Wang, L.; Han, H.; Dou, K.F. Notch signal protects non-parenchymal cells from ischemia/reperfusion injury in vitro by repressing ROS. Ann. Hepatol. 2013, 12, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Kamat, P.K.; Ahmad, A.S.; Doré, S. Distinctive effect of anesthetics on the effect of limb remote ischemic postconditioning following ischemic stroke. PLoS ONE 2020, 15, 31945776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miró-Mur, F.; Pérez-de-Puig, I.; Ferrer-Ferrer, M.; Urra, X.; Justicia, C.; Chamorro, A.; Planas, A.M. Immature monocytes recruited to the ischemic mouse brain differentiate into macrophages with features of alternative activation. Brain. Behav. Immun. 2016, 53, 18–33. [Google Scholar] [CrossRef] [Green Version]
- Fury, W.; Park, K.W.; Wu, Z.; Kim, E.; Woo, M.S.; Bai, Y.; MacDonald, L.E.; Croll, S.D.; Cho, S. Sustained Increases in Immune Transcripts and Immune Cell Trafficking during the Recovery of Experimental Brain Ischemia. Stroke 2020, 51, 2514–2525. [Google Scholar] [CrossRef]
- Ryckman, C.; Vandal, K.; Rouleau, P.; Talbot, M.; Tessier, P.A. Proinflammatory Activities of S100: Proteins S100A8, S100A9, and S100A8/A9 Induce Neutrophil Chemotaxis and Adhesion. J. Immunol. 2003, 170, 3233–3242. [Google Scholar] [CrossRef] [Green Version]
- Dessing, M.C.; Tammaro, A.; Pulskens, W.P.; Teske, G.J.; Butter, L.M.; Claessen, N.; Van Eijk, M.; Van Der Poll, T.; Vogl, T.; Roth, J.; et al. The calcium-binding protein complex S100A8/A9 has a crucial role in controlling macrophage-mediated renal repair following ischemia/reperfusion. Kidney Int. 2015, 87, 85–94. [Google Scholar] [CrossRef]
- Vandal, K.; Rouleau, P.; Boivin, A.; Ryckman, C.; Talbot, M.; Tessier, P.A. Blockade of S100A8 and S100A9 Suppresses Neutrophil Migration in Response to Lipopolysaccharide. J. Immunol. 2003, 171, 2602–2609. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, L.; Han, Z.; Dong, J.; Pang, D.; Fu, Y.; Li, L. KLF4 alleviates cerebral vascular injury by ameliorating vascular endothelial inflammation and regulating tight junction protein expression following ischemic stroke. J. Neuroinflamm. 2020, 17, 32264912. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.H.; Lee, M.Y.; Yu, J.H.; Kim, M.S.; Song, M.; Seo, C.H.; Kim, H.; Cho, S.R. In situ pluripotency factor expression promotes functional recovery from cerebral ischemia. Mol. Ther. 2016, 24, 1538–1549. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Li, L. The critical role of KLF4 in regulating the activation of A1/A2 reactive astrocytes following ischemic stroke. J. Neuroinflamm. 2023, 20, 36823628. [Google Scholar] [CrossRef]
- Brouns, R.; De Deyn, P.P. The complexity of neurobiological processes in acute ischemic stroke. Clin. Neurol. Neurosurg. 2009, 111, 483–495. [Google Scholar] [CrossRef]
- Neumann, J.T.; Cohan, C.H.; Dave, K.R.; Wright, C.B.; Perez-Pinzon, M.A. Global cerebral ischemia: Synaptic and cognitive dysfunction. Curr. Drug Targets 2013, 14, 20–35. [Google Scholar] [CrossRef]
- Murphy, T.H.; Corbett, D. Plasticity during stroke recovery: From synapse to behaviour. Nat. Rev. Neurosci. 2009, 10, 861–872. [Google Scholar] [CrossRef]
- Filippenkov, I.B.; Stavchansky, V.V.; Denisova, A.E.; Yuzhakov, V.V.; Sevan’kaeva, L.E.; Sudarkina, O.Y.; Dmitrieva, V.G.; Gubsky, L.V.; Myasoedov, N.F.; Limborska, S.A.; et al. Novel Insights into the Protective Properties of ACTH (4-7) PGP (Semax) Peptide at the Transcriptome Level Following Cerebral Ischaemia-Reperfusion in Rats. Genes 2020, 11, 681. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, M.; Deng, D. KLF2 protects BV2 microglial cells against oxygen and glucose deprivation injury by modulating BDNF/TrkB pathway. Gene 2020, 735, 144277. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, M.; Zhou, Y.; Li, P. Case Report: A novel mutation in WFS1 gene (c.1756G>A p.A586T) is responsible for early clinical features of cognitive impairment and recurrent ischemic stroke. Front. Genet. 2023, 14, 36816038. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shpetko, Y.Y.; Filippenkov, I.B.; Denisova, A.E.; Stavchansky, V.V.; Gubsky, L.V.; Limborska, S.A.; Dergunova, L.V. Isoflurane Anesthesia’s Impact on Gene Expression Patterns of Rat Brains in an Ischemic Stroke Model. Genes 2023, 14, 1448. https://doi.org/10.3390/genes14071448
Shpetko YY, Filippenkov IB, Denisova AE, Stavchansky VV, Gubsky LV, Limborska SA, Dergunova LV. Isoflurane Anesthesia’s Impact on Gene Expression Patterns of Rat Brains in an Ischemic Stroke Model. Genes. 2023; 14(7):1448. https://doi.org/10.3390/genes14071448
Chicago/Turabian StyleShpetko, Yana Y., Ivan B. Filippenkov, Alina E. Denisova, Vasily V. Stavchansky, Leonid V. Gubsky, Svetlana A. Limborska, and Lyudmila V. Dergunova. 2023. "Isoflurane Anesthesia’s Impact on Gene Expression Patterns of Rat Brains in an Ischemic Stroke Model" Genes 14, no. 7: 1448. https://doi.org/10.3390/genes14071448
APA StyleShpetko, Y. Y., Filippenkov, I. B., Denisova, A. E., Stavchansky, V. V., Gubsky, L. V., Limborska, S. A., & Dergunova, L. V. (2023). Isoflurane Anesthesia’s Impact on Gene Expression Patterns of Rat Brains in an Ischemic Stroke Model. Genes, 14(7), 1448. https://doi.org/10.3390/genes14071448