Abstract
Drosophila has been a premier model organism for over a century and many discoveries in flies have furthered our understanding of human disease. Flies have been successfully applied to many aspects of health-based research spanning from behavioural addiction, to dysplasia, to RNA dysregulation and protein misfolding. Recently, Drosophila tissues have been used to study biomolecular condensates and their role in multicellular systems. Identified in a wide range of plant and animal species, biomolecular condensates are dynamic, non-membrane-bound sub-compartments that have been observed and characterised in the cytoplasm and nuclei of many cell types. Condensate biology has exciting research prospects because of their diverse roles within cells, links to disease, and potential for therapeutics. In this review, we will discuss processing bodies (P bodies), a conserved biomolecular condensate, with a particular interest in how Drosophila can be applied to advance our understanding of condensate biogenesis and their role in disease.
1. Introduction
Biomolecular condensates, commonly thought to form by phase separation, are self-organising regions of the cytoplasm or nucleoplasm [1]. Having been observed in a myriad of cell types, condensates are thought to be involved in a wide variety of functions, including DNA replication [2], ribosome formation [3,4], and the post-transcriptional regulation of mRNA [5,6]. Processing bodies (P bodies) are an evolutionarily conserved condensate, first discovered in yeast [7] and subsequently detected in many species, including Arabidopsis [8,9], Caenorhabditis elegans [10,11,12], Drosophila [13,14], mice [15], and humans [16,17]. Lacking a surrounding membrane and composed primarily of RNAs and proteins, P bodies are a type of ribonucleoprotein (RNP) granule (note that the terms condensate and granule are often used interchangeably in the literature). Moreover, as hubs of RNA metabolism, P bodies have been defined by the presence of specific proteins associated with mRNA degradation and translational repression [7,13].
Work in yeast, mammalian cell lines (in cellulo), and in vitro systems have been foundational in our understanding of P bodies, detailing their composition [6,18], principles of assembly and disassembly [19,20], responses to environmental change [21,22], and potential roles in neurodegeneration [23], viral infection [24], and cancer pathogenesis [25]. The next step is to further understand the biological role of P bodies, and this will benefit from experiments using multicellular model systems that are amenable to genetic and physical manipulation, as well as providing a whole organism platform to assess the impact of a disease. Drosophila is particularly well suited to the study of P bodies in vivo (defined in this context as P bodies that exist within a complex cellular system, tissue, or organism) due to their conservation of key proteins and organ systems as well as their ability to model human disease.
2. Drosophila P bodies Have Complex Compositions and Multiple Functions and Are Present in Diverse Tissues
P bodies were first observed in cultured Drosophila Schneider 2 (S2) cells [13,26] and much of what we know relating to the formation, maintenance, and protein content of P bodies in flies comes from this system [13]. Whilst powerful, there are limitations to S2 cells as they behave differently depending on their lineage history and experimental context [27]. P bodies have since been identified and are being studied in a wide range of Drosophila cells and tissues to elucidate their function from the molecular to the organismal level.
In mammalian cultured stem cells, P bodies are generally thought to help balance between maintaining stemness and cell differentiation [28,29]. Similarly, in Drosophila intestinal stem cells, in vivo P bodies have been detected, but these were absent in the differentiated daughter enterocyte cells [30]. By repressing the formation of P bodies, pro-differentiation protein expression is elevated and a loss of the parent stem cells in the intestine is observed [30]. P bodies, in this context, are proposed to prevent differentiation whilst simultaneously keeping these cells primed for this transition.
In contrast, both the germinal stem cells as well as the differentiated cells in the Drosophila testes, the spermatogonia and spermatocytes, contain P bodies [27,31]. The DEAD box helicase maternal expression at 31B (Me31B), a conserved P body protein, has been shown to be critical for the translational repression of nanos mRNA, which prevents the de-differentiation of the spermatogonia. However, the link between the function of Me31B and P bodies has not been explicitly investigated [32]. This finding underlines an important paradox when considering P bodies, or any biomolecular condensate: is the observed role of a condensate protein based on the role of the protein itself or the role of the protein in the context of the condensate?
To date, much of the developmental and organismal-based P body data in Drosophila comes from studies in the egg chamber, containing the oocyte and nurse cells, and early embryo [14]. More broadly, decades of research have identified multiple populations of RNP granules in the oocyte, supporting nurse cells, and embryos which are known to have divergent functions, unique subcellular localisations, and overlapping yet distinct protein compositions [33]. For example, early observations in nurse cells identified ‘sponge bodies’ based on their electron microscopy morphology [34]. Whilst sponge bodies have been classified as discrete granules [35], they are increasingly grouped with the P bodies in the literature due to similarities in protein content and proposed function. This highlights one of the challenges as the field matures, which is to determine what gives a condensate its identity and how that identity is reflected in the naming convention.
2.1. Focus on: P Body Protein Content in Drosophila
Proteins make up a significant proportion of P bodies and this specific subsection will highlight our current understanding of those proteins in Drosophila. To elucidate P body biogenesis in Drosophila, it is critical to understand the entire protein complement of P bodies. While co-immunoprecipitation and interactome analysis of a key P body marker, Me31B, in the egg chamber [36,37] and embryo [36] have been completed, it is challenging to discern which of the proteins identified in these experiments are bona fide P body components. This is due to the promiscuous nature of Me31B; it is present in several different germ granules in the egg chamber [33] and has a significant presence in the dilute cytoplasmic phase [38]. Additionally, technical challenges have prevented the deciphering of the entire protein content of P bodies, which has previously been possible in cell culture by fluorescence-activated particle sorting followed by mass spectrometry [6]. Co-localisation studies in S2 cells and egg chambers have informed our current knowledge of the P body proteins in Drosophila (Table 1). However, this table likely underrepresents the grand total of proteins present in P bodies.

Table 1.
Drosophila P body proteins: their role, localisation to P bodies in S2 cells and/or egg chambers, and the human and budding yeast orthologues; not conserved is abbreviated to NC, (?) denotes uncertainty in the orthologue identity.
Below, we focus on some notable data from Table 1:
- Co-localisation studies in the female germline have shown unique RNA binding proteins present in these P bodies, such as Cup, Oo18 RNA binding protein (Orb), Squid (Sqd), Heterogeneous nuclear ribonucleoprotein at 27C (Hrb27C), Exuperantia (Exu), Ypsilon Schachtel (Yps), Lost, and Bruno 1 (Bru1) [35,40,42].
- The CCR4/Not complex, which is localised to P bodies in yeast, mammalian cells, and S2 cells [7,13,17], appears to localise to different cytoplasmic granules in the egg chamber [14].
- The miRNA machinery is localised to P bodies in S2 cells [13], but this is not the case in the egg chamber or embryos, the miRNA machinery present in GW bodies is separate from the P bodies [44].
Together, this highlights that the P body protein content is not necessarily consistent across cell and tissue types, and this presumably influences the recruitment and regulation of mRNAs.
For in vivo phase separation to occur, the protein content of a granule must be governed by certain rules including: (1) a network of interactions between the proteins must exist that are dense and redundant; (2) a significant proportion of the proteins must be able to bind to RNA specifically or non-specifically; (3) a large proportion of proteins must have low-complexity sequences or high levels of intrinsic disorder to allow for weak, non-specific interactions between proteins and RNA [20]. These three requirements are well met for known Drosophila P body proteins. Rules 1 and 2 are demonstrated for Drosophila P body proteins in Figure 1, and rule 3 was demonstrated by completing disorder predictions across the set of known P body proteins in egg chambers. Over 50% of the residues in the P body proteome are predicted to be disordered, and the fraction of disorder of proteins associated with the P body is over 99.9% more disordered than any possible random sized-matched set of proteins taken from the Drosophila proteome [38].
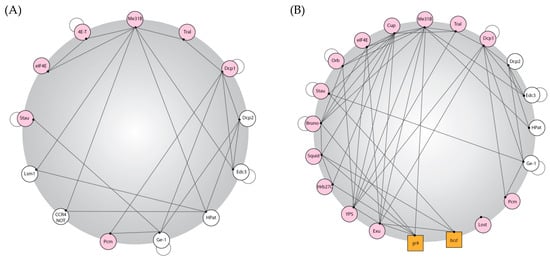
Figure 1.
Drosophila P body protein interactions. P body proteins in Drosophila have dense interaction networks coupled with the ability to bind RNA. Created using publicly available data on BioGRID and based on interaction maps produced for yeast and human P body proteomes [20]. (A,B) An interaction map of Drosophila P body proteins. Proteins are represented in circles and RNA is represented in squares; a line shows potential protein–protein or protein–RNA interactions (these interactions were elucidated using biochemistry; thus, the spatial information, whether this interaction is P-body-specific, is lost, so these diagrams represent maximal/potential interactions that could be taking place in a P body); a pink circle shows that the protein can bind RNA; and an adjoining circle denotes that the protein can self-bind. (A) Interactions between canonical P-body components in Drosophila; (B) interactions between P body components found in Drosophila egg chambers. To consider all possible factors, minimal stringency was applied to the existing data in the field.
Importantly, there is significant tissue specificity in the protein complement of P bodies, and proteins could be recruited in a cell-dependent context to provide specific functions and likely provide additional contacts to increase the interaction density and RNA binding.
2.2. In Vivo P Bodies Can Undergo Regulated Changes
Patient biopsies have shown that P bodies within the same tissue type in humans can also exhibit variation [45], and the Drosophila egg chamber provides a system to test the complexity of such in vivo P bodies. Examples of this diversity include the stoichiometry of resident P body proteins that appears to differ between the oocyte and nurse cells despite these cells being interconnected via cytoplasmic bridges [35,40]. P body RNA content can also be different depending on the subcellular localisation within the oocyte [38,40,42]. Second, immunoelectron microscopy on ultra-thin frozen sections shows that oocyte P bodies have an ultrastructural organisation consisting of an inner core that is devoid of ribosomes and enriched with specific proteins and where non-translating mRNA is stored, and a periphery that is enriched with ribosomes, a translational activator, and an actively translating mRNA [42]. Third, gurken (grk) mRNA translation only occurs at P bodies located at the dorso-anterior corner of the oocyte [40]; this is thought to be due to the post-translational modification of translational activators in P bodies specifically at this sub-cellular location [46].
Developmental and environmental cues can also dramatically influence P body content, form, and function. In the early egg chamber, nurse cell P bodies have been shown to be rapidly and reversibly enlarged upon the addition of environmental stressors [47]. Similar observations have been reported in mammalian and yeast P bodies. Starvation cues originating in the brain of Drosophila drive post-translational modifications and the re-organisation of the microtubule network in egg chambers, which leads to the aggregation of the P bodies [48,49]. This change is hypothesised to allow for the reversible storage and protection of oocyte-specific RNAs until the environmental stress has passed [48].
In mature oocytes, bicoid (bcd) mRNA is localised to the oocyte anterior, where it is similarly stored and translationally repressed in stable P bodies. At this stage, the entirety of the bcd mRNA content is found in P bodies [38,42]; this is likely to ensure complete translational repression until protein expression is required in the embryo for axis patterning. This is in contrast with evidence from mammalian cells that shows of RNAs that localise to P bodies, individual species only have 15–30% of their transcripts in P bodies [50]. This suggests that one possible way that cells can ‘tune’ the expression levels of proteins is by associating different amounts of RNA with P bodies.
In some diseases associated with RNA dysregulation, RNA can become disproportionately segregated into aggregates [51]. There is growing therapeutic interest in the disassembly of these and other aggregates that coincide with pathogenesis. Interestingly, there is a time in Drosophila development when the stable co-localisation between RNA and P bodies dramatically changes, which could help us to understand the properties of condensate disassembly. Egg activation, a universal event that ensures that the oocyte is competent to be fertilised and begin embryogenesis [52], has been shown to result in the disassembly of stable P bodies [38,42,53]. Best supported by data from bcd mRNA, the events of egg activation are hypothesised to release repressed mRNAs that are stored in the core of the P body, thus ensuring the correct spatiotemporal translation [38].
Later, in early embryogenesis, smaller and more dynamic P bodies reform [38], and this offers a unique avenue for the study of de novo condensate formation. Current work shows degradation intermediates for short-lived mRNAs accumulating in reformed P bodies during embryogenesis [54], and P body components Me31B, Decapping protein 1 (DCP1), Staufen (Stau), and Pacman (Pcm) accumulate with oskar (osk) mRNA, and this correlates with the degradation of the mRNA [55]. Additionally, biochemical data suggest that Me31B transitions from a translational repressor to a beacon for mRNA degradation after the maternal to zygotic transition (MZT) [56]. Overall, the ability of Me31B protein and P bodies to change architecture and execute different functions depending on the cellular environment highlights an exciting aspect of condensate biology.
In mammalian and Drosophila neurones, differences in P body components are particularly evident [57,58]. A variety of granules exist, with most containing a limited subset of the canonical P body proteins and a range of specialised components [59,60]. The differences in these neuronal RNP granules have led to inconsistent nomenclature, where granules are sometimes termed P bodies and other times are not. This raises more general questions—what makes a granule a P body and what gives a biomolecular condensate its identity?
Nevertheless, these granules share some functional and material similarities to P bodies in oocytes and embryos and are similarly postulated to act as sites of translational repression until environmental cues lead to their disassembly [61,62]. Neuronal RNP granules also have been found to reform after dispersal [63], similar to P bodies in the early embryo. It is tempting to speculate that there are conserved principles regulating RNP granule assembly and disassembly between dissimilar cell types.
3. Current Understanding of the Requirements for Drosophila P Body Formation
Proteins are critical drivers of phase separation and P body formation. In vitro, protein properties, including valency [64,65], regions of disorder [66,67,68], sequences of low complexity [69,70,71], and the capacity to form weak, non-specific, temporary interactions, have been shown to be relevant to influence phase separation [71,72,73,74,75,76,77]. In mammalian cells, three key P body proteins that have some of these properties, DDX6 (Me31B), Lsm14A (Trailer hitch (Tral)), and eIF4E-Transporter (4E-T/Cup), were required for P body formation under all conditions tested [17,78,79,80], suggesting that these proteins act as scaffolds for P body assembly [19] (scaffolds are broadly defined as proteins and/or RNAs that function to concentrate condensate components [81]). Budding yeast appear more complicated, with Enhancer of decapping 3 (Edc3) and Sm-like protein 4 (LSm4) acting together to form the scaffold [82,83].
As discussed, Drosophila P bodies exist in multiple tissue types and often have specific protein contents. Disruption of P body proteins has revealed differences in the proteins required for P body formation between Drosophila tissues (Table 2).
Table 2.
Available data on the effect that the disruption of canonical P body proteins has on the assembly and size of P bodies in five Drosophila tissues, human immortalised cell lines, and budding yeast. The protein used to visualise the P bodies in the experiment referenced is noted in parentheses.
Below, we highlight some noteworthy data from Table 2.
- HPat and Ge-1 are the only two proteins that appear to be required for the formation of P bodies in more than one cell type in flies [13,30,41]. These could potentially represent the ‘core assembly machinery’ for Drosophila.
- Me31B can act as a scaffold for phase separation in vitro [38], and loss of Me31B results in the disassembly of P bodies in S2 cells [13]. However, in Drosophila nurse cells, when Me31B was mutated, such that it was not able to self-aggregate or be recruited to condensates, Cup and Tral still formed condensates [84]. This suggests that Me31B is not specifically required for condensate formation in this scenario [30,84].
- Tral is not required for P body formation in S2 cells [13], but Tral knockdown leads to smaller P bodies in intestinal stem cells [30], their disassembly in nurse cells [81], and an altered morphology in the mature oocyte [38].
- Edc3 has an inconsistent role in P body formation, with knockdown of Edc3 showing no observable effect in S2 cells [13] but an increased size in intestinal stem cells’ P bodies [30].
- In S2 cells, all members of the miRNA machinery tested were shown to be necessary for P body formation [13], whereas reduction of these factors had no direct consequence on P bodies in intestinal stem cells [30].
It is worth considering why there could be such different outcomes in P body morphology when disrupting a specific protein. Firstly, there may be intrinsic differences in the cellular environment that drive condensate assembly and properties, including protein and RNA composition, concentrations, stoichiometries, binding capacity, and post-translational modifications. In addition, variance could arise from differences in the experimental conditions. These may include the visualisation of P bodies with different marker proteins, different methods used to alter the level of protein expression, and stress conditions introduced by the experimental setup. Additional experiments will be required to verify whether these observed inconsistencies are due to genuine cellular differences.
4. In Vivo Exploration of P Body Biogenesis
The Drosophila egg chamber offers an insightful model to test how phase separation functions in vivo, with many techniques readily available for use in Drosophila (highlighted in Section 4.1). This is illustrated by elegant work on the phase-separated condensates responsible for the transport and translational regulation of osk mRNA, a posterior determinant in Drosophila (note that similar techniques could be applied to Drosophila P bodies in the future). Historically, Bruno 1 (Bru 1) has been shown to bind to sites in the 3′ UTR of osk mRNA and mediate oligomer formation [91], which allows individual mRNPs to self-assemble into higher-order structures capable of moving many mRNAs in a single transport particle [91]. Recently, it was confirmed in vitro that Bru 1 can act as a scaffold for phase separation and that the N terminal domain is critical for this self-assembly process in vivo [92]. Consistent with predictions of what makes a protein a scaffold [93], Bru 1 is modular, with multiple RNA recognition motifs, regions of intrinsic disorder, and several low-complexity domains.
The osk transport particles, also exemplify how the physical nature of the phase-separated granule influences its biological function. These particles have been shown to be ‘solid-like’ and this is required to maintain the RNA within the granule. When the condensate properties were experimentally altered to be more ‘liquid-like’, osk mRNA was prematurely released from the transport particle, resulting in mis-localised translation [92]. Similar experiments in Drosophila would be useful to identify whether the physical nature of the P body is necessary for their biological function in other contexts.
Whilst often overlooked, RNA does appear to have a complex role in the biogenesis of biomolecular condensates [94]. RNA has been shown to be critical in the formation of P bodies in yeast and mammalian cell lines [7,16], and manipulation by RNase A or cycloheximide in Drosophila S2 cells and nurse cells shows that these P bodies are also highly dependent on RNA for their integrity [13,14]. Contrastingly, P bodies are more resistant to a reduction in RNA in oocytes and embryos [14,38,44], suggesting that protein–protein interactions are more important for the structures of these P bodies [38].
Unfortunately, there are only a few well-documented examples of RNAs that associate with P bodies in Drosophila [42,54,95], and more research is needed to fully understand the importance of RNA in P body biogenesis.
Possible ways to identify additional RNAs that are associated with P bodies include particle sorting followed by transcriptomics, which has been successfully implemented in mammalian cells [6] and the more common and lower-throughput approach of assessing co-localisation between P body proteins and RNAs by single-molecule fluorescence in situ hybridisation (smFISH). To find candidates for co-localisation studies, RNAs can be selected based on a low translational status and GC content, particularly in the 3′ UTR [50]. Of note, bcd mRNA when stored in P bodies fits both criteria.
Once RNAs that localise to P bodies have been identified, Drosophila are optimal for the in vivo analysis of the sequence and structural properties of RNA that influence phase separation. This is exemplified by studies of polar granules, a specialised population of RNP granules that are required to specify the fate of the future germ cells in a variety of metazoans [96]. In Drosophila, these granules share significant protein overlap with P bodies [97,98] but, critically, contain ribosomes [99] as well as polar granule specific proteins and RNAs [100]. Over 200 mRNAs are known to be enriched in polar granules [100], and those RNAs tested have been shown to be a highly stable component of the granules [101,102].
However, similarly to oocyte and embryonic P bodies, recent research has shown that it is not the RNA but the proteins that are necessary to regulate the nucleation of the polar granules [103]. Despite this, in vivo RNA, when recruited to an established granule, is capable of self-organisation into higher-order structures [104]. Transgenics and CRISPR approaches have begun to unravel how specific 3′ UTR sequences and structural components in some polar granule RNAs have the ability to drive the recruitment of the RNA to polar granules and then promote the self-assembly of these RNAs within the polar granules [102,105,106,107]. Together, this highlights the critical role that RNA, its sequence, and its structure can play in vivo to organise phase separation. Similar analyses in other condensates in Drosophila will reveal whether these are conserved properties of RNA.
While condensate assembly has received significant research focus, disassembly is now becoming an increasingly popular avenue of research due to the pathological implications and therapeutic potential. Again, the mature oocyte and early embryo provide an attractive in vivo system to explore the molecular mechanism of disassembly.
In the mature oocyte, P bodies have been shown to exist in a stable state, which is thought to be essential for the long-term storage of RNA and sustained translational repression [38]. The precise mechanism that leads to the disassembly of P bodies at egg activation, and the ensuing translation of the released RNA, is not well understood. In vitro studies provide insights that could be applied to articulate the pathways that regulate condensate disassembly at egg activation (Table 3).
Table 3.
A summary of the in vitro mechanisms of the disassembly of condensates and the parallel mechanisms that occur at egg activation.
It seems likely that these mechanisms of disassembly are interconnected in vivo. For example, we know that the calcium rise triggers changes in the post-translational modifications of P body proteins [111], which could occur through PNG, a serine-threonine kinase and the major regulator of post-translational modification at egg activation. In the unactivated oocyte, Gnu (a regulatory subunit of the PNG complex) is localised to P bodies, and, at this point in time, it is unable to bind to and activate the PNG complex [121]. It is tempting to hypothesise that the increase in cytoplasmic calcium, which leads to the activation of Gnu [111,122] and the assembly of the active PNG complex at the P body, could, in turn, disassemble the P body as a result of the phosphorylation of core P body components, the latter of which is supported by strong biochemical evidence [112].
4.1. Focus on: Techniques for the Study of P Bodies in Drosophila
Drosophila enable the combination of genetic power with a wide array of visualisation techniques for proteins and RNAs. This provides an important model system to study the biological relevance of phase separation in vivo and this subsection will cover the available techniques for this investigation.
A wealth of publicly available resources enable the tissue- or cell-specific manipulation of most genes, including their ectopic expression, overexpression, knockdown, and knockout [123,124,125]. Additionally, specific features of a protein or RNA of interest can be altered in particular cells. These methods have been successfully used to study condensates in Drosophila—for example, by altering protein binding sites [27,84]; adding or removing protein regions, namely known aggregation domains or intrinsically disordered regions [92]; changing post-translational modifications using phosphomimetic and non-phosphorylatable forms of a protein [92]; disrupting the RNA sequence [101,102,104]; or altering the RNA secondary structure [102,104].
Advances in microscopy coupled with methods adapted specifically for Drosophila enable the high-resolution visualisation of the P body components and experimentation on the biological effect of altering the protein and RNA in P bodies (Figure 2 and Table 4).
Stem Loop and Coat Protein Binding Systems (MS2-MCP System): The premier method to visualise RNA in living cells is through the insertion of specific secondary structures (stem loops) into the RNA of interest (typically in the 3′ UTR). The second component, the coat protein, is conjugated to a fluorophore. When co-expressed in the same cell, the coat protein can bind to the stem loop with high affinity and thus decorate the mRNA in living cells [126]. This technique (predominantly using the MS2 bacteriophage) has been used to great success in Drosophila [127] and has even been optimised for single-molecule resolution at low laser power [128,129,130].
Single-Molecule Fluorescent In Situ Hybridisation (smFISH): The gold standard for the visualisation of mRNA is smFISH. By creating multiple short nucleotide oligomer probes conjugated to a fluorophore [131], it is possible to label RNA with great specificity and a minimal background. In combination with super-resolution imaging, this method can visualise the sub-granule localisation of single mRNA molecules [38,54,101,102,103,104,105,106].
Multiplexed smFISH: Two sets of smFISH probes can be created for the same RNA species: one allows for the visualisation of the 5′ UTR and the other the 3′ UTR. Full-length RNA and mRNA that have undergone 5′ to 3′ decay will have a different spectral signal, thus enabling decay intermediates to be spatially visualised [54,132]. However, RNAs with short 5′ or 3′ UTRs may be precluded from visualisation with this technique.
Translating RNA Imaging by Coat Knock-Off (TRICK): This system simultaneously utilises the MS2 and PP7 RNA stem loops and their respective coat proteins. In a single RNA species, the PP7 RNA stem loops are engineered in the open reading frame and the MS2 stem loops are placed in the 3′ UTR. When co-expressed with the coat proteins, the untranslated RNA is labelled by both coat protein fluorophores. During the pioneer round of translation, the ribosome(s) knocks off the PP7 coat protein from the RNA but the MS2 coat protein remains bound. In this way, two spectrally different signals appear from untranslated RNA and one signal from translated RNA [133,134]. This has system been successfully implemented in Drosophila [133,134].
SunTag System (a Novel Protein Scaffold, a Repeating Peptide Array [135]): This technique was recently adapted to Drosophila and allows the visualisation of the nascent translation of proteins at the single molecule level. Multiple copies of the GCN4 epitope from yeast are placed at the start of the coding sequence for a protein of interest. Once translated the epitope is recognised by a constitutively expressed cytoplasmic binding partner conjugated to a fluorophore. Multiple copies of the epitope amplify the fluorescent signal, allowing for the visualisation of single molecules of the nascent transcript in real time [129,130,135,136,137,138,139,140,141].

Figure 2.
Examples of methods to investigate RNA in Drosophila. (A) MS2/MCP system: twist mRNA in nuclear cycle 12 and 13 Drosophila embryos, blue represents nuclei not undergoing twist transcription, white represent nuclei undergoing twist transcription and red represents a twist transcription foci, scale bar 10 µm, courtesy of the Lagha Lab [129]. (B) smFISH: primordial germ cell (pgc) and nanos (nos) mRNA visualised here simultaneously with Osk protein during late oogenesis in both the bulk cytoplasm (white box) and germ plasm (yellow box), scale bar 5 µm, courtesy of the Gavis Lab [102]. (C) Multiplexed smFISH: 5′ and 3′ untranslated regions (UTR) of deformed (dfd) mRNA visualised with Me31B protein in the early Drosophila embryo, orange and yellow arrow heads show co-localisation and loss of co-localisation between the 5′ and 3′ ends of the RNA respectively, scale bar 1 µm, courtesy of the Ashe Lab [54]. (D) SunTag system: newly translated Hunchback (Hb) protein visualised simultaneously with hb mRNA, magenta arrow heads show hb mRNA not co-localised Hb protein, green arrow heads show Hb protein not co-localised with hb mRNA and white arrows show a co-localisation between newly translated Hb protein and hb mRNA, scale bar 5 µm, courtesy of the Ashe Lab [140]. Copyright permission has been obtained for all images and there is no conflict of interest.
Figure 2.
Examples of methods to investigate RNA in Drosophila. (A) MS2/MCP system: twist mRNA in nuclear cycle 12 and 13 Drosophila embryos, blue represents nuclei not undergoing twist transcription, white represent nuclei undergoing twist transcription and red represents a twist transcription foci, scale bar 10 µm, courtesy of the Lagha Lab [129]. (B) smFISH: primordial germ cell (pgc) and nanos (nos) mRNA visualised here simultaneously with Osk protein during late oogenesis in both the bulk cytoplasm (white box) and germ plasm (yellow box), scale bar 5 µm, courtesy of the Gavis Lab [102]. (C) Multiplexed smFISH: 5′ and 3′ untranslated regions (UTR) of deformed (dfd) mRNA visualised with Me31B protein in the early Drosophila embryo, orange and yellow arrow heads show co-localisation and loss of co-localisation between the 5′ and 3′ ends of the RNA respectively, scale bar 1 µm, courtesy of the Ashe Lab [54]. (D) SunTag system: newly translated Hunchback (Hb) protein visualised simultaneously with hb mRNA, magenta arrow heads show hb mRNA not co-localised Hb protein, green arrow heads show Hb protein not co-localised with hb mRNA and white arrows show a co-localisation between newly translated Hb protein and hb mRNA, scale bar 5 µm, courtesy of the Ashe Lab [140]. Copyright permission has been obtained for all images and there is no conflict of interest.

Table 4.
A technical summary of the RNA and cellular process visualisation techniques.
Table 4.
A technical summary of the RNA and cellular process visualisation techniques.
| Technique | Information Gained | Live or Fixed | Single- Molecule Resolution Feasible | Multiplexing Possible (Currently) | Signal to Noise Ratio | Potential to Affect RNA Localisation | Super- Resolution Imaging Possible | Adapted to Drosophila |
| Stem loop and coat binding system | RNA localisation [126] | Live [126] | Yes [128] | Yes [128] | Low [126] | Yes [142] | Yes [128] | Yes [127,129,130] |
| smFISH | RNA localisation [131] | Fixed [131] | Yes [131] | Yes [131] | High [131] | No [131] | Yes [54,101,102,103,104,105,106] | Yes [54,101,102,103,104,105,106] |
| Multiplexed smFISH | mRNA decay [132] | Fixed [132] | Yes [132] | Yes [132] | High [132] | No [132] | Yes [132] | Yes [54] |
| TRICK | Nascent translation [133,134] | Live [133,134] | Yes [133,134] | No | Low [133,134] | Yes [133,134] | Yes [133,134] | Yes [133,134] |
| SunTag | Nascent translation [136,137,138,139] | Live [129,136,137,138,139] | Yes [136,137,138,139] | No | High [136,137,138,139] | Low [136,137,138,139] | Yes [136,137,138,139] | Yes [129,140,141] |
5. Using Drosophila as a Model to Understand the Role of P Bodies in Human Diseases
Combining the excellent genetic amenability with the conservation of key organ systems [143,144], Drosophila has a strong history in modelling a myriad of human diseases (see https://www.sdbonline.org/sites/fly/modelsystem/aamodelsystem.htm, accessed on 18 August 2023 for a comprehensive summary, or [145,146,147,148,149,150,151,152,153]). These attributes can also be applied to the study of pathogenesis caused by the dysregulation of RNP granules at a molecular, cellular, tissue, and organismal level. In Drosophila, most research in this area has been focused on stress granules [154,155,156,157,158] as they have more established links to disease phenotypes—for example, in amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD) [159,160,161,162]. However, emerging evidence suggests that P body components are likely implicated in human diseases and Drosophila is well equipped for research in this field.
A major constituent of Lewy bodies, a pathological hallmark of Parkinson’s disease [163], α-synuclein was recently shown to affect the modularity of P bodies in human cells and yeast [23]. Expressing human α-synuclein in Drosophila [164,165,166], it was confirmed that α-synuclein directly interacts with P body proteins and modulates condensate structure [23]. Moreover, the knockdown of various P body proteins in Drosophila modified the α-synuclein-mediated toxicity and locomotor deficiency in adult flies [23].
In patients, a rare missense mutation in DDX6 (a core P body component) leads to intellectual disability, developmental delays, and similar dysmorphic features [167]. Examined in fibroblasts, as neurones are unable to be assayed from patients, P bodies are shown to disassemble when these missense mutations are present, which leads to alterations in translation [167]. This suggests that DDX6 is likely critical for neuronal development but its role in regulating neuronal RNP granules requires additional research. Looking to the future, Drosophila will be a useful model to study these and other neuronal diseases [164,168].
The genetic toolkit of Drosophila allows the straightforward expression of patient-relevant mutations and human disease genes. Mechanistically, changes in aggregation and phase separation can be identified and connected to observable neurodegeneration phenotypes at the individual neurone and whole organism level [154,155,156,157,158,164,165,166]. The screening of genes and small molecules to identify modifiers of disease-relevant mutations is routine in Drosophila and may help to identify novel therapeutics [23,155,156,157,158].
Similar approaches in Drosophila are applicable to other complex human diseases, such as cancer, as they are well suited to experimentation on many of the hallmarks of tumorigenesis. At this time, the use of Drosophila in P body-specific cancer biology research is yet to be realised. What we do have evidence for shows that the link between P bodies and cancer is in its infancy, with the role of P bodies being highly dependent on the cell type and mutational history [25]. Early evidence suggests that P bodies may control changes to the translational landscape that occur during key events in disease progression, such as the epithelial to mesenchymal transition (EMT) [169,170]. Drosophila would be a valuable resource to test the role of P bodies in the EMT during normal development and pathogenesis.
Due to the role of P bodies in RNA metabolism, they appear to be a target for a variety of viruses, which collectively have a devastating impact on human health worldwide [171,172,173,174,175,176,177,178,179,180,181,182]. Typical P body functions are often perturbed as a consequence of viral activity in human cell lines [171], and the modulation of P bodies by viruses is conserved to Drosophila [172,173,174,175]. Viral proteins and RNAs can localise to and interact with P bodies, resulting in their disassembly or aggregation [173,174,175,176,177,178,179,180,181]. In addition viruses can disrupt P bodies by hijacking components for their own replication [182,183,184] or transcription [172].
A significant number of viruses that impact P bodies use arthropods as a vector—for example, the flaviviruses (Dengue virus, West Nile virus) [185], the bunyaviruses (Rift Valley Fever virus) [172,186], and the old world alphaviruses (Chikungunya virus) [187,188]. The interactions between these viruses and cellular processes may be deeply conserved as these insect-borne viruses can replicate in evolutionarily distant hosts [189]. By combining the genetic tractability and conservation of innate immune biology [190], Drosophila could be used to probe for the insect and human anti-viral factors that act in the interplay between P bodies and viruses.
However, the last common ancestor of humans and fruit flies lived over 600 million years ago [191] and there has been obvious divergence in this time. Drosophila lacks the organ system complexity of their human counterparts and data interpretation should reflect this when modelling complex diseases. Despite this, human and Drosophila cells are observed to be strikingly similar in both normal and diseased conditions [145]. Over 75% of human disease genes [192,193] and all known canonical human P body components have functional homologues in Drosophila. Thus, they are particularly well suited to understanding the fundamental mechanisms underpinning intricate human diseases.
6. Concluding Remarks: Drosophila as a Model for the Future Study of P Bodies
Drosophila is a good model to answer many outstanding questions about P bodies, RNP granules, biomolecular condensates, and phase separation in an in vivo context. This is due to the biologically relevant changes that P bodies undergo during development, the conservation of P body proteins, genetic malleability, and the high-resolution imaging techniques to examine RNA and proteins in living and fixed Drosophila tissue. Looking to the future, the following are major questions in the field of condensate biology that we feel Drosophila is particularly well suited to address:
- What regulates the assembly and disassembly of condensates in vivo?
- What effect do proteins, and their specific domains, have on condensate properties?
- What effect do post-translational modifications have on condensate integrity?
- How do RNAs, and certain motifs, contribute to the formation of RNP granules?
- Which RNA structures, sequences, and post-transcriptional modifications affect the ability of RNA to associate with RNP granules?
- Are the material properties of RNP granules intrinsic to their biological function?
- What are the biological functions of phase separation?
Time will tell whether biomolecular condensates fulfil their promise of having a major role in biology and human disease, and Drosophila will be important in this endeavour.
Author Contributions
Conceptualization, E.L.W. and T.T.W.; formal analysis, E.L.W.; writing—original draft preparation, E.L.W.; writing—review and editing, E.L.W. and T.T.W.; project administration, E.L.W. and T.T.W.; funding acquisition, E.L.W. and T.T.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Wellcome Trust (200734/Z/16/Z) (to T.T.W) and a BBSRC DTP scholarship (to E.L.W).
Data Availability Statement
Data sharing not applicable.
Acknowledgments
We are grateful to Elizbeth Gavis, Hilary Ashe and Mounia Lagha for use of their images in Figure 2.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Banani, S.F.; Lee, H.O.; Hyman, A.A.; Rosen, M.K. Biomolecular condensates: Organizers of cellular biochemistry. Nat. Rev. Mol. Cell Biol. 2017, 18, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Parker, M.W.; Bell, M.; Mir, M.; Kao, J.A.; Darzacq, X.; Botchan, M.R.; Berger, J.M. A new class of disordered elements controls DNA replication through initiator self-assembly. eLife 2019, 8, e48562. [Google Scholar] [CrossRef] [PubMed]
- Feric, M.; Vaidya, N.; Harmon, T.S.; Mitrea, D.M.; Zhu, L.; Richardson, T.M.; Kriwacki, R.W.; Pappu, R.V.; Brangwynne, C.P. Coexisting liquid phases underlie nucleolar sub-compartments. Cell 2016, 165, 1686–1697. [Google Scholar] [CrossRef] [PubMed]
- Brangwynne, C.P.; Mitchison, T.J.; Hyman, A.A. Active liquid-like behavior of nucleoli determines their size and shape in Xenopus laevis oocytes. Proc. Natl. Acad. Sci. USA 2011, 108, 4334–4339. [Google Scholar] [CrossRef]
- Horvathova, I.; Voigt, F.; Kotrys, A.V.; Zhan, Y.; Artus-Revel, C.G.; Eglinger, J.; Stadler, M.B.; Giorgetti, L.; Chao, J.A. The Dynamics of mRNA Turnover Revealed by Single-Molecule Imaging in Single Cells. Mol. Cell 2017, 68, 615–625.e9. [Google Scholar] [CrossRef]
- Hubstenberger, A.; Courel, M.; Bénard, M.; Souquere, S.; Ernoult-Lange, M.; Chouaib, R.; Yi, Z.; Morlot, J.-B.; Munier, A.; Fradet, M.; et al. P-Body Purification Reveals the Condensation of Repressed mRNA Regulons. Mol. Cell 2017, 68, 144–157.e5. [Google Scholar] [CrossRef]
- Sheth, U.; Parker, R. Decapping and Decay of Messenger RNA Occur in Cytoplasmic Processing Bodies. Science 2003, 300, 805–808. [Google Scholar] [CrossRef]
- Xu, J.; Chua, N.-H. Arabidopsis decapping 5 is required for mRNA decapping, P-body formation, and translational repression during postembryonic development. Plant Cell 2009, 21, 3270–3279. [Google Scholar] [CrossRef]
- Xu, J.; Yang, J.-Y.; Niu, Q.-W.; Chua, N.-H. Arabidopsis DCP2, DCP1, and VARICOSE Form a Decapping Complex Required for Postembryonic Development. Plant Cell 2006, 18, 3386–3398. [Google Scholar] [CrossRef]
- Gallo, C.M.; Munro, E.; Rasoloson, D.; Merritt, C.; Seydoux, G. Processing bodies and germ granules are distinct RNA granules that interact in C. elegans embryos. Dev. Biol. 2008, 323, 76–87. [Google Scholar] [CrossRef]
- Ding, L.; Spencer, A.; Morita, K.; Han, M. The developmental timing regulator AIN-1 interacts with miRISCs and may target the argonaute protein ALG-1 to cytoplasmic P bodies in C. elegans. Mol. Cell 2005, 19, 437–447. [Google Scholar] [CrossRef]
- Zhang, L.; Ding, L.; Cheung, T.H.; Dong, M.-Q.; Chen, J.; Sewell, A.K.; Liu, X.; Yates, J.R.; Han, M. Systematic identification of miRISC proteins, miRNAs, and their mRNA targets in C. elegans by their interactions with GW182 family proteins AIN-1 and AIN-2. Mol. Cell 2007, 28, 598–613. [Google Scholar] [CrossRef] [PubMed]
- Eulalio, A.; Behm-Ansmant, I.; Schweizer, D.; Izaurralde, E. P-body formation is a consequence, not the cause, of RNA-mediated gene silencing. Mol. Cell Biol. 2007, 27, 3970–3981. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-D.; Jiao, X.; Grima, D.; Newbury, S.F.; Kiledjian, M.; Chou, T.-B. Drosophila processing bodies in oogenesis. Dev. Biol. 2008, 322, 276–288. [Google Scholar] [CrossRef]
- Flemr, M.; Ma, J.; Schultz, R.M.; Svoboda, P. P-Body Loss Is Concomitant with Formation of a Messenger RNA Storage Domain in Mouse Oocytes. Biol. Reprod. 2010, 82, 1008–1017. [Google Scholar] [CrossRef] [PubMed]
- Cougot, N.; Babajko, S.; Séraphin, B. Cytoplasmic foci are sites of mRNA decay in human cells. J. Cell Biol. 2004, 165, 31–40. [Google Scholar] [CrossRef]
- Andrei, M.A.; Ingelfinger, D.; Heintzmann, R.; Achsel, T.; Rivera-Pomar, R.; Lührmann, R. A role for eIF4E and eIF4E-transporter in targeting mRNPs to mammalian processing bodies. RNA 2005, 11, 717–727. [Google Scholar] [CrossRef]
- Xing, W.; Muhlrad, D.; Parker, R.; Rosen, M.K. A quantitative inventory of yeast P body proteins reveals principles of composition and specificity. eLife 2020, 9, e56525. [Google Scholar] [CrossRef]
- Standart, N.; Weil, D. P-Bodies: Cytosolic Droplets for Coordinated mRNA Storage. Trends Genet. 2018, 34, 612–626. [Google Scholar] [CrossRef]
- Luo, Y.; Na, Z.; Slavoff, S.A. P-Bodies: Composition, Properties, and Functions. Biochemistry 2018, 57, 2424–2431. [Google Scholar] [CrossRef]
- Wang, C.; Schmich, F.; Srivatsa, S.; Weidner, J.; Beerenwinkel, N.; Spang, A. Context-dependent deposition and regulation of mRNAs in P-bodies. eLife 2018, 7, e29815. [Google Scholar] [CrossRef] [PubMed]
- Riggs, C.L.; Kedersha, N.; Ivanov, P.; Anderson, P. Mammalian stress granules and P bodies at a glance. J. Cell Sci. 2020, 133, jcs242487. [Google Scholar] [CrossRef] [PubMed]
- Hallacli, E.; Kayatekin, C.; Nazeen, S.; Wang, X.H.; Sheinkopf, Z.; Sathyakumar, S.; Sarkar, S.; Jiang, X.; Dong, X.; Di Maio, R.; et al. The Parkinson’s disease protein α-synuclein is a modulator of processing bodies and mRNA stability. Cell 2022, 185, 2035–2056.e33. [Google Scholar] [CrossRef]
- Boccaccio, G.L.; Thomas, M.G.; García, C.C. Membraneless Organelles and Condensates Orchestrate Innate Immunity Against Viruses. J. Mol. Biol. 2023, 435, 167976. [Google Scholar] [CrossRef] [PubMed]
- Lavalée, M.; Curdy, N.; Laurent, C.; Fournié, J.-J.; Franchini, D.-M. Cancer cell adaptability: Turning ribonucleoprotein granules into targets. Trends Cancer 2021, 7, 902–915. [Google Scholar] [CrossRef] [PubMed]
- Rehwinkel, J.; Behm-Ansmant, I.; Gatfield, D.; Izaurralde, E. A crucial role for GW182 and the DCP1:DCP2 decapping complex in miRNA-mediated gene silencing. RNA 2005, 11, 1640–1647. [Google Scholar] [CrossRef] [PubMed]
- Layana, C.; Vilardo, E.S.; Corujo, G.; Hernández, G.; Rivera-Pomar, R. Drosophila Me31B is a Dual eIF4E-Interacting Protein. J. Mol. Biol. 2023, 435, 167949. [Google Scholar] [CrossRef]
- Di Stefano, B.; Luo, E.-C.; Haggerty, C.; Aigner, S.; Charlton, J.; Brumbaugh, J.; Ji, F.; Rabano Jiménez, I.; Clowers, K.J.; Huebner, A.J.; et al. The RNA Helicase DDX6 Controls Cellular Plasticity by Modulating P-Body Homeostasis. Cell Stem Cell 2019, 25, 622–638.e13. [Google Scholar] [CrossRef]
- Kolaj, A.; Zahr, S.K.; Wang, B.S.; Krawec, T.; Kazan, H.; Yang, G.; Kaplan, D.R.; Miller, F.D. The P-body protein 4E-T represses translation to regulate the balance between cell genesis and establishment of the postnatal NSC pool. Cell Rep. 2023, 42, 112242. [Google Scholar] [CrossRef]
- Buddika, K.; Huang, Y.-T.; Ariyapala, I.S.; Butrum-Griffith, A.; Norrell, S.A.; O’Connor, A.M.; Patel, V.K.; Rector, S.A.; Slovan, M.; Sokolowski, M.; et al. Coordinated repression of pro-differentiation genes via P-bodies and transcription maintains Drosophila intestinal stem cell identity. Curr. Biol. 2022, 32, 386–397.e6. [Google Scholar] [CrossRef]
- Zabolotskaya, M.V.; Grima, D.P.; Lin, M.-D.; Chou, T.-B.; Newbury, S.F. The 5′-3′ exoribonuclease Pacman is required for normal male fertility and is dynamically localized in cytoplasmic particles in Drosophila testis cells. Biochem. J. 2008, 416, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.; Venkei, Z.G.; Watase, G.J.; Bisai, B.; Pletcher, S.; Lee, C.-Y.; Yamashita, Y.M. me31B regulates stem cell homeostasis by preventing excess dedifferentiation in the Drosophila male germline. J. Cell Sci. 2021, 134, jcs258757. [Google Scholar] [CrossRef] [PubMed]
- Sankaranarayanan, M.; Weil, T.T. Granule regulation by phase separation during Drosophila oogenesis. Emerg. Top. Life Sci. 2020, 4, 355–364. [Google Scholar]
- Wilsch-Bräuninger, M.; Schwarz, H.; Nüsslein-Volhard, C. A Sponge-like Structure Involved in the Association and Transport of Maternal Products during Drosophila Oogenesis. J. Cell Biol. 1997, 139, 817–829. [Google Scholar] [CrossRef] [PubMed]
- Snee, M.J.; Macdonald, P.M. Dynamic organization and plasticity of sponge bodies. Dev. Dyn. 2009, 238, 918–930. [Google Scholar] [CrossRef] [PubMed]
- McCambridge, A.; Solanki, D.; Olchawa, N.; Govani, N.; Trinidad, J.C.; Gao, M. Comparative Proteomics Reveal Me31B’s Interactome Dynamics, Expression Regulation, and Assembly Mechanism into Germ Granules during Drosophila Germline Development. Sci. Rep. 2020, 10, 564. [Google Scholar] [CrossRef]
- DeHaan, H.; McCambridge, A.; Armstrong, B.; Cruse, C.; Solanki, D.; Trinidad, J.C.; Arkov, A.L.; Gao, M. An in vivo proteomic analysis of the Me31B interactome in Drosophila germ granules. FEBS Lett. 2017, 591, 3536–3547. [Google Scholar] [CrossRef]
- Sankaranarayanan, M.; Emenecker, R.J.; Wilby, E.L.; Jahnel, M.; Trussina, I.R.E.A.; Wayland, M.; Alberti, S.; Holehouse, A.S.; Weil, T.T. Adaptable P body physical states differentially regulate bicoid mRNA storage during early Drosophila development. Dev. Cell 2021, 56, 2886–2901.e6. [Google Scholar] [CrossRef]
- Ferraiuolo, M.A.; Basak, S.; Dostie, J.; Murray, E.L.; Schoenberg, D.R.; Sonenberg, N. A role for the eIF4E-binding protein 4E-T in P-body formation and mRNA decay. J. Cell Biol. 2005, 170, 913–924. [Google Scholar] [CrossRef]
- Davidson, A.; Parton, R.M.; Rabouille, C.; Weil, T.T.; Davis, I. Localized Translation of gurken/TGF-α mRNA during Axis Specification Is Controlled by Access to Orb/CPEB on Processing Bodies. Cell Rep. 2016, 14, 2451–2462. [Google Scholar] [CrossRef]
- Fan, S.-J.; Marchand, V.; Ephrussi, A. Drosophila Ge-1 Promotes P Body Formation and oskar mRNA Localization. PLoS ONE 2011, 6, e20612. [Google Scholar] [CrossRef] [PubMed]
- Weil, T.T.; Parton, R.M.; Herpers, B.; Soetaert, J.; Veenendaal, T.; Xanthakis, D.; Dobbie, I.M.; Halstead, J.M.; Hayashi, R.; Rabouille, C.; et al. Drosophila patterning is established by differential association of mRNAs with P bodies. Nat. Cell Biol. 2012, 14, 1305–1313. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Amikura, R.; Hanyu, K.; Kobayashi, S. Me31B silences translation of oocyte-localizing RNAs through the formation of cytoplasmic RNP complex during Drosophila oogenesis. Development 2001, 128, 3233–3242. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.H.; Barbee, S.A.; Blankenship, J.T. GW-Bodies and P-Bodies Constitute Two Separate Pools of Sequestered Non-Translating RNAs. PLoS ONE 2016, 11, e0150291. [Google Scholar] [CrossRef]
- Pérez-Vilaró, G.; Fernández-Carrillo, C.; Mensa, L.; Miquel, R.; Sanjuan, X.; Forns, X.; Pérez-del-Pulgar, S.; Díez, J. Hepatitis C virus infection inhibits P-body granule formation in human livers. J. Hepatol. 2015, 62, 785–790. [Google Scholar] [CrossRef]
- Derrick, C.J.; Weil, T.T. Translational control of gurken mRNA in Drosophila development. Cell Cycle 2017, 16, 23–32. [Google Scholar] [CrossRef]
- Shimada, Y.; Burn, K.M.; Niwa, R.; Cooley, L. Reversible response of protein localization and microtubule organization to nutrient stress during Drosophila early oogenesis. Dev. Biol. 2011, 355, 250–262. [Google Scholar] [CrossRef]
- Burn, K.M.; Shimada, Y.; Ayers, K.; Vemuganti, S.; Lu, F.; Hudson, A.M.; Cooley, L. Somatic insulin signaling regulates a germline starvation response in Drosophila egg chambers. Dev. Biol. 2015, 398, 206–217. [Google Scholar] [CrossRef]
- Wippich, F.; Vaishali; Hennrich, M.L.; Ephrussi, A. Nutritional stress-induced regulation of microtubule organization and mRNP transport by HDAC1 controlled α-tubulin acetylation. Commun. Biol. 2023, 6, 776. [Google Scholar] [CrossRef]
- Courel, M.; Clément, Y.; Bossevain, C.; Foretek, D.; Vidal Cruchez, O.; Yi, Z.; Bénard, M.; Benassy, M.-N.; Kress, M.; Vindry, C.; et al. GC content shapes mRNA storage and decay in human cells. eLife 2019, 8, e49708. [Google Scholar] [CrossRef]
- Marcelo, A.; Koppenol, R.; de Almeida, L.P.; Matos, C.A.; Nóbrega, C. Stress granules, RNA-binding proteins and polyglutamine diseases: Too much aggregation? Cell Death Dis. 2021, 12, 592. [Google Scholar] [CrossRef] [PubMed]
- Horner, V.L.; Wolfner, M.F. Transitioning from egg to embryo: Triggers and mechanisms of egg activation. Dev. Dyn. 2008, 237, 527–544. [Google Scholar] [CrossRef] [PubMed]
- York-Andersen, A.H.; Parton, R.M.; Bi, C.J.; Bromley, C.L.; Davis, I.; Weil, T.T. A single and rapid calcium wave at egg activation in Drosophila. Biol. Open 2015, 4, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Beadle, L.F.; Love, J.C.; Shapovalova, Y.; Artemev, A.; Rattray, M.; Ashe, H.L. Combined modelling of mRNA decay dynamics and single-molecule imaging in the Drosophila embryo uncovers a role for P-bodies in 5′ to 3′ degradation. PLOS Biol. 2023, 21, e3001956. [Google Scholar]
- Eichler, C.E.; Hakes, A.C.; Hull, B.; Gavis, E.R. Compartmentalized oskar degradation in the germ plasm safeguards germline development. eLife 2020, 9, e49988. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Ly, M.; Lugowski, A.; Laver, J.D.; Lipshitz, H.D.; Smibert, C.A.; Rissland, O.S. ME31B globally represses maternal mRNAs by two distinct mechanisms during the Drosophila maternal-to-zygotic transition. eLife 2017, 6, e27891. [Google Scholar] [CrossRef]
- Barbee, S.A.; Estes, P.S.; Cziko, A.-M.; Hillebrand, J.; Luedeman, R.A.; Coller, J.M.; Johnson, N.; Howlett, I.C.; Geng, C.; Ueda, R.; et al. Staufen- and FMRP-containing neuronal RNPs are structurally and functionally related to somatic P bodies. Neuron 2006, 52, 997–1009. [Google Scholar] [CrossRef]
- Vessey, J.P.; Vaccani, A.; Xie, Y.; Dahm, R.; Karra, D.; Kiebler, M.A.; Macchi, P. Dendritic Localization of the Translational Repressor Pumilio 2 and Its Contribution to Dendritic Stress Granules. J. Neurosci. 2006, 26, 6496–6508. [Google Scholar] [CrossRef]
- Miller, L.C.; Blandford, V.; McAdam, R.; Sanchez-Carbente, M.R.; Badeaux, F.; DesGroseillers, L.; Sossin, W.S. Combinations of DEAD box proteins distinguish distinct types of RNA: Protein complexes in neurons. Mol. Cell Neurosci. 2009, 40, 485–495. [Google Scholar] [CrossRef]
- Cougot, N.; Bhattacharyya, S.N.; Tapia-Arancibia, L.; Bordonné, R.; Filipowicz, W.; Bertrand, E.; Rage, F. Dendrites of Mammalian Neurons Contain Specialized P-Body-Like Structures That Respond to Neuronal Activation. J. Neurosci. 2008, 28, 13793–13804. [Google Scholar] [CrossRef]
- Formicola, N.; Vijayakumar, J.; Besse, F. Neuronal ribonucleoprotein granules: Dynamic sensors of localized signals. Traffic 2019, 20, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Formicola, N.; Heim, M.; Dufourt, J.; Lancelot, A.-S.; Nakamura, A.; Lagha, M.; Besse, F. Tyramine induces dynamic RNP granule remodeling and translation activation in the Drosophila brain. eLife 2021, 10, e65742. [Google Scholar] [CrossRef] [PubMed]
- Zeitelhofer, M.; Karra, D.; Macchi, P.; Tolino, M.; Thomas, S.; Schwarz, M.; Kiebler, M.; Dahm, R. Dynamic Interaction between P-Bodies and Transport Ribonucleoprotein Particles in Dendrites of Mature Hippocampal Neurons. J. Neurosci. 2008, 28, 7555–7562. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Banjade, S.; Cheng, H.-C.; Kim, S.; Chen, B.; Guo, L.; Llaguno, M.; Hollingsworth, J.V.; King, D.S.; Banani, S.F.; et al. Phase transitions in the assembly of multivalent signalling proteins. Nature 2012, 483, 336–340. [Google Scholar] [CrossRef]
- Chen, C.; Jia, H.; Nakamura, Y.; Kanekura, K.; Hayamizu, Y. Effect of Multivalency on Phase-Separated Droplets Consisting of Poly(PR) Dipeptide Repeats and RNA at the Solid/Liquid Interface. ACS Omega 2022, 7, 19280–19287. [Google Scholar] [CrossRef]
- Shapiro, D.M.; Ney, M.; Eghtesadi, S.A.; Chilkoti, A. Protein Phase Separation Arising from Intrinsic Disorder: First-Principles to Bespoke Applications. J. Phys. Chem. B 2021, 125, 6740–6759. [Google Scholar] [CrossRef]
- Martin, E.W.; Holehouse, A.S. Intrinsically disordered protein regions and phase separation: Sequence determinants of assembly or lack thereof. Emerg. Top. Life Sci. 2020, 4, 307–329. [Google Scholar]
- Posey, A.E.; Holehouse, A.S.; Pappu, R.V. Phase Separation of Intrinsically Disordered Proteins. Methods Enzymol. 2018, 611, 1–30. [Google Scholar]
- Molliex, A.; Temirov, J.; Lee, J.; Coughlin, M.; Kanagaraj, A.P.; Kim, H.J.; Mittag, T.; Taylor, J.P. Phase separation by low complexity domains promotes stress granule assembly and drives pathological fibrillization. Cell 2015, 163, 123–133. [Google Scholar] [CrossRef]
- Lee, J.; Cho, H.; Kwon, I. Phase separation of low-complexity domains in cellular function and disease. Exp. Mol. Med. 2022, 54, 1412–1422. [Google Scholar] [CrossRef]
- Kato, M.; Han, T.W.; Xie, S.; Shi, K.; Du, X.; Wu, L.C.; Mirzaei, H.; Goldsmith, E.J.; Longgood, J.; Pei, J.; et al. Cell-free Formation of RNA Granules: Low Complexity Sequence Domains Form Dynamic Fibers within Hydrogels. Cell 2012, 149, 753–767. [Google Scholar] [CrossRef] [PubMed]
- Brangwynne, C.P.; Tompa, P.; Pappu, R.V. Polymer physics of intracellular phase transitions. Nat. Phys. 2015, 11, 899–904. [Google Scholar] [CrossRef]
- Nott, T.J.; Petsalaki, E.; Farber, P.; Jervis, D.; Fussner, E.; Plochowietz, A.; Craggs, T.D.; Bazett-Jones, D.P.; Pawson, T.; Forman-Kay, J.D.; et al. Phase Transition of a Disordered Nuage Protein Generates Environmentally Responsive Membraneless Organelles. Mol. Cell 2015, 57, 936–947. [Google Scholar] [CrossRef]
- Pak, C.W.; Kosno, M.; Holehouse, A.S.; Padrick, S.B.; Mittal, A.; Ali, R.; Yunus, A.A.; Liu, D.R.; Pappu, R.V.; Rosen, M.K. Sequence Determinants of Intracellular Phase Separation by Complex Coacervation of a Disordered Protein. Mol. Cell 2016, 63, 72–85. [Google Scholar] [CrossRef]
- Riback, J.A.; Katanski, C.D.; Kear-Scott, J.L.; Pilipenko, E.V.; Rojek, A.E.; Sosnick, T.R.; Drummond, D.A. Stress-Triggered Phase Separation Is an Adaptive, Evolutionarily Tuned Response. Cell 2017, 168, 1028–1040.e19. [Google Scholar] [CrossRef] [PubMed]
- Murthy, A.C.; Dignon, G.L.; Kan, Y.; Zerze, G.H.; Parekh, S.H.; Mittal, J.; Fawzi, N.L. Molecular interactions underlying liquid-liquid phase separation of the FUS low complexity domain. Nat. Struct. Mol. Biol. 2019, 26, 637–648. [Google Scholar] [CrossRef]
- Dzuricky, M.; Rogers, B.A.; Shahid, A.; Cremer, P.S.; Chilkoti, A. De novo engineering of intracellular condensates using artificial disordered proteins. Nat. Chem. 2020, 12, 814–825. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-H.; Yu, J.H.; Gulick, T.; Bloch, K.D.; Bloch, D.B. RNA-associated protein 55 (RAP55) localizes to mRNA processing bodies and stress granules. RNA 2006, 12, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Ayache, J.; Bénard, M.; Ernoult-Lange, M.; Minshall, N.; Standart, N.; Kress, M.; Weil, D. P-body assembly requires DDX6 repression complexes rather than decay or Ataxin2/2L complexes. Mol. Biol. Cell 2015, 26, 2579–2595. [Google Scholar] [CrossRef]
- Serman, A.; Le Roy, F.; Aigueperse, C.; Kress, M.; Dautry, F.; Weil, D. GW body disassembly triggered by siRNAs independently of their silencing activity. Nucleic Acids Res. 2007, 35, 4715–4727. [Google Scholar] [CrossRef]
- Banani, S.F.; Rice, A.M.; Peeples, W.B.; Lin, Y.; Jain, S.; Parker, R.; Rosen, M.K. Compositional Control of Phase-Separated Cellular Bodies. Cell 2016, 166, 651–663. [Google Scholar] [CrossRef]
- Huch, S.; Nissan, T. An mRNA decapping mutant deficient in P body assembly limits mRNA stabilization in response to osmotic stress. Sci. Rep. 2017, 7, 44395. [Google Scholar] [CrossRef]
- Decker, C.J.; Teixeira, D.; Parker, R. Edc3p and a glutamine/asparagine-rich domain of Lsm4p function in processing body assembly in Saccharomyces cerevisiae. J. Cell Biol. 2007, 179, 437–449. [Google Scholar] [CrossRef]
- Kara, E.; McCambridge, A.; Proffer, M.; Dilts, C.; Pumnea, B.; Eshak, G.; Smith, K.; Fielder, L.; Doyle, D.; Ortega, B.; et al. An In Vivo Analysis of the Functional Motifs of DEAD-box RNA Helicase Me31B in Drosophila Fertility and Germline Development. bioRxiv. 2022. [CrossRef]
- Teixeira, D.; Parker, R. Analysis of P-Body Assembly in Saccharomyces cerevisiae. Mol. Biol. Cell 2007, 18, 2274–2287. [Google Scholar] [CrossRef] [PubMed]
- Rajyaguru, P.; She, M.; Parker, R. Scd6 targets eIF4G to repress translation: RGG-motif proteins as a class of eIF4G-binding proteins. Mol. Cell 2012, 45, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Yang, W.-H.; Gulick, T.; Bloch, K.D.; Bloch, D.B. Ge-1 is a central component of the mammalian cytoplasmic mRNA processing body. RNA 2005, 11, 1795–1802. [Google Scholar] [CrossRef]
- Marnef, A.; Maldonado, M.; Bugaut, A.; Balasubramanian, S.; Kress, M.; Weil, D.; Standart, N. Distinct functions of maternal and somatic Pat1 protein paralogs. RNA 2010, 16, 2094–2107. [Google Scholar] [CrossRef]
- Ozgur, S.; Chekulaeva, M.; Stoecklin, G. Human Pat1b Connects Deadenylation with mRNA Decapping and Controls the Assembly of Processing Bodies. Mol. Cell Biol. 2010, 30, 4308–4323. [Google Scholar] [CrossRef]
- Yang, Z.; Jakymiw, A.; Wood, M.R.; Eystathioy, T.; Rubin, R.L.; Fritzler, M.J.; Chan, E.K.L. GW182 is critical for the stability of GW bodies expressed during the cell cycle and cell proliferation. J. Cell Sci. 2004, 117, 5567–5578. [Google Scholar] [CrossRef]
- Chekulaeva, M.; Hentze, M.W.; Ephrussi, A. Bruno acts as a dual repressor of oskar translation, promoting mRNA oligomerization and formation of silencing particles. Cell 2006, 124, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Bose, M.; Lampe, M.; Mahamid, J.; Ephrussi, A. Liquid-to-solid phase transition of oskar ribonucleoprotein granules is essential for their function in Drosophila embryonic development. Cell 2022, 185, 1308–1324.e23. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Choi, J.-M.; Holehouse, A.S.; Lee, H.O.; Zhang, X.; Jahnel, M.; Maharana, S.; Lemaitre, R.; Pozniakovsky, A.; Drechsel, D.; et al. A molecular grammar governing the driving forces for phase separation of prion-like RNA binding proteins. Cell 2018, 174, 688–699.e16. [Google Scholar] [CrossRef]
- Roden, C.; Gladfelter, A.S. RNA contributions to the form and function of biomolecular condensates. Nat. Rev. Mol. Cell Biol. 2021, 22, 183–195. [Google Scholar] [CrossRef]
- Delanoue, R.; Herpers, B.; Soetaert, J.; Davis, I.; Rabouille, C. Drosophila Squid/hnRNP helps Dynein switch from a gurken mRNA transport motor to an ultrastructural static anchor in sponge bodies. Dev. Cell 2007, 13, 523–538. [Google Scholar] [CrossRef] [PubMed]
- Strome, S.; Updike, D. Specifying and protecting germ cell fate. Nat. Rev. Mol. Cell Biol. 2015, 16, 406–416. [Google Scholar] [CrossRef]
- Hakes, A.C.; Gavis, E.R. Plasticity of Drosophila germ granules during germ cell development. PLOS Biol. 2023, 21, e3002069. [Google Scholar] [CrossRef]
- Thomson, T.; Liu, N.; Arkov, A.; Lehmann, R.; Lasko, P. Isolation of new polar granule components in Drosophila reveals P body and ER associated proteins. Mech. Dev. 2008, 125, 865–873. [Google Scholar] [CrossRef]
- Mahowald, A.P. Fine structure of pole cells and polar granules in Drosophila melanogaster. J. Exp. Zool. 1962, 151, 201–215. [Google Scholar] [CrossRef]
- Trcek, T.; Lehmann, R. Germ granules in Drosophila. Traffic 2019, 20, 650–660. [Google Scholar] [CrossRef]
- Trcek, T.; Douglas, T.E.; Grosch, M.; Yin, Y.; Eagle, W.V.I.; Gavis, E.R.; Shroff, H.; Rothenberg, E.; Lehmann, R. Sequence-Independent Self-Assembly of Germ Granule mRNAs into Homotypic Clusters. Mol. Cell 2020, 78, 941–950.e12. [Google Scholar] [CrossRef] [PubMed]
- Niepielko, M.G.; Eagle, W.V.I.; Gavis, E.R. Stochastic Seeding Coupled with mRNA Self-Recruitment Generates Heterogeneous Drosophila Germ Granules. Curr. Biol. 2018, 28, 1872–1881.e3. [Google Scholar] [CrossRef] [PubMed]
- Curnutte, H.A.; Lan, X.; Sargen, M.; Ieong, S.M.A.; Campbell, D.; Kim, H.; Liao, Y.; Lazar, S.B.; Trcek, T. Proteins rather than mRNAs regulate nucleation and persistence of Oskar germ granules in Drosophila. Cell Rep. 2023, 42, 112723. [Google Scholar] [CrossRef] [PubMed]
- Eagle, W.V.I.; Yeboah-Kordieh, D.K.; Niepielko, M.G.; Gavis, E.R. Distinct cis-acting elements mediate targeting and clustering of Drosophila polar granule mRNAs. Development 2018, 145, dev164657. [Google Scholar] [CrossRef]
- Little, S.C.; Sinsimer, K.S.; Lee, J.J.; Wieschaus, E.F.; Gavis, E.R. Independent and coordinate trafficking of single Drosophila germ plasm mRNAs. Nat. Cell Biol. 2015, 17, 558–568. [Google Scholar] [CrossRef]
- Trcek, T.; Grosch, M.; York, A.; Shroff, H.; Lionnet, T.; Lehmann, R. Drosophila germ granules are structured and contain homotypic mRNA clusters. Nat. Commun. 2015, 6, 7962. [Google Scholar] [CrossRef]
- Valentino, M.; Ortega, B.M.; Ulrich, B.; Doyle, D.A.; Farnum, E.D.; Joiner, D.A.; Gavis, E.R.; Niepielko, M.G. Computational modeling offers new insight into Drosophila germ granule development. Biophys. J. 2022, 121, 1465–1482. [Google Scholar] [CrossRef]
- Bratek-Skicki, A.; Pancsa, R.; Meszaros, B.; Van Lindt, J.; Tompa, P. A guide to regulation of the formation of biomolecular condensates. FEBS J. 2020, 287, 1924–1935. [Google Scholar] [CrossRef]
- Kaneuchi, T.; Sartain, C.V.; Takeo, S.; Horner, V.L.; Buehner, N.A.; Aigaki, T.; Wolfner, M.F. Calcium waves occur as Drosophila oocytes activate. Proc. Natl. Acad. Sci. USA 2015, 112, 791–796. [Google Scholar] [CrossRef]
- Alberti, S. Phase separation in biology. Curr. Biol. 2017, 27, R1097–R1102. [Google Scholar] [CrossRef]
- Zhang, Z.; Ahmed-Braimah, Y.H.; Goldberg, M.L.; Wolfner, M.F. Calcineurin-dependent Protein Phosphorylation Changes During Egg Activation in Drosophila melanogaster. Mol. Cell Proteom. 2019, 18, S145–S158. [Google Scholar] [CrossRef]
- Hara, M.; Lourido, S.; Petrova, B.; Lou, H.J.; Von Stetina, J.R.; Kashevsky, H.; Turk, B.E.; Orr-Weaver, T.L. Identification of PNG kinase substrates uncovers interactions with the translational repressor TRAL in the oocyte-to-embryo transition. eLife 2018, 7, e33150. [Google Scholar] [CrossRef] [PubMed]
- Doane, W.W. Completion of meiosis in uninseminated eggs of Drosophila melanogaster. Science 1960, 132, 677–678. [Google Scholar] [CrossRef] [PubMed]
- Page, A.W.; Orr-Weaver, T.L. Activation of the meiotic divisions in Drosophila oocytes. Dev. Biol. 1997, 183, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Mahowald, A.P.; Goralski, T.J.; Caulton, J.H. In vitro activation of Drosophila eggs. Dev. Biol. 1983, 98, 437–445. [Google Scholar] [CrossRef]
- Heifetz, Y.; Yu, J.; Wolfner, M.F. Ovulation triggers activation of Drosophila oocytes. Dev. Biol. 2001, 234, 416–424. [Google Scholar] [CrossRef]
- York-Andersen, A.H.; Wood, B.W.; Wilby, E.L.; Berry, A.S.; Weil, T.T. Osmolarity-regulated swelling initiates egg activation in Drosophila. Open Biol. 2021, 11, 210067. [Google Scholar] [CrossRef]
- Mohapatra, S.; Wegmann, S. Biomolecular condensation involving the cytoskeleton. Brain Res. Bull. 2023, 194, 105–117. [Google Scholar] [CrossRef]
- Wiegand, T.; Hyman, A.A. Drops and fibers—How biomolecular condensates and cytoskeletal filaments influence each other. Emerg. Top. Life Sci. 2020, 4, 247–261. [Google Scholar]
- York-Andersen, A.H.; Hu, Q.; Wood, B.W.; Wolfner, M.F.; Weil, T.T. A calcium-mediated actin redistribution at egg activation in Drosophila. Mol. Reprod. Dev. 2020, 87, 293–304. [Google Scholar] [CrossRef]
- Avilés-Pagán, E.E.; Hara, M.; Orr-Weaver, T.L. The GNU subunit of PNG kinase, the developmental regulator of mRNA translation, binds BIC-C to localize to RNP granules. eLife 2021, 10, e67294. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Petrova, B.; Orr-Weaver, T.L. Control of PNG kinase, a key regulator of mRNA translation, is coupled to meiosis completion at egg activation. eLife 2017, 6, e22219. [Google Scholar] [CrossRef] [PubMed]
- Brand, A.H.; Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 1993, 118, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Perkins, L.A.; Holderbaum, L.; Tao, R.; Hu, Y.; Sopko, R.; McCall, K.; Yang-Zhou, D.; Flockhart, I.; Binari, R.; Shim, H.-S.; et al. The Transgenic RNAi Project at Harvard Medical School: Resources and Validation. Genetics 2015, 201, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Zirin, J.; Hu, Y.; Liu, L.; Yang-Zhou, D.; Colbeth, R.; Yan, D.; Ewen-Campen, B.; Tao, R.; Vogt, E.; VanNest, S.; et al. Large-Scale Transgenic Drosophila Resource Collections for Loss- and Gain-of-Function Studies. Genetics 2020, 214, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, E.; Chartrand, P.; Schaefer, M.; Shenoy, S.M.; Singer, R.H.; Long, R.M. Localization of ASH1 mRNA particles in living yeast. Mol. Cell 1998, 2, 437–445. [Google Scholar] [CrossRef]
- Forrest, K.M.; Gavis, E.R. Live imaging of endogenous RNA reveals a diffusion and entrapment mechanism for nanos mRNA localization in Drosophila. Curr. Biol. 2003, 13, 1159–1168. [Google Scholar] [CrossRef]
- Tantale, K.; Mueller, F.; Kozulic-Pirher, A.; Lesne, A.; Victor, J.-M.; Robert, M.-C.; Capozi, S.; Chouaib, R.; Bäcker, V.; Mateos-Langerak, J.; et al. A single-molecule view of transcription reveals convoys of RNA polymerases and multi-scale bursting. Nat. Commun. 2016, 7, 12248. [Google Scholar] [CrossRef]
- Dufourt, J.; Bellec, M.; Trullo, A.; Dejean, M.; De Rossi, S.; Favard, C.; Lagha, M. Imaging translation dynamics in live embryos reveals spatial heterogeneities. Science 2021, 372, 840–844. [Google Scholar] [CrossRef]
- Vinter, D.J.; Hoppe, C.; Ashe, H.L. Live and fixed imaging of translation sites at single mRNA resolution in the Drosophila embryo. STAR Protoc. 2021, 2, 100812. [Google Scholar] [CrossRef]
- Raj, A.; van den Bogaard, P.; Rifkin, S.A.; van Oudenaarden, A.; Tyagi, S. Imaging individual mRNA molecules using multiple singly labeled probes. Nat. Methods 2008, 5, 877–879. [Google Scholar] [CrossRef] [PubMed]
- Kramer, S. Simultaneous detection of mRNA transcription and decay intermediates by dual colour single mRNA FISH on subcellular resolution. Nucleic Acids Res. 2017, 45, e49. [Google Scholar] [CrossRef] [PubMed]
- Halstead, J.M.; Wilbertz, J.H.; Wippich, F.; Lionnet, T.; Ephrussi, A.; Chao, J.A. Chapter Six—TRICK: A Single-Molecule Method for Imaging the First Round of Translation in Living Cells and Animals. In Methods in Enzymology; Filonov, G.S., Jaffrey, S.R., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 572, pp. 123–157. [Google Scholar]
- Halstead, J.M.; Lionnet, T.; Wilbertz, J.H.; Wippich, F.; Ephrussi, A.; Singer, R.H.; Chao, J.A. Translation. An RNA biosensor for imaging the first round of translation from single cells to living animals. Science 2015, 347, 1367–1671. [Google Scholar] [CrossRef] [PubMed]
- Tanenbaum, M.E.; Gilbert, L.A.; Qi, L.S.; Weissman, J.S.; Vale, R.D. A protein-tagging system for signal amplification in gene expression and fluorescence imaging. Cell 2014, 159, 635–646. [Google Scholar] [CrossRef]
- Yan, X.; Hoek, T.A.; Vale, R.D.; Tanenbaum, M.E. Dynamics of Translation of Single mRNA Molecules In Vivo. Cell 2016, 165, 976–989. [Google Scholar] [CrossRef] [PubMed]
- Pichon, X.; Bastide, A.; Safieddine, A.; Chouaib, R.; Samacoits, A.; Basyuk, E.; Peter, M.; Mueller, F.; Bertrand, E. Visualization of single endogenous polysomes reveals the dynamics of translation in live human cells. J. Cell Biol. 2016, 214, 769–781. [Google Scholar] [CrossRef]
- Wang, C.; Han, B.; Zhou, R.; Zhuang, X. Real-time imaging of translation on single mRNA transcripts in live cells. Cell 2016, 165, 990–1001. [Google Scholar] [CrossRef]
- Wu, B.; Eliscovich, C.; Yoon, Y.J.; Singer, R.H. Translation dynamics of single mRNAs in live cells and neurons. Science 2016, 352, 1430–1435. [Google Scholar] [CrossRef]
- Vinter, D.J.; Hoppe, C.; Minchington, T.G.; Sutcliffe, C.; Ashe, H.L. Dynamics of hunchback translation in real-time and at single-mRNA resolution in the Drosophila embryo. Development 2021, 148, dev196121. [Google Scholar] [CrossRef]
- Bellec, M.; Chen, R.; Dhayni, J.; Favard, C.; Trullo, A.; Lenden-Hasse, H.; Lehmann, R.; Bertrand, E.; Lagha, M.; Dufourt, J. Boosting the toolbox for live imaging of translation. BioRxiv 2023, preprint. [Google Scholar]
- Garcia, J.F.; Parker, R. MS2 coat proteins bound to yeast mRNAs block 5′ to 3′ degradation and trap mRNA decay products: Implications for the localization of mRNAs by MS2-MCP system. RNA 2015, 21, 1393–1395. [Google Scholar] [CrossRef] [PubMed]
- Pandey, U.B.; Nichols, C.D. Human disease models in Drosophila melanogaster and the role of the fly in therapeutic drug discovery. Pharmacol. Rev. 2011, 63, 411–436. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Ligoxygakis, P. Beyond Host Defense: Deregulation of Drosophila Immunity and Age-Dependent Neurodegeneration. Front. Immunol. 2020, 11, 1574. [Google Scholar] [CrossRef]
- Verheyen, E.M. The power of Drosophila in modeling human disease mechanisms. Dis. Model. Mech. 2022, 15, dmm049549. [Google Scholar] [CrossRef]
- Cheng, L.; Baonza, A.; Grifoni, D. Drosophila Models of Human Disease. BioMed Res. Int. 2018, 2018, 7214974. [Google Scholar] [CrossRef]
- Tapia, A.; Giachello, C.N.; Palomino-Schätzlein, M.; Baines, R.A.; Galindo, M.I. Generation and Characterization of the Drosophila melanogaster paralytic Gene Knock-Out as a Model for Dravet Syndrome. Life 2021, 11, 1261. [Google Scholar] [CrossRef]
- Kim, K.; Lane, E.A.; Saftien, A.; Wang, H.; Xu, Y.; Wirtz-Peitz, F.; Perrimon, N. Drosophila as a model for studying cystic fibrosis pathophysiology of the gastrointestinal system. Proc. Natl. Acad. Sci. USA 2020, 117, 10357–10367. [Google Scholar] [CrossRef] [PubMed]
- Huggett, S.B.; Hatfield, J.S.; Walters, J.D.; McGeary, J.E.; Welsh, J.W.; Mackay, T.F.C.; Anholt, R.R.H.; Palmer, R.H.C. Ibrutinib as a potential therapeutic for cocaine use disorder. Transl. Psychiatry 2021, 11, 623. [Google Scholar] [CrossRef]
- Rouka, E.; Gourgoulianni, N.; Lüpold, S.; Hatzoglou, C.; Gourgoulianis, K.I.; Zarogiannis, S.G. Prediction and enrichment analyses of the Homo sapiens-Drosophila melanogaster COPD-related orthologs: Potential for modeling of human COPD genomic responses with the fruit fly. Am. J. Physiol. Integr. Comp. Physiol. 2022, 322, R77–R82. [Google Scholar] [CrossRef]
- Kotian, N.; Troike, K.M.; Curran, K.N.; Lathia, J.D.; McDonald, J.A. A Drosophila RNAi screen reveals conserved glioblastoma-related adhesion genes that regulate collective cell migration. G3 GenesGenomesGenetics 2021, 12, jkab356. [Google Scholar] [CrossRef]
- Belfer, S.J.; Bashaw, A.G.; Perlis, M.L.; Kayser, M.S. A Drosophila model of sleep restriction therapy for insomnia. Mol. Psychiatry 2021, 26, 492–507. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.-X.; Li, S.-S.; Li, A.-Q.; Liu, Z.-Y.; Neely, G.G.; Wang, Q.-P. dSec16 Acting in Insulin-like Peptide Producing Cells Controls Energy Homeostasis in Drosophila. Life 2022, 13, 81. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Matsukawa, K.; Watanabe, N.; Kishino, Y.; Kunugi, H.; Ihara, R.; Wakabayashi, T.; Hashimoto, T.; Iwatsubo, T. Self-assembly of FUS through its low-complexity domain contributes to neurodegeneration. Hum. Mol. Genet. 2018, 27, 1353–1365. [Google Scholar] [CrossRef]
- An, H.; Litscher, G.; Watanabe, N.; Wei, W.; Hashimoto, T.; Iwatsubo, T.; Buchman, V.L.; Shelkovnikova, T.A. ALS-linked cytoplasmic FUS assemblies are compositionally different from physiological stress granules and sequester hnRNPA3, a novel modifier of FUS toxicity. Neurobiol. Dis. 2022, 162, 105585. [Google Scholar] [CrossRef]
- Matsukawa, K.; Kukharsky, M.S.; Park, S.K.; Park, S.; Watanabe, N.; Iwatsubo, T.; Shelkovnikova, T.A. Long non-coding RNA NEAT1_1 ameliorates TDP-43 toxicity in in vivo models of TDP-43 proteinopathy. RNA Biol. 2021, 18, 1546–1554. [Google Scholar] [CrossRef] [PubMed]
- Markmiller, S.; Soltanieh, S.; Server, K.L.; Mak, R.; Jin, W.; Fang, M.Y.; Luo, E.-C.; Krach, F.; Yang, D.; Sen, A.; et al. Context-Dependent and Disease-Specific Diversity in Protein Interactions within Stress Granules. Cell 2018, 172, 590–604.e13. [Google Scholar] [CrossRef]
- Sharpe, J.L.; Harper, N.S.; Garner, D.R.; West, R.J.H. Modeling C9orf72-Related Frontotemporal Dementia and Amyotrophic Lateral Sclerosis in Drosophila. Front. Cell Neurosci. 2021, 15, 770937. [Google Scholar] [CrossRef]
- Sreedharan, J.; Blair, I.P.; Tripathi, V.B.; Hu, X.; Vance, C.; Rogelj, B.; Ackerley, S.; Durnall, J.C.; Williams, K.L.; Buratti, E.; et al. TDP-43 mutations in familial and sporadic amyotrophic lateral sclerosis. Science 2008, 319, 1668–1672. [Google Scholar] [CrossRef]
- Vance, C.; Rogelj, B.; Hortobágyi, T.; De Vos, K.J.; Nishimura, A.L.; Sreedharan, J.; Hu, X.; Smith, B.; Ruddy, D.; Wright, P.; et al. Mutations in FUS, an RNA Processing Protein, Cause Familial Amyotrophic Lateral Sclerosis Type 6. Science 2009, 323, 1208–1211. [Google Scholar] [CrossRef]
- Patel, A.; Lee, H.O.; Jawerth, L.; Maharana, S.; Jahnel, M.; Hein, M.Y.; Stoynov, S.; Mahamid, J.; Saha, S.; Franzmann, T.M.; et al. A Liquid-to-Solid Phase Transition of the ALS Protein FUS Accelerated by Disease Mutation. Cell 2015, 162, 1066–1077. [Google Scholar] [CrossRef]
- Mackenzie, I.R.; Nicholson, A.M.; Sarkar, M.; Messing, J.; Purice, M.D.; Pottier, C.; Rademakers, R. TIA1 Mutations in Amyotrophic Lateral Sclerosis and Frontotemporal Dementia Promote Phase Separation and Alter Stress Granule Dynamics. Neuron 2017, 95, 808–816.e9. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Pu, J. α-Synuclein in Parkinson’s Disease: From Pathogenetic Dysfunction to Potential Clinical Application. Park. Dis. 2016, 2016, 1720621. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.L.; Feany, M.B. Glial α-synuclein promotes neurodegeneration characterized by a distinct transcriptional program in vivo. Glia 2019, 67, 1933–1957. [Google Scholar] [CrossRef] [PubMed]
- Feany, M.B.; Bender, W.W. A Drosophila model of Parkinson’s disease. Nature 2000, 404, 394–398. [Google Scholar] [CrossRef]
- Xiong, Y.; Yu, J. Modeling Parkinson’s Disease in Drosophila: What Have We Learned for Dominant Traits? Front. Neurol. 2018, 9, 228. [Google Scholar] [CrossRef]
- Balak, C.; Benard, M.; Schaefer, E.; Iqbal, S.; Ramsey, K.; Ernoult-Lange, M.; Mattioli, F.; Llaci, L.; Geoffroy, V.; Courel, M.; et al. Rare De Novo Missense Variants in RNA Helicase DDX6 Cause Intellectual Disability and Dysmorphic Features and Lead to P-Body Defects and RNA Dysregulation. Am. J. Hum. Genet. 2019, 105, 509–525. [Google Scholar] [CrossRef]
- Chan, H.Y.E.; Bonini, N.M. Drosophila models of human neurodegenerative disease. Cell Death Differ. 2000, 7, 1075–1080. [Google Scholar] [CrossRef]
- Hardy, S.D.; Shinde, A.; Wang, W.-H.; Wendt, M.K.; Geahlen, R.L. Regulation of epithelial-mesenchymal transition and metastasis by TGF-β, P-bodies, and autophagy. Oncotarget 2017, 8, 103302–103314. [Google Scholar] [CrossRef]
- Tang, L.; Zhao, P.; Kong, D. Muscleblind-like 1 destabilizes Snail mRNA and suppresses the metastasis of colorectal cancer cells via the Snail/E-cadherin axis. Int. J. Oncol. 2019, 54, 955–965. [Google Scholar] [CrossRef]
- Corcoran, J.A.; Khaperskyy, D.A.; Johnston, B.P.; King, C.A.; Cyr, D.P.; Olsthoorn, A.V.; McCormick, C. Kaposi’s sarcoma-associated herpesvirus G-protein-coupled receptor prevents AU-rich-element-mediated mRNA decay. J. Virol. 2012, 86, 8859–8871. [Google Scholar] [CrossRef]
- Hopkins, K.C.; McLane, L.M.; Maqbool, T.; Panda, D.; Gordesky-Gold, B.; Cherry, S. A genome-wide RNAi screen reveals that mRNA decapping restricts bunyaviral replication by limiting the pools of Dcp2-accessible targets for cap-snatching. Genes Dev. 2013, 27, 1511–1525. [Google Scholar] [CrossRef]
- Khong, A.; Jan, E. Modulation of Stress Granules and P Bodies during Dicistrovirus Infection. J. Virol. 2011, 85, 1439–1451. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Magwire, M.M.; Bayer, F.; Jiggins, F.M. A Polymorphism in the Processing Body Component Ge-1 Controls Resistance to a Naturally Occurring Rhabdovirus in Drosophila. PLOS Pathog. 2016, 12, e1005387. [Google Scholar]
- Göertz, G.P.; van Bree, J.W.M.; Hiralal, A.; Fernhout, B.M.; Steffens, C.; Boeren, S.; Visser, T.M.; Vogels, C.B.F.; Abbo, S.R.; Fros, J.J.; et al. Subgenomic flavivirus RNA binds the mosquito DEAD/H-box helicase ME31B and determines Zika virus transmission by Aedes aegypti. Proc. Natl. Acad. Sci. USA 2019, 116, 19136–19144. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, S.; Xu, F.; Mei, S.; Liu, X.; Yin, L.; Zhao, F.; Zhao, X.; Sun, H.; Xiong, Z.; et al. MOV10 sequesters the RNP of influenza A virus in the cytoplasm and is antagonized by viral NS1 protein. Biochem. J. 2019, 476, 467–481. [Google Scholar] [CrossRef]
- Liu, Y.-C.; Mok, B.W.-Y.; Wang, P.; Kuo, R.-L.; Chen, H.; Shih, S.-R. Cellular 5′-3′ mRNA Exoribonuclease XRN1 Inhibits Interferon β Activation and Facilitates Influenza A Virus Replication. mBio 2021, 12, e0094521. [Google Scholar] [CrossRef]
- Welke, R.-W.; Sperber, H.S.; Bergmann, R.; Koikkarah, A.; Menke, L.; Sieben, C.; Krüger, D.H.; Chiantia, S.; Herrmann, A.; Schwarzer, R. Characterization of Hantavirus N Protein Intracellular Dynamics and Localization. Viruses 2022, 14, 457. [Google Scholar] [CrossRef]
- Oceguera, A.; Peralta, A.V.; Martínez-Delgado, G.; Arias, C.F.; López, S. Rotavirus RNAs sponge host cell RNA binding proteins and interfere with their subcellular localization. Virology 2018, 525, 96–105. [Google Scholar] [CrossRef]
- Kuroshima, T.; Matsuda, A.Y.; Hossain, E.; Yasuda, M.; Kitamura, T.; Kitagawa, Y.; Higashino, F. Adenovirus infection controls processing bodies to stabilize AU-rich element-containing mRNA. Virology 2022, 573, 124–130. [Google Scholar] [CrossRef]
- Fan, S.; Xu, Z.; Liu, P.; Qin, Y.; Chen, M. Enterovirus 71 2A Protease Inhibits P-Body Formation to Promote Viral RNA Synthesis. J. Virol. 2021, 95, e0092221. [Google Scholar] [CrossRef]
- Perez-Pepe, M.; Fernández-Alvarez, A.J.; Boccaccio, G.L. Life and Work of Stress Granules and Processing Bodies: New Insights into Their Formation and Function. Biochemistry 2018, 57, 2488–2498. [Google Scholar] [CrossRef]
- Fernández-Carrillo, C.; Pérez-Vilaró, G.; Díez, J.; Pérez-del-Pulgar, S. Hepatitis C virus plays with fire and yet avoids getting burned. A review for clinicians on processing bodies and stress granules. Liver Int. 2018, 38, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Ariumi, Y. Host Cellular RNA Helicases Regulate SARS-CoV-2 Infection. J. Virol. 2022, 96, e00002-22. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, N.M. Challenges and opportunities in controlling mosquito-borne infections. Nature 2018, 559, 490–497. [Google Scholar] [CrossRef] [PubMed]
- Kwaśnik, M.; Rożek, W.; Rola, J. Rift Valley Fever—A Growing Threat to Humans and Animals. J. Vet. Res. 2021, 65, 7–14. [Google Scholar] [CrossRef]
- Liu, L.; Weiss, E.; Panas, M.D.; Götte, B.; Sellberg, S.; Thaa, B.; McInerney, G.M. RNA processing bodies are disassembled during Old World alphavirus infection. J. Gen. Virol. 2019, 100, 1375–1389. [Google Scholar] [CrossRef]
- Morrison, T.E. Reemergence of Chikungunya Virus. J. Virol. 2014, 88, 11644–11647. [Google Scholar] [CrossRef]
- Hopkins, K.; Cherry, S. Bunyaviral cap-snatching vs. decapping. Cell Cycle 2013, 12, 3711–3712. [Google Scholar] [CrossRef]
- Harnish, J.M.; Link, N.; Yamamoto, S. Drosophila as a Model for Infectious Diseases. Int. J. Mol. Sci. 2021, 22, 2724. [Google Scholar] [CrossRef]
- Hedges, S.B.; Dudley, J.; Kumar, S. TimeTree: A public knowledge-base of divergence times among organisms. Bioinformatics 2006, 22, 2971–2972. [Google Scholar] [CrossRef]
- Rubin, G.M.; Yandell, M.D.; Wortman, J.R.; Gabor, G.L.; Miklos, G.L.G.; Nelson, C.R.; Hariharan, I.K.; Fortini, M.E.; Li, P.W.; Apweiler, R.; et al. Comparative genomics of the eukaryotes. Science 2000, 287, 2204–2215. [Google Scholar] [CrossRef] [PubMed]
- Reiter, L.T.; Potocki, L.; Chien, S.; Gribskov, M.; Bier, E. A Systematic Analysis of Human Disease-Associated Gene Sequences in Drosophila melanogaster. Genome Res. 2001, 11, 1114–1125. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).