
Genome-Wide Identification of the bHLH Gene Family in Rhododendron delavayi and Its Expression Analysis in Different Floral Tissues

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Identification of the Complete Genome of bHLH Genes in R. delavayi, Prediction of Physicochemical Properties of Encoded Proteins, and Analysis of Gene Locus Prediction
2.2.2. Phylogenetic and Evolutionary Analysis of the R. delavayi bHLH Gene Family
2.2.3. Structural Analysis of Genes in the R. delavayi bHLH Gene Family
2.2.4. Conserved Motif Analysis of R. delavayi bHLH Gene Family Proteins
2.2.5. Promoter Analysis of the R. delavayi bHLH Genes
2.2.6. Expression Pattern Analysis of the R. delavayi bHLH Gene Family
3. Result and Analysis
3.1. Identification and Physicochemical Properties of R. delavayi bHLH Gene Family Proteins
3.2. Phylogenetic Analysis of the R. delavayi bHLH Gene Family
3.3. Gene Structure Analysis of the R. delavayi bHLH Gene Family
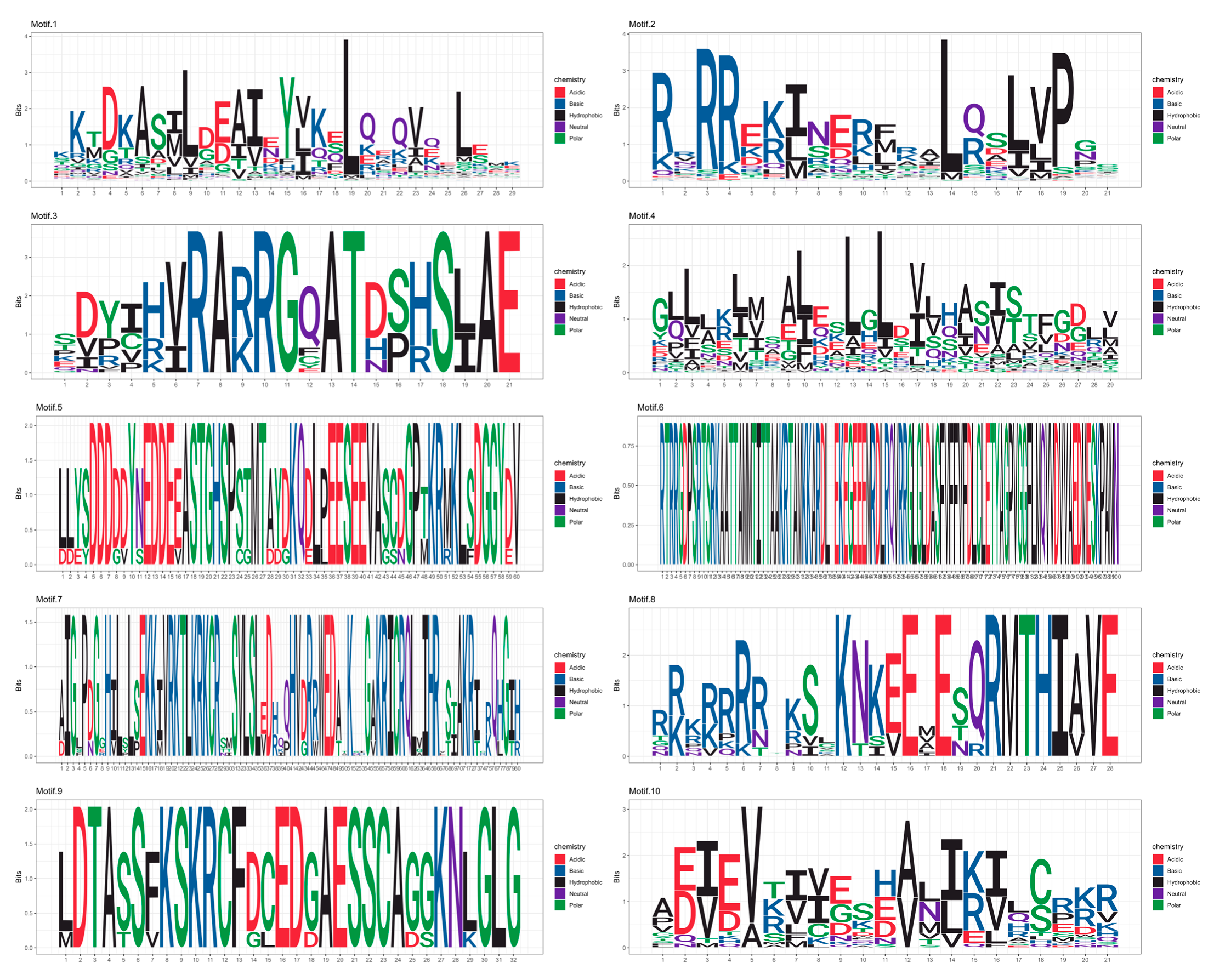
3.4. Conserved Motif Analysis of the R. delavayi bHLH Gene Family
3.5. Cis-Regulatory Element Analysis of R. delavayi bHLH Gene Family Promoters
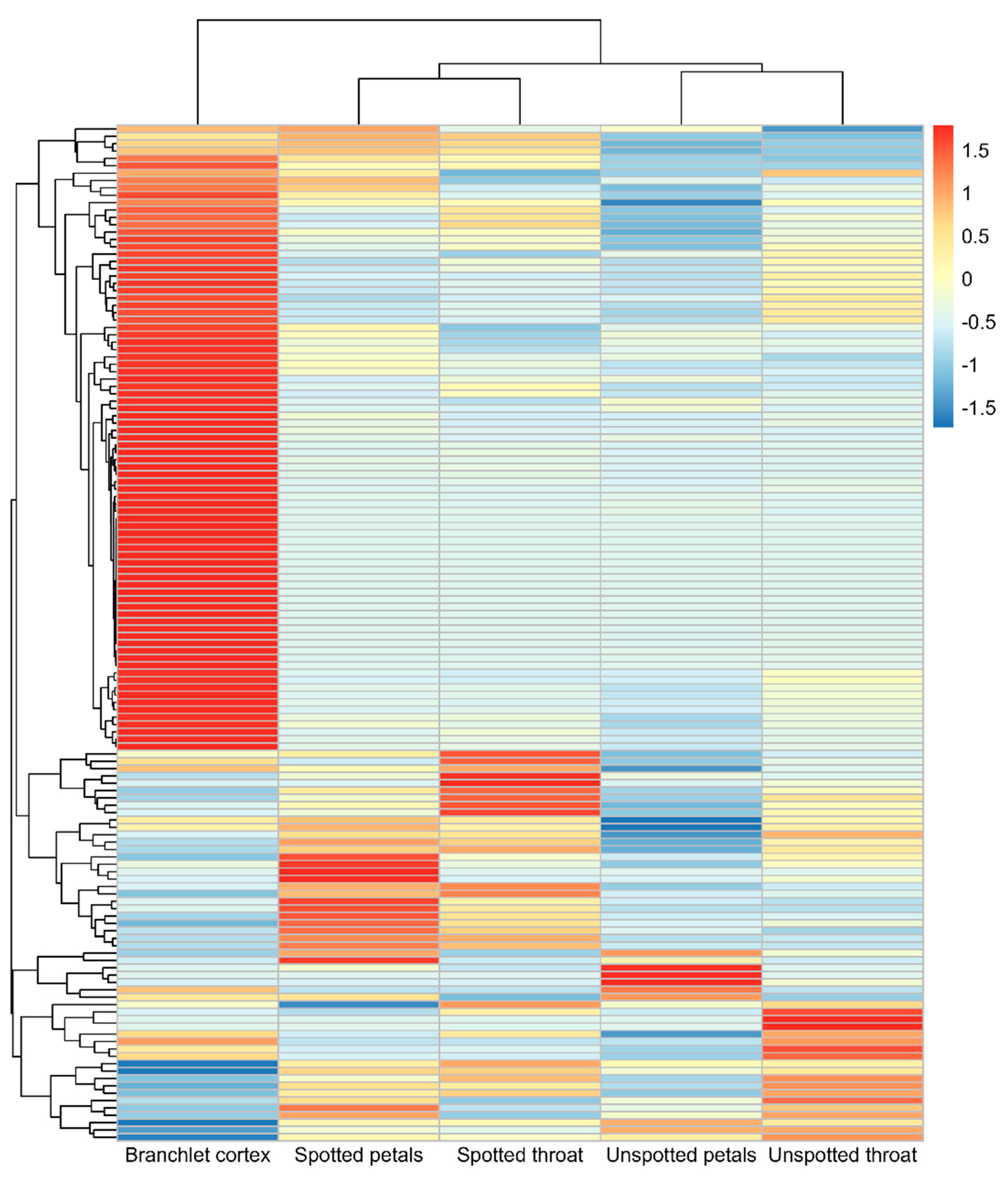
3.6. Expression Pattern Analysis of R. delavayi bHLH Gene Family Members
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xia, J.; Li, Q.L.; Cheng, S.L. Global Rhododendron Experts Gathered in Macheng. 2010. Available online: https://www.gmw.cn/01gmrb/2010-04/30/content_1107339.htm (accessed on 23 August 2024).
- Committee on the Flora of China. Flora of China; Chinese Academy of Sciences: Beijing, China, 1991; Volume 57. [Google Scholar]
- Zhou, W.W. Yunnan Agricultural University National Rhododendron Germplasm Resource Bank Let Yunnan Rhododendron Resources Really Go Out of the Mountain. Chin. Flower. Hortic. 2020, 23, 36–37. [Google Scholar]
- Ou, J.; Chen, X. Analysis of the resources of the subgenus Rhododendron evergreen in Guizhou Province and the prospect of garden application. Jiangsu Agric. Sci. 2012, 40, 200–203. [Google Scholar]
- Fu, T. Spatial Distribution Characteristics of Rhododendrons of Different Age Classes and Their Main Influencing Factors. Master’s Thesis, Guizhou University, Guizhou, China, 2022. [Google Scholar]
- Wang, H.F.; Ou, J.; Wang, X.J.; Ke, K.; Fan, Y.; Zhou, Y.M.; Fan, X.Y. Identification and expression analysis of a family of bHLH transcription factors in Rhododendron rhododendron. Guangxi Plants 2023, 1–21. [Google Scholar]
- Sun, W.; Zhou, N.; Wang, Y.; Sun, S.; Zhang, Y.; Ju, Z.; Yi, Y. Characterization and functional analysis of RdDFR1 regulation on flower color formation in Rhododendron delavayi. Plant Physiol. Biochem. PPB 2021, 169, 203–210. [Google Scholar] [CrossRef]
- Cai, Y.F.; Wang, J.H.; Zhang, L.; Song, J.; Peng, L.C.; Zhang, S.B. Physiological and transcriptomic analysis highlight key metabolic pathways in relation to drought tolerance in Rhododendron delavayi. Physiol. Mol. Biol. Plants 2019, 25, 991–1008. [Google Scholar] [CrossRef]
- Wang, H.; Mu, K.; Liu, C.; Guo, Y.; Deng, X. Gene expression profiling of Rhododendron pulchrum leaves under drought stress. Tree Genet. Genomes 2020, 16, 58. [Google Scholar] [CrossRef]
- Guo, A.Y.; Chen, X.; Gao, G.; Zhang, H.; Zhu, Q.H. PlantTFDB: A comprehensive plant transcription factor database. Nucl. Acid. Res. 2008, 36 (Suppl. S1), D966–D969. [Google Scholar] [CrossRef]
- Wang, J.; Yang, S.; Sun, Q.W.; Chen, H.Y.; Yang, T.; Huang, Y. Genome-wide identification and expression analysis of the bHLH transcription factor family in Dendrobium cinereum. Biotechnol. Bull. 2024, 40, 203–218. [Google Scholar]
- Atchley, W.R.; Terhalle, W.; Dress, A. Positional Dependence, Cliques, and Predictive Motifs in the bHLH Protein Domain. J. Mol. Evol. 1999, 48, 501–516. [Google Scholar] [CrossRef]
- Liu, W.; Tai, H.; Li, S.; Gao, W.; Zhao, M.; Xie, C.; Li, W.X. b HLH 122 is important for drought and osmotic stress resistance in Arabidopsisand in the repression of ABA catabolism. N. Phytol. 2014, 201, 1192–1204. [Google Scholar] [CrossRef]
- He, K.P.; Wu, C. Effects of bHLH transcription factors on plant morphogenesis. Anhui Agric. Sci. 2010, 38, 19957–19959. [Google Scholar]
- Gu, C.S.; Zhao, C.L.; Zang, L.F.; Wen, G.S.; Xu, S.Z.; Zhang, H.L. Longitudinal Changes of Transcriptional Levels of bHLH, MYB and WD40 and Their Relationships with Total Anthocyanin Contents in the Green-purple Aerial Stems of Panax notoginseng. J. Yunnan Agric. Univ. (Nat. Sci.) 2020, 35, 581–590. [Google Scholar]
- David, S.; Christopher, W.; Philippe, L.; Berardini, T.Z.; Margarita, G.H.; Hartmut, F.; Donghui, L.; Tom, M.; Robert, M.; Larry, P. The Arabidopsis Information Resource (TAIR): Gene structure and function annotation. Nucleic Acids Res. 2007, 36 (Suppl. S1), D1009–D1014. [Google Scholar]
- Long, F.; Wu, H.; Li, H.; Zuo, W.; Ao, Q. Genome-Wide Analysis of MYB Transcription Factors and Screening of MYBs Involved in the Red Color Formation in Rhododendron delavayi. Int. J. Mol. Sci. 2023, 24, 4641. [Google Scholar] [CrossRef]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, 29–37. [Google Scholar] [CrossRef]
- Katoh, K.; Kuma, K.I.; Toh, H.; Miyata, T. MAFFT version 5: Improvement in accuracy of multiple sequence alignment. Nucleic Acids Res. 2005, 33, 511–518. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2–Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.Y. ggtree: An r package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2014, 31, 1296. [Google Scholar] [CrossRef]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef]
- Li, X.; Ma, L.; Mei, X.; Liu, Y.; Huang, H. ggmotif: An R Package for the extraction and visualization of motifs from MEME software. PLoS ONE 2022, 17, e0276979. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H. ggplot2. Wiley Interdiscip. Rev. Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Danecek, P.; Bonfield, J.K.; Liddle, J.; Marshall, J.; Ohan, V.; Pollard, M.O.; Whitwham, A.; Keane, T.; McCarthy, S.A.; Davies, R.M.; et al. Twelve years of SAMtools and BCFtools. Gigascience 2021, 10, giab008. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- He, Z.M.; Zhang, J.X.; Yu, L.P.; Hu, K.; JI, F.Q.; Huang, T.R.; Yang, M.F.; Ma, L.Q. Identification and analysis of bHLH transcription factor family in red raspberry based on transcriptome sequencing. Guangxi Plant 2024, 1–11. [Google Scholar]
- Wei, K.; Chen, H. Comparative functional genomics analysis of bHLH gene family in rice, maize and wheat. BMC Plant Biol. 2018, 18, 1–21. [Google Scholar] [CrossRef]
- Zhao, S.; Li, X.; Su, X.Y.; Li, X.E.; Zhao, Y.; Ren, Y.L.; Sun, S.N.; Li, P. Identification and bioinformatics analysis of a family of bHLH transcription factors in Corydalis nipponensis. Mol. Plant Breed. 2024, 1–29. [Google Scholar]
- Nawaz, Z.; Kakar, K.; Saand, M.A.; Shu, Q.Y. Cyclic nucleotide-gated ion channel gene family in rice, identification, characterization and experimental analysis of expression response to plant hormones, biotic and abiotic stresses. Bmc Genom. 2014, 15, 853. [Google Scholar] [CrossRef]
- Zhao, L. Effect of bHLH Homologous Gene Expression on Flower Color of Chrysanthemums. Master’s Thesis, Beijing Forestry University, Beijing, China, 2011. [Google Scholar]
- Li, Y.Q.; Wu, N.; Luo, J.J. Cloning and functional analysis of bHLH, a gene related to floral chromosome synthesis in Ocimum sanctum. Biotechnol. Bull. 2024, 40, 1–12. [Google Scholar]
- Zhang, Y.Y. Study on the Distribution Pattern of Flower Color Number and Regulatory Genes of Sichuan Solanum Orchid Population. Master’s Thesis, 2019. [Google Scholar]
- Zeng, S.; Wu, M.; Zou, C.; Liu, X.; Wang, Y. Comparative analysis of anthocyanin biosynthesis during fruit development in two Lycium species. Physiol. Plant 2014, 150, 505–516. [Google Scholar] [CrossRef]
- Ge, S.B.; Zhang, X.N.; Han, W.Y. Research progress on plant flavonoids biosynthesis and their anti-stress mechanism. Acta Hortic. Sin. 2023, 50, 209–224. [Google Scholar]
- He, W.Z.; Lei, W.Q.; Guo, X.X. Identification of the MYB gene family and functional analysis of key genes related to blue flower coloration in Agapanthus praecox. Acta Hortic. Sin. 2023, 50, 1255–1268. [Google Scholar]
- Zou, H.Z.; Han, L.L.; Zhou, L. Identification of MYB family members and functional verification of PdMYB2 in Paeonia delavayi. For. Res. 2022, 35, 1–13. [Google Scholar]
- Wan, Y.B. Identification and functional analysis of transcription factors related to flower color formation in Pachysandra chinensis. Chin. Acad. Agric. Sci. 2023. [Google Scholar]
- Zhou, L.; Huang, S.M.; Su, W.K. Identification of bHLH gene family in Camellia sinensis and screening of genes related to flower color formation. Biotechnol. Bull. 2024, 40, 142–151. [Google Scholar]
- Chen, J.H.; Jiang, L.L.; Qian, J.Y. Screening and expression analysis of IIIf subgroup gene of bHLH family in Zinnia elegans. J. Agric. Biotechnol. 2023, 31, 61–72. [Google Scholar]
- Ya, H.J.; Tian, J.H.; Shen, L.W. Identification and analysis of a family of bHLH transcription factors in Rhea [Mucuna pruriens] from Yunnan. Southwest J. Agric. 1–10.








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, J.; Wu, Y.-W.; Dong, Y.; Pu, R.; Li, X.-J.; Lyu, Y.-M.; Bai, T.; Zhang, J.-L. Genome-Wide Identification of the bHLH Gene Family in Rhododendron delavayi and Its Expression Analysis in Different Floral Tissues. Genes 2024, 15, 1256. https://doi.org/10.3390/genes15101256
Dong J, Wu Y-W, Dong Y, Pu R, Li X-J, Lyu Y-M, Bai T, Zhang J-L. Genome-Wide Identification of the bHLH Gene Family in Rhododendron delavayi and Its Expression Analysis in Different Floral Tissues. Genes. 2024; 15(10):1256. https://doi.org/10.3390/genes15101256
Chicago/Turabian StyleDong, Jian, Ya-Wen Wu, Yan Dong, Ran Pu, Xue-Jiao Li, Ying-Min Lyu, Tian Bai, and Jing-Li Zhang. 2024. "Genome-Wide Identification of the bHLH Gene Family in Rhododendron delavayi and Its Expression Analysis in Different Floral Tissues" Genes 15, no. 10: 1256. https://doi.org/10.3390/genes15101256