
Cognitive Function Is Associated with the Genetically Determined Efficiency of DNA Repair Mechanisms

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. DNA Repair Single Nucleotide Polymorphism
2.3. Cognitive Measures
2.4. Brain Measures
2.4.1. Image Acquisition
2.4.2. Segmentation and Image Analysis
2.5. Socio-Demographic and Health Measures
2.6. Statistical Analysis
3. Results
3.1. SNPs—Cognition Associations
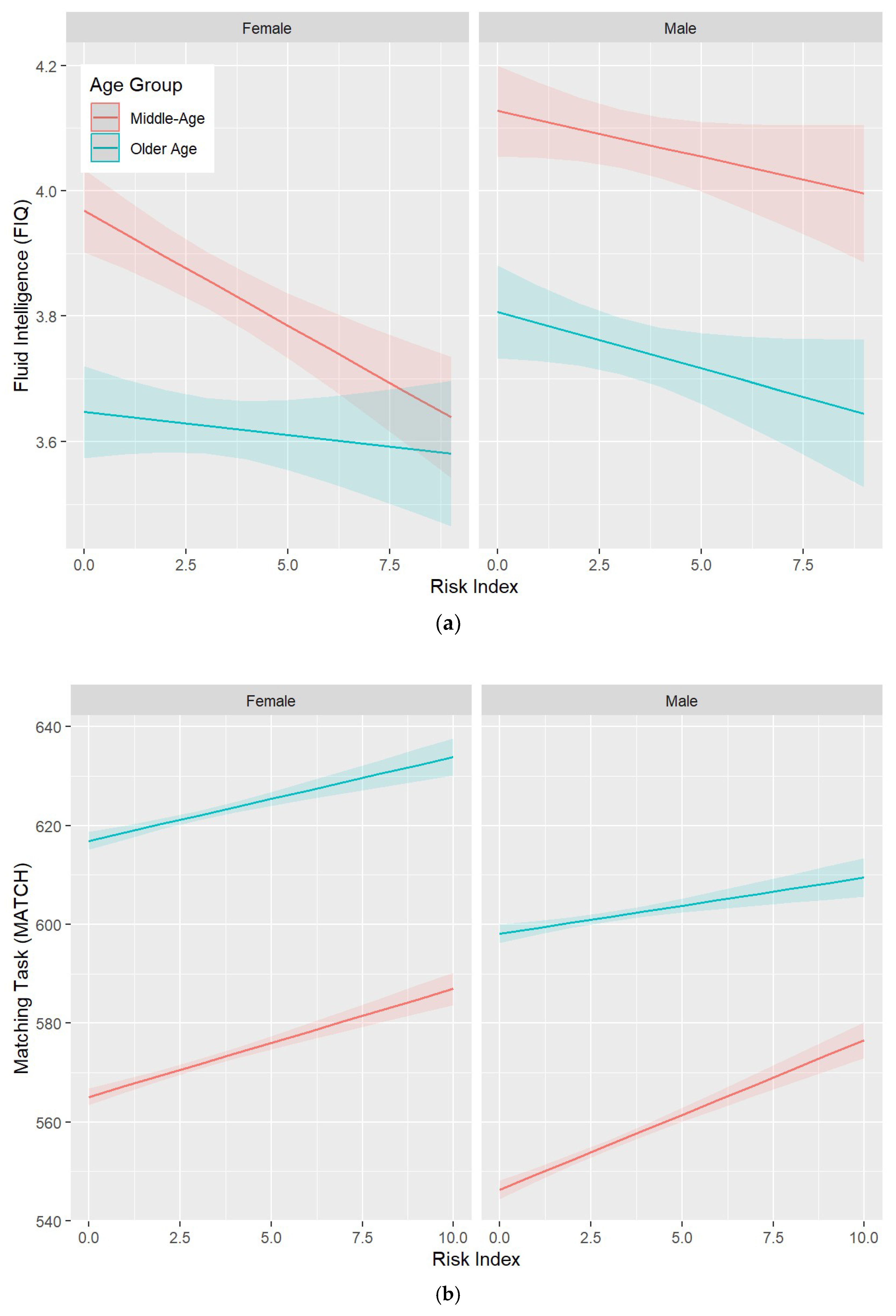
3.2. Protect, Harm, and Risk Indexes
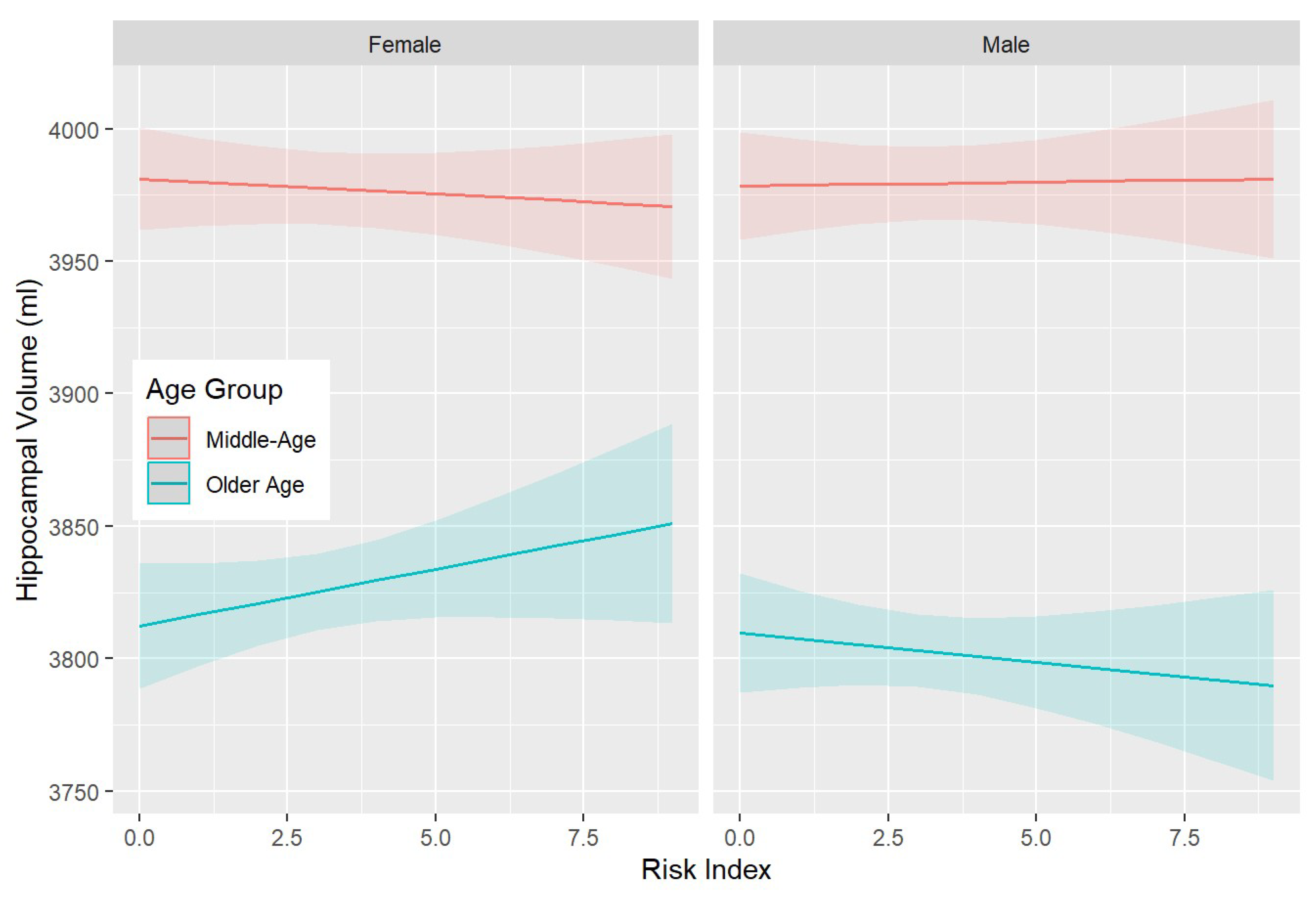
3.3. SNPs—Brain Associations
3.4. Latent Class Analyses
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tabatabaei-Jafari, H.; Shaw, M.E.; Walsh, E.; Cherbuin, N.; for the Alzheimer’s Disease Neuroimaging Initiative (ADNI). Cognitive/Functional Measures Predict Alzheimer’s Disease, Dependent on Hippocampal Volume. J. Gerontol. Ser. B 2020, 75, 1393–1402. [Google Scholar] [CrossRef] [PubMed]
- Nichols, E.; Steinmetz, J.D.; Vollset, S.E.; Fukutaki, K.; Chalek, J.; Abd-Allah, F.; Abdoli, A.; Abualhasan, A.; Abu-Gharbieh, E.; Akram, T.T.; et al. Estimation of the global prevalence of dementia in 2019 and forecasted prevalence in 2050: An analysis for the Global Burden of Disease Study 2019. Lancet Public Health 2022, 7, e105–e125. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Whitsel, E.; Avery, C.; Hughes, T.M.; Griswold, M.E.; Sedaghat, S.; Gottesman, R.F.; Mosley, T.H.; Heiss, G.; Lutsey, P.L. Variation in Population Attributable Fraction of Dementia Associated With Potentially Modifiable Risk Factors by Race and Ethnicity in the US. JAMA Netw. Open 2022, 5, e2219672. [Google Scholar] [CrossRef] [PubMed]
- Baumgart, M.; Snyder, H.M.; Carrillo, M.C.; Fazio, S.; Kim, H.; Johns, H. Summary of the evidence on modifiable risk factors for cognitive decline and dementia: A population-based perspective. Alzheimer’s Dement. 2015, 11, 718–726. [Google Scholar] [CrossRef]
- Yusuf, S.; Joseph, P.; Rangarajan, S.; Islam, S.; Mente, A.; Hystad, P.; Brauer, M.; Kutty, V.R.; Gupta, R.; Wielgosz, A.; et al. Modifiable risk factors, cardiovascular disease, and mortality in 155 722 individuals from 21 high-income, middle-income, and low-income countries (PURE): A prospective cohort study. Lancet 2020, 395, 795–808. [Google Scholar] [CrossRef]
- van den Berg, N.; Rodríguez-Girondo, M.; van Dijk, I.K.; Mourits, R.J.; Mandemakers, K.; Janssens, A.A.P.O.; Beekman, M.; Smith, K.R.; Slagboom, P.E. Longevity defined as top 10% survivors and beyond is transmitted as a quantitative genetic trait. Nat. Commun. 2019, 10, 35. [Google Scholar] [CrossRef]
- Deelen, J.; Evans, D.S.; Arking, D.E.; Tesi, N.; Nygaard, M.; Liu, X.; Wojczynski, M.K.; Biggs, M.L.; van der Spek, A.; Atzmon, G.; et al. A meta-analysis of genome-wide association studies identifies multiple longevity genes. Nat. Commun. 2019, 10, 3669. [Google Scholar] [CrossRef]
- Cherbuin, N.; Walsh, E.; Baune, B.T.; Anstey, K.J. Oxidative stress, inflammation and risk of neurodegeneration in a population sample. Eur. J. Neurol. 2019, 26, 1347–1354. [Google Scholar] [CrossRef]
- Cherbuin, N.; Walsh, E.I.; Leach, L.; Brüstle, A.; Burns, R.; Anstey, K.J.; Sachdev, P.S.; Baune, B.T. Systemic Inflammation Predicts Alzheimer Pathology in Community Samples without Dementia. Biomedicines 2022, 10, 1240. [Google Scholar] [CrossRef]
- King, E.; O’Brien, J.T.; Donaghy, P.; Morris, C.; Barnett, N.; Olsen, K.; Martin-Ruiz, C.; Taylor, J.P.; Thomas, A.J. Peripheral inflammation in prodromal Alzheimer’s and Lewy body dementias. J. Neurol. Neurosurg. Psychiatry 2018, 89, 339–345. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, E.; Morales-Pison, S.; Urbina, F.; Solari, A. Aging Hallmarks and the Role of Oxidative Stress. Antioxidants 2023, 12, 651. [Google Scholar] [CrossRef] [PubMed]
- Maynard, S.; Fang, E.F.; Scheibye-Knudsen, M.; Croteau, D.L.; Bohr, V.A. DNA Damage, DNA Repair, Aging, and Neurodegeneration. Cold Spring Harb. Perspect. Med. 2015, 5, a025130. [Google Scholar] [CrossRef] [PubMed]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Dharmalingam, P.; Vasquez, V.; Mitra, J.; Boldogh, I.; Rao, K.S.; Kent, T.A.; Mitra, S.; Hegde, M.L. Chronic oxidative damage together with genome repair deficiency in the neurons is a double whammy for neurodegeneration: Is damage response signaling a potential therapeutic target? Mech. Ageing Dev. 2017, 161, 163–176. [Google Scholar] [CrossRef]
- Sykora, P.; Wilson, D.M.; Bohr, V.A. Base excision repair in the mammalian brain: Implication for age related neurodegeneration. Mech. Ageing Dev. 2013, 134, 440–448. [Google Scholar] [CrossRef]
- Pezone, A.; Olivieri, F.; Napoli, M.V.; Procopio, A.; Avvedimento, E.V.; Gabrielli, A. Inflammation and DNA damage: Cause, effect or both. Nat. Rev. Rheumatol. 2023, 19, 200–211. [Google Scholar] [CrossRef]
- Zuo, L.; Prather, E.R.; Stetskiv, M.; Garrison, D.E.; Meade, J.R.; Peace, T.I.; Zhou, T. Inflammaging and Oxidative Stress in Human Diseases: From Molecular Mechanisms to Novel Treatments. Int. J. Mol. Sci. 2019, 20, 4472. [Google Scholar] [CrossRef]
- Glassner, B.J.; Rasmussen, L.J.; Najarian, M.T.; Posnick, L.M.; Samson, L.D. Generation of a strong mutator phenotype in yeast by imbalanced base excision repair. Proc. Natl. Acad. Sci. USA 1998, 95, 9997–10002. [Google Scholar] [CrossRef]
- Thompson, B.A.; Spurdle, A.B.; Plazzer, J.P.; Greenblatt, M.S.; Akagi, K.; Al-Mulla, F.; Bapat, B.; Bernstein, I.; Capellá, G.; den Dunnen, J.T.; et al. Application of a 5-tiered scheme for standardized classification of 2360 unique mismatch repair gene variants in the InSiGHT locus-specific database. Nat. Genet 2014, 46, 107–115. [Google Scholar] [CrossRef]
- Cheong, A.; Nagel, Z.D. Human Variation in DNA Repair, Immune Function, and Cancer Risk. Front Immunol. 2022, 13, 899574. [Google Scholar] [CrossRef] [PubMed]
- Siggens, L.; Figg, N.; Bennett, M.; Foo, R. Nutrient deprivation regulates DNA damage repair in cardiomyocytes via loss of the base-excision repair enzyme OGG1. FASEB J. 2012, 26, 2117–2124. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Sowers, J.R.; Zhang, Y.; Ren, J. Targeting DNA damage response in cardiovascular diseases: From pathophysiology to therapeutic implications. Cardiovasc. Res. 2022, 26, 2117–2124. [Google Scholar] [CrossRef] [PubMed]
- Vinokurov, A.Y.; Stelmashuk, O.A.; Ukolova, P.A.; Zherebtsov, E.A.; Abramov, A.Y. Brain region specificity in reactive oxygen species production and maintenance of redox balance. Free Radic. Biol. Med. 2021, 174, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.I.; Duszenko, M.; Gospodaryov, D.V.; Garaschuk, O. Oxidative Stress and Energy Metabolism in the Brain: Midlife as a Turning Point. Antioxidants 2021, 10, 1715. [Google Scholar] [CrossRef]
- Iyama, T.; Wilson, D.M. DNA repair mechanisms in dividing and non-dividing cells. DNA Repair 2013, 12, 620–636. [Google Scholar] [CrossRef] [PubMed]
- Rolseth, V.; Rundén-Pran, E.; Luna, L.; McMurray, C.; Bjørås, M.; Ottersen, O.P. Widespread distribution of DNA glycosylases removing oxidative DNA lesions in human and rodent brains. DNA Repair 2008, 7, 1578–1588. [Google Scholar] [CrossRef]
- Galick, H.A.; Kathe, S.; Liu, M.; Robey-Bond, S.; Kidane, D.; Wallace, S.S.; Sweasy, J.B. Germ-line variant of human NTH1 DNA glycosylase induces genomic instability and cellular transformation. Proc. Natl. Acad. Sci. USA 2013, 110, 14314–14319. [Google Scholar] [CrossRef]
- Kwiatkowski, D.; Czarny, P.; Toma, M.; Jurkowska, N.; Sliwinska, A.; Drzewoski, J.; Bachurska, A.; Szemraj, J.; Maes, M.; Berk, M.; et al. Associations between DNA Damage, DNA Base Excision Repair Gene Variability and Alzheimer’s Disease Risk. Dement. Geriatr. Cogn. Disord. 2016, 41, 152–171. [Google Scholar] [CrossRef]
- Alateeq, K.; Walsh, E.I.; Abhayaratna, W.P.; Cherbuin, N. Effects of Higher Normal Blood Pressure on Brain Are Detectable before Middle-Age and Differ by Sex. J. Clin. Med. 2022, 11, 3127. [Google Scholar] [CrossRef]
- Ambikairajah, A.; Tabatabaei-Jafari, H.; Hornberger, M.; Cherbuin, N. Age, menstruation history, and the brain. Menopause 2020, 28, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Shaw, M.E.; Abhayaratna, W.P.; Anstey, K.J.; Cherbuin, N. Increasing Body Mass Index at Midlife is Associated with Increased Cortical Thinning in Alzheimer’s Disease-Vulnerable Regions. J. Alzheimers. Dis. 2017, 59, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Linzer, D.A.; Lewis, J.B. poLCA: An R Package for Polytomous Variable Latent Class Analysis. J. Stat. Softw. 2011, 42, 1–29. [Google Scholar] [CrossRef]
- Hildrestrand, G.A.; Rolseth, V.; Kunath, N.; Suganthan, R.; Jensen, V.; Bugaj, A.M.; Fernandez-Berrocal, M.S.; Sikko, S.B.; Vetlesen, S.; Kuśnierczyk, A.; et al. NEIL1 and NEIL2 DNA glycosylases modulate anxiety and learning in a cooperative manner in mice. Commun. Biol. 2021, 4, 1354. [Google Scholar] [CrossRef] [PubMed]
- Canugovi, C.; Misiak, M.; Scheibye-Knudsen, M.; Croteau, D.L.; Mattson, M.P.; Bohr, V.A. Loss of NEIL1 causes defects in olfactory function in mice. Neurobiol. Aging 2015, 36, 1007–1012. [Google Scholar] [CrossRef] [PubMed]
- Canugovi, C.; Yoon, J.S.; Feldman, N.H.; Croteau, D.L.; Mattson, M.P.; Bohr, V.A. Endonuclease VIII-like 1 (NEIL1) promotes short-term spatial memory retention and protects from ischemic stroke-induced brain dysfunction and death in mice. Proc. Natl. Acad. Sci. USA 2012, 109, 14948–14953. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.Y.; Lin, Y.C.; Calkins, M.J.; Chen, P.C. Endonuclease VIII-like 1 deficiency impairs survival of newly generated hippocampal neurons and memory performance in young-adult male mice. Life Sci. 2020, 254, 117755. [Google Scholar] [CrossRef]
- Ertuzun, T.; Semerci, A.; Cakir, M.E.; Ekmekcioglu, A.; Gok, M.O.; Soltys, D.T.; de Souza-Pinto, N.C.; Sezerman, U.; Muftuoglu, M. Investigation of base excision repair gene variants in late-onset Alzheimer’s disease. PLoS ONE 2019, 14, e0221362. [Google Scholar] [CrossRef]
- Sliwinska, A.; Sitarek, P.; Toma, M.; Czarny, P.; Synowiec, E.; Krupa, R.; Wigner, P.; Bialek, K.; Kwiatkowski, D.; Korycinska, A.; et al. Decreased expression level of BER genes in Alzheimer’s disease patients is not derivative of their DNA methylation status. Prog. Neuropsychopharmacol. Biol. Psychiatry 2017, 79 Pt B, 311–316. [Google Scholar] [CrossRef]
- Egiazarian, M.A.; Strømstad, S.; Sakshaug, T.; Nunez-Nescolarde, A.B.; Bethge, N.; Bjørås, M.; Scheffler, K. Age- and sex-dependent effects of DNA glycosylase Neil3 on amyloid pathology, adult neurogenesis, and memory in a mouse model of Alzheimer’s disease. Free Radic. Biol. Med. 2022, 193, 685–693. [Google Scholar] [CrossRef]
- Cheriyan, J.; Kim, S.; Wolansky, L.J.; Cook, S.D.; Cadavid, D. Impact of Inflammation on Brain Volume in Multiple Sclerosis. Arch. Neurol. 2012, 69, 82–88. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Genes | Function in Base Excision Repair | SNPs |
|---|---|---|
| OGG1 | Detection and excision of pyrimidines in double-stranded DNA, including the most frequently occurring oxidized DNA lesion, 8-oxoguanine. | rs1052133 rs104893751 |
| NEIL1 | Detection and excision of pyrimidines, 8-oxoguanine, and formamidopyrimidine from both single and double-stranded DNA, preferably in bubble-structured DNA, as well as in close proximity to another DNA lesion. Contributes particularly to transcription-associated DNA repair. | rs7402844 rs5745906 |
| NEIL2 | Similar to NEIL1, however, its expression is cell-cycle independent with a particular affinity for cytosine-derived lesions such as 5-hydroxyuracil. | rs6601606 |
| NEIL3 | Similar to NEIL1 and NEIL2, but mostly expressed in development, e.g., in brain regions rich in progenitor cells (subventricular zone, hippocampus, cerebellum), decreasing with age. | rs10013040 rs13112390 rs13112358 rs1395479 |
| MUTYH | Provides protection against the mispairing of adenosine with 8-oxoguanine by removing the adenosine base and prevents the accumulation of 8-oxoguanine lesions. MUTYH can also remove other sources of lesions, such as oxidized adenines. | rs34612342 rs200165598 |
| NHTHL1 | Detection and excision of pyrimidines and purines. Specific protective action against telomeric lesions. | rs150766139 rs2516739 |
| Female [n = 264,576] | Male [n = 223,437] | Total [n = 488,013] | |
|---|---|---|---|
| Age (years) | |||
| Mean (SD) | 56.36 (8.00) | 56.75 (8.20) | 56.54 (8.09) |
| Education (highest qualification) | |||
| Primary | 44,727 (16.9%) | 38,546 (17.3%) | 83,273 (17.1%) |
| Secondary | 107,069 (40.5%) | 76,355 (34.2%) | 183,424 (37.6%) |
| Prof certificate/diploma | 27,067 (10.2%) | 30,204 (13.5%) | 57,271 (11.7%) |
| Tertiary | 82,457 (31.2%) | 75,318 (33.7%) | 157,775 (32.3%) |
| Unknown | 3256 (1.2%) | 3014 (1.3%) | 6270 (1.3%) |
| Smoke (ever) | |||
| No | 117,855 (44.8%) | 77,159 (34.7%) | 195,014 (40.2%) |
| Yes | 145,436 (55.2%) | 145,119 (65.3%) | 290,555 (59.8%) |
| Alcohol | |||
| Current | 238,953 (90.3%) | 208,754 (93.4%) | 447,707 (91.7%) |
| Never | 15,359 (5.8%) | 6193 (2.8%) | 21,552 (4.4%) |
| Past | 9631 (3.6%) | 7893 (3.5%) | 17,524 (3.6%) |
| Unknown | 633 (0.2%) | 597 (0.3%) | 1230 (0.3%) |
| BMI (kg/m2) | |||
| Mean (SD) | 27.07 (5.18) | 27.83 (4.24) | 27.42 (4.79) |
| Range | 12.12–74.68 | 12.81–68.41 | 12.12–74.68 |
| Diabetes | |||
| No | 253,463 (95.8%) | 206,618 (92.5%) | 460,081 (94.3%) |
| Yes | 10,177 (3.8%) | 15,718 (7.0%) | 25,895 (5.3%) |
| Unknown | 936 (0.4%) | 1101 (0.5%) | 2037 (0.4%) |
| Cognitive Measures | |||||
|---|---|---|---|---|---|
| SNP Variants (1/2 Alleles vs. None) | FIQ | SDMT | MATCH | TRAIL1 | TRAIL2 |
| rs1052133(1) | 0.669 | ||||
| p = 0.052 | |||||
| rs1052133(2) | 2.322 ** | ||||
| p = 0.002 | |||||
| rs7402844(1) | 0.080 ** | −8.690 *** | −0.661 * | ||
| p = 0.001 | p < 0.00001 | p = 0.034 | |||
| rs7402844(2) | 0.102 *** | −10.890 *** | −0.921 ** | ||
| p = 0.00001 | p = 0.000 | p = 0.003 | |||
| rs6601606(1) | −0.187 *** | 6.889 *** | |||
| p < 0.00001 | p < 0.00001 | ||||
| rs6601606(2) | −1.559 *** | 24.501 ** | |||
| p < 0.00001 | p = 0.0004 | ||||
| rs13112358(1) | 0.081 ** | −2.537 ** | |||
| p = 0.001 | p = 0.0004 | ||||
| rs13112358(2) | 0.094 ** | −4.148 *** | |||
| p = 0.0001 | p < 0.00001 | ||||
| rs2516739(1) | −0.003 | 0.899 * | |||
| p = 0.811 | p = 0.010 | ||||
| rs2516739(2) | −0.079 ** | 5.669 *** | |||
| p = 0.003 | p < 0.00001 | ||||
| rs1395479(1) | 0.115 ** | −2.809 *** | |||
| p = 0.0001 | p < 0.00001 | ||||
| rs1395479(2) | 0.077 | −3.556 *** | |||
| p = 0.163 | p < 0.00001 | ||||
| rs200165598(1) | 18.339 ** | ||||
| p = 0.003 | |||||
| Constant | 3.536 *** | 16.337 *** | 614.845 *** | 47.156 *** | 87.129 *** |
| p < 0.00001 | p < 0.00001 | p < 0.00001 | p < 0.00001 | p < 0.00001 | |
| Observations | 120,453 | 115,893 | 478,185 | 101,909 | 101,788 |
| Log Likelihood | −248,761 | −342,715 | −2,935,306 | −415,735 | −466,588 |
| Akaike Inf. Crit. | 497,552 | 685,449 | 5,870,652 | 831,487 | 933,196 |
| Dependent Variable | MATCH | ||
|---|---|---|---|
| Protect | Harm | Risk | |
| Age (years) | 3.136 *** | 3.668 *** | 3.978 *** |
| p < 0.00001 | p < 0.00001 | p < 0.00001 | |
| Sex (Male) | −16.503 *** | −18.113 *** | −17.666 *** |
| p < 0.00001 | p < 0.00001 | p < 0.00001 | |
| Index | −2.050 *** | 1.315 *** | 1.684 *** |
| p < 0.00001 | p = 0.00005 | p < 0.00001 | |
| Age × Index | 0.135 *** | −0.75 *** | −0.109 *** |
| p < 0.00001 | p = 0.008 | p < 0.00001 | |
| Sex × Index | −0.387 | 0.162 | 0.364 |
| p = 0.295 | p = 0.722 | p = 0.204 | |
| Constant | 609.515 *** | 600.920 *** | 596.709 *** |
| p < 0.00001 | p < 0.00001 | p < 0.00001 | |
| Observations | 476,240 | 480,300 | 473,791 |
| Log Likelihood | −2,923,622 | −2,948,533 | −2,908,645 |
| Akaike Inf. Crit. | 5,847,265 | 5,897,086 | 5,817,310 |
| Brain Volumes | ||||||
|---|---|---|---|---|---|---|
| SNP Variants (1/2 Alleles vs. None) | LHC | RHC | GM | WM | WMH | |
| Age (years) | −12.031 *** | −12.582 *** | −753.194 *** | −138.614 *** | 135.550 *** | |
| p < 0.00001 | p < 0.00001 | p < 0.00001 | p < 0.00001 | p < 0.00001 | ||
| Sex (Male) | 3.885 | −14.718 *** | 3426.919 *** | −4741.967 *** | −163.435 *** | |
| p = 0.321 | p = 0.0004 | p < 0.00001 | p < 0.00001 | p < 0.00001 | ||
| rs34612342(1) | 43.861 * | 34.667 | 2674.997 * | |||
| p = 0.052 | p = 0.145 | p = 0.071 | ||||
| rs200495564(1) | −472.157 | −137.377 | ||||
| p = 0.126 | p = 0.674 | |||||
| rs13112390(1) | −2476.289 *** | |||||
| p = 0.009 | ||||||
| rs13112390(2) | −3008.684 *** | |||||
| p = 0.004 | ||||||
| rs13112358(1) | 1914.805 ** | |||||
| p = 0.022 | ||||||
| rs13112358(2) | 2233.274 ** | |||||
| p = 0.019 | ||||||
| rs6601606(1) | −40.886 | |||||
| p = 0.598 | ||||||
| rs6601606(2) | 1219.892 * | |||||
| p = 0.059 | ||||||
| Constant | 1473.881 *** | 1503.877 *** | 104,839.200 *** | −81,903.510 *** | −2138.101 *** | |
| p < 0.00001 | p < 0.00001 | p < 0.00001 | p < 0.00001 | p < 0.00001 | ||
| Observations | 38,991 | 38,991 | 38,781 | 39,025 | 39,044 | |
| Log Likelihood | −278,780 | −280,983 | −438,138 | −442,333.300 | −362,143 | |
| Akaike Inf. Crit. | 557,572 | 561,979 | 876,293 | 884,676.600 | 724,299 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cherbuin, N.; Patel, H.; Walsh, E.I.; Ambikairajah, A.; Burns, R.; Brüstle, A.; Rasmussen, L.J. Cognitive Function Is Associated with the Genetically Determined Efficiency of DNA Repair Mechanisms. Genes 2024, 15, 153. https://doi.org/10.3390/genes15020153
Cherbuin N, Patel H, Walsh EI, Ambikairajah A, Burns R, Brüstle A, Rasmussen LJ. Cognitive Function Is Associated with the Genetically Determined Efficiency of DNA Repair Mechanisms. Genes. 2024; 15(2):153. https://doi.org/10.3390/genes15020153
Chicago/Turabian StyleCherbuin, Nicolas, Hardip Patel, Erin I. Walsh, Ananthan Ambikairajah, Richard Burns, Anne Brüstle, and Lene Juel Rasmussen. 2024. "Cognitive Function Is Associated with the Genetically Determined Efficiency of DNA Repair Mechanisms" Genes 15, no. 2: 153. https://doi.org/10.3390/genes15020153
APA StyleCherbuin, N., Patel, H., Walsh, E. I., Ambikairajah, A., Burns, R., Brüstle, A., & Rasmussen, L. J. (2024). Cognitive Function Is Associated with the Genetically Determined Efficiency of DNA Repair Mechanisms. Genes, 15(2), 153. https://doi.org/10.3390/genes15020153