
Comprehensive Assessment of Reference Gene Expression within the Whitefly Dialeurodes citri Using RT-qPCR
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Samples of Citrus Whitefly
2.3. Total RNA Extraction and cDNA Synthesis
2.4. Gene Cloning and Primer Design
2.5. RT-qPCR of Each Reference Gene
2.6. Determination of Reference Gene Expression
2.7. Gene Expression Level Analysis Using Various Reference Genes
3. Results
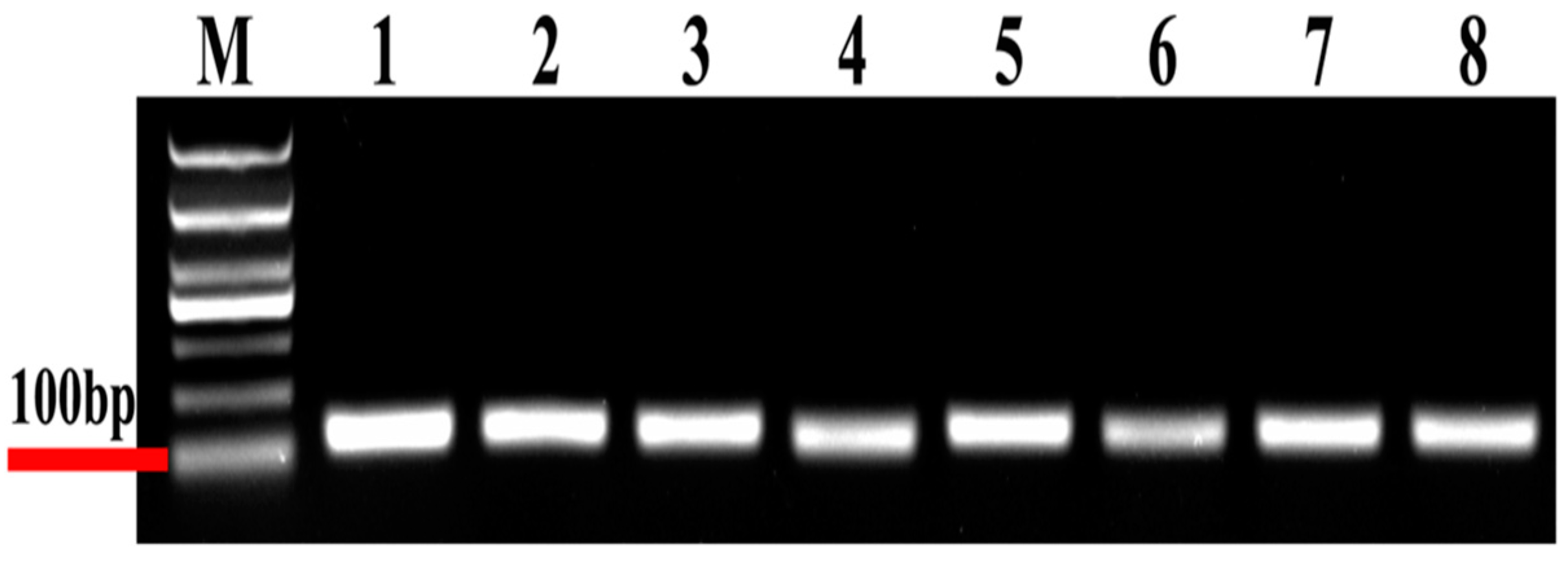
3.1. PCR Amplification and Performance of Candidate Reference Genes in D. citri
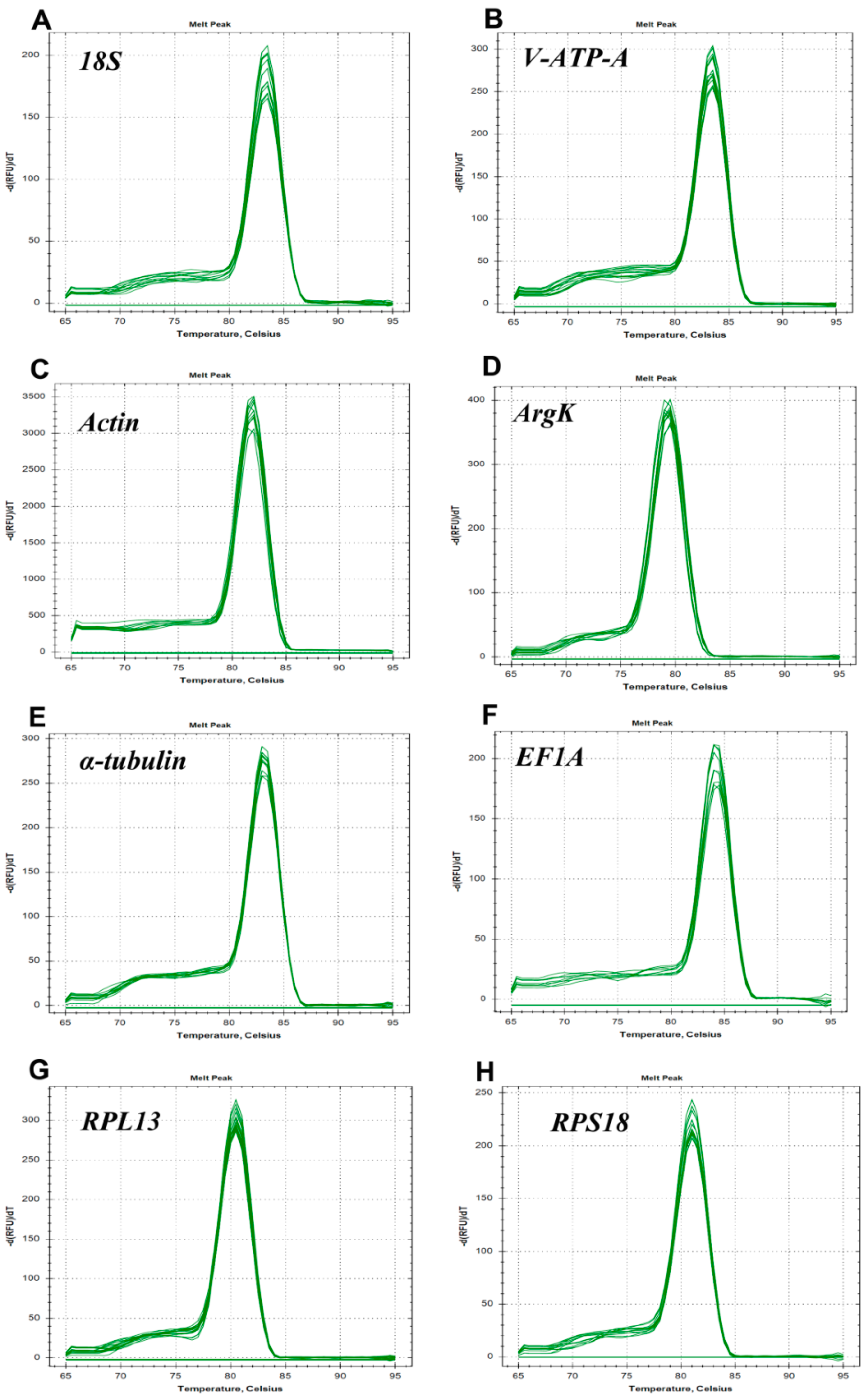
3.2. Expression Levels of Reference Genes
3.3. Stability of the Reference Genes under Specific Experimental Conditions
3.4. Stability Ranking of The Eight Reference Genes
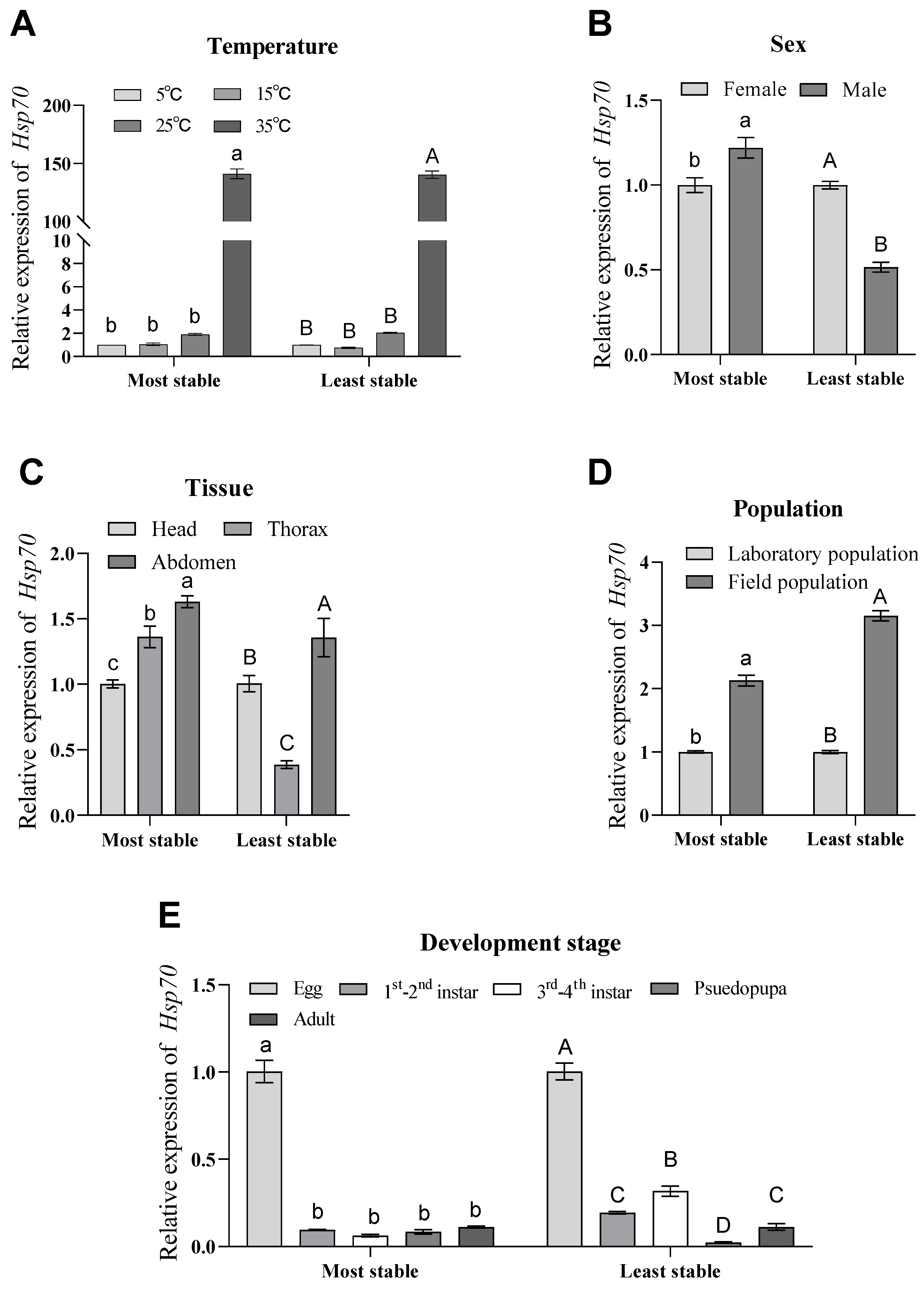
3.5. Validation of the Selected Reference Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Argov, Y.; Rössler, Y.; Voet, H.; Rosen, D. Spatial Dispersion and Sampling of Citrus Whitefly, Dialeurodes citri, for Control Decisions in a Citrus Orchard. Agric. For. Entomol. 1999, 1, 305–318. [Google Scholar] [CrossRef]
- Bellows, T.S.; Meisenbacher, C. Field Population Biology of Citrus Whitefly, Dialeurodes citri (Ashmead) (Heteroptera: Aleyrodidae), on Oranges in California. Popul. Ecol. 2007, 49, 127–134. [Google Scholar] [CrossRef]
- Martin, J.H.; Mifsud, D.; Rapisarda, C. The Whiteflies (Hemiptera: Aleyrodidae) of Europe and the Mediterranean Basin. Bull. Entomol. Res. 2000, 90, 407–448. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.H.; Liu, C.H.; Wang, Q.; Wang, Y.L.; Zhou, C.Y.; Zhou, Y. Identification of Dialeurodes citri as a Vector of Citrus Yellow Vein Clearing Virus in China. Plant Dis. 2019, 103, 65–68. [Google Scholar] [CrossRef]
- Wang, P.P.; Song, X.H.; Zhang, H.Y. Isolation and Characterization of Aschersonia placenta from Citrus Orchards and Its Pathogenicity Towards Dialeurodes citri (Ashmead). J. Invertebr. Pathol. 2013, 112, 122–128. [Google Scholar] [CrossRef]
- Kunimi, Y. Current Status and Prospects on Microbial Control in Japan. J. Invertebr. Pathol. 2007, 95, 181–186. [Google Scholar] [CrossRef]
- Chen, C.-Y.; Lai, C.-Y.; Kuo, M.-H. Temperature Effect on the Growth of Buchnera Endosymbiont in Aphis craccivora (Hemiptera: Aphididae). Symbiosis 2009, 49, 53–59. [Google Scholar] [CrossRef]
- Ferguson, B.S.; Nam, H.; Hopkins, R.G.; Morrison, R.F. Impact of Reference Gene Selection for Target Gene Normalization on Experimental Outcome Using Real-Time qRT-PCR in Adipocytes. PLoS ONE 2010, 5, e15208. [Google Scholar] [CrossRef]
- Li, R.; Xie, W.; Wang, S.; Wu, Q.; Yang, N.; Yang, X.; Pan, H.; Zhou, X.; Bai, L.; Xu, B.; et al. Reference Gene Selection for qRT-PCR Analysis in the Sweetpotato Whitefly, Bemisia Tabaci (Hemiptera: Aleyrodidae). PLoS ONE 2013, 8, e53006. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.; Guo, Y.; Zhou, X.; Gao, X. Expression Profiling in Bemisia tabaci under Insecticide Treatment: Indicating the Necessity for Custom Reference Gene Selection. PLoS ONE 2014, 9, e87514. [Google Scholar] [CrossRef]
- Suzuki, T.; Higgins, P.J.; Crawford, D.R. Control Selection for RNA Quantitation. Biotechniques 2000, 29, 332–337. [Google Scholar] [CrossRef]
- Huggett, J.; Dheda, K.; Bustin, S.; Zumla, A. Real-Time RT-qPCR Normalisation: Strategies and Considerations. Genes Immun. 2005, 6, 279–284. [Google Scholar] [CrossRef]
- Lv, Z.H.; Pan, H.P.; Zhang, W.; Ding, T.B.; Chu, D. Reference Gene Selection for RT-qPCR Analysis in Two Invasive Whiteflies after the Acquisition of Vectored or Non-Vectored Viruses. J. Asia Pac. Entomol. 2018, 21, 19–24. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-qPCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, 0034.1. [Google Scholar] [CrossRef] [PubMed]
- Gachon, C.; Mingam, A.; Charrier, B. Real-Time PCR: What Relevance to Plant Studies? J. Exp. Bot. 2004, 55, 1445–1454. [Google Scholar] [CrossRef]
- Hellemans, J.; Vandesompele, J. Selection of Reliable Reference Genes for RT-qPCR Analysis. Quant. Real Time PCR Methods Protoc. 2014, 1160, 19–26. [Google Scholar] [CrossRef]
- Pan, H.P.; Yang, X.W.; Siegfried, B.D.; Zhou, X.G. A Comprehensive Selection of Reference Genes for RT-qPCR Analysis in a Predatory Lady Beetle, Hippodamia convergens (Coleoptera Coccinellidae). PLoS ONE 2015, 10, e0125868. [Google Scholar] [CrossRef]
- Guo, C.-F.; Pan, H.-P.; Zhang, L.-H.; Ou, D.; Lu, Z.-T.; Khan, M.M.; Qiu, B.-L. Comprehensive Assessment of Candidate Reference Genes for Gene Expression Studies Using RT-qPCR in Tamarixia radiata, a Predominant Parasitoid of Diaphorina citri. Genes 2020, 11, 1178. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Qiu, J.; Hu, Y.; Xu, P.; Deng, Y.; Tian, L.; Wei, Y.; Sang, W.; Liu, Y.; Qiu, B. Silencing of V-ATPase-E Gene Causes Midgut Apoptosis of Diaphorina citri and Affects Its Acquisition of Huanglongbing Pathogen. Insect Sci. 2023, 30, 1022–1034. [Google Scholar] [CrossRef]
- Khan, M.M.; Guo, C.-F.; Peng, J.; Fan, Z.-Y.; Hafeez, M.; Ali, D.; Wang, K.; Almarzoug, M.H.; Qiu, B.-L. Screening and Validation of Reference Genes Using in RT-qPCR for Gene Expression Studies in Paederus fuscipes, a Medically and Agriculturally Important Insect. J. King Saud Univ. Sci. 2022, 34, 101654. [Google Scholar] [CrossRef]
- Liu, D.W.; Chen, S.T.; Liu, H.P. Choice of Endogenous Control for Gene Expression in Nonsmall Cell Lung Cancer. Eur. Respir. J. 2005, 26, 1002–1008. [Google Scholar] [CrossRef]
- Lü, J.; Chen, S.; Guo, M.; Ye, C.; Qiu, B.; Wu, J.; Yang, C.; Pan, H. Selection and Validation of Reference Genes for RT-qPCR Analysis of the Ladybird Beetle Henosepilachna vigintioctomaculata. Front. Physiol. 2018, 9, 1614. [Google Scholar] [CrossRef]
- Lü, J.; Yang, C.X.; Zhang, Y.J.; Pan, H.P. Selection of Reference Genes for the Normalization of RT-qPCR Data in Gene Expression Studies in Insects: A Systematic Review. Front. Physiol. 2018, 9, 1560. [Google Scholar] [CrossRef]
- Pan, H.P.; Yang, X.W.; Bidne, K.; Hellmich, R.L.; Siegfried, B.D.; Zhou, X.G. Selection of Reference Genes for RT-qPCR Analysis in the Monarch Butterfly, Danaus plexippus (L.), a Migrating Bio-Indicator. PLoS ONE 2015, 10, e0129482. [Google Scholar] [CrossRef] [PubMed]
- Volkov, R.A.; Panchuk, L.L.; Schöffl, F. Heat-Stress-Dependency and Developmental Modulation of Gene Expression: The Potential of House-Keeping Genes as Internal Standards in mRNA Expression Profiling Using Real-Time RT-qPCR. J. Exp. Bot. 2003, 54, 2343–2349. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.X.; Pan, H.P.; Liu, Y.; Zhou, X.G. Selection of Reference Genes for Expression Analysis Using Quantitative Real-Time PCR in the Pea Aphid, Acyrthosiphon pisum (Harris) (Hemiptera, Aphidiae). PLoS ONE 2014, 9, e110454. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Pan, H.; Noland, J.E.; Zhang, D.; Zhang, Z.; Liu, Y.; Zhou, X. Selection of Reference Genes for RT-qPCR Analysis in a Predatory Biological Control Agent, Coleomegilla maculata (Coleoptera: Coccinellidae). Sci. Rep. 2015, 5, 18201. [Google Scholar] [CrossRef]
- Yang, X.W.; Pan, H.P.; Yuan, L.; Zhou, X.G. Reference Gene Selection for RT-qPCR Analysis in Harmonia axyridis, a Global Invasive Lady Beetle. Sci. Rep. 2018, 8, 2689. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.F.; Lin, Q.C.; Zhou, X.H.; Zhang, X.Y.; Liu, T.L.; Yu, Y. Identification and Validation of Reference Genes for Quantitative Real-Time PCR in Drosophila suzukii (Diptera Drosophilidae). PLoS ONE 2014, 9, e106800. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The Miqe Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Selvey, S.; Thompson, E.W.; Matthaei, K.; Lea, R.A.; Irving, M.G.; Griffiths, L.R. β-Actin-an Unsuitable Internal Control for RT-qPCR. Mol. Cell. Probes 2001, 15, 307–311. [Google Scholar] [CrossRef]
- Yuan, M.; Lu, Y.; Zhu, X.; Wan, H.; Shakeel, M.; Zhan, S.; Jin, B.-R.; Li, J. Selection and Evaluation of Potential Reference Genes for Gene Expression Analysis in the Brown Planthopper, Nilaparvata lugens (Hemiptera: Delphacidae) Using Reverse-Transcription Quantitative PCR. PLoS ONE 2014, 9, e86503. [Google Scholar] [CrossRef]
- Dai, T.M.; Lü, Z.C.; Liu, W.X.; Wan, F.H. Selection and Validation of Reference Genes for qRT-PCR Analysis During Biological Invasions: The Thermal Adaptability of Bemisia tabaci Med. PLoS ONE 2017, 12, e0173821. [Google Scholar] [CrossRef]
- Ni, M.; Li, Z.; Li, J.; He, H.; Wang, Y.; Jiang, Y.; Wang, X.; Li, Z.; Li, M.; Xu, H. Selection and Validation of Reference Genes for the Normalization of Quantitative Real-Time PCR in Different Muscle Tissues of Rabbits. BMC Zool. 2022, 7, 60. [Google Scholar] [CrossRef]
- Wieczorek, H.; Huss, M.; Merzendorfer, H.; Reineke, S.; Vitavska, O.; Zeiske, W. The Insect Plasma Membrane H+V-Atpase: Intra-, Inter-, and Supramolecular Aspects. J. Bioenerg. Biomembr. 2003, 35, 359–366. [Google Scholar] [CrossRef]
- Lu, Y.; Yuan, M.; Gao, X.; Kang, T.; Zhan, S.; Wan, H.; Li, J. Identification and Validation of Reference Genes for Gene Expression Analysis Using Quantitative PCR in Spodoptera litura (Lepidoptera: Noctuidae). PLoS ONE 2013, 8, e68059. [Google Scholar] [CrossRef]
- Sun, M.; Lu, M.-X.; Tang, X.-T.; Du, Y.-Z. Exploring Valid Reference Genes for Quantitative Real-Time PCR Analysis in Sesamia inferens (Lepidoptera: Noctuidae). PLoS ONE 2015, 10, e0115979. [Google Scholar] [CrossRef]
- Zhu, X.; Yuan, M.; Shakeel, M.; Zhang, Y.; Wang, S.; Wang, X.; Zhan, S.; Kang, T.; Li, J. Selection and Evaluation of Reference Genes for Expression Analysis Using qRT-PCR in the Beet Armyworm Spodoptera exigua (Hübner) (Lepidoptera: Noctuidae). PLoS ONE 2014, 9, e84730. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of Real-Time Quantitative Reverse Transcription-PCR Data: A Model-Based Variance Estimation Approach to Identify Genes Suited for Normalization, Applied to Bladder and Colon Cancer Data Sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of Stable Housekeeping Genes, Differentially Regulated Target Genes and Sample Integrity: Bestkeeper—Excel-Based Tool Using Pair-Wise Correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Silver, N.; Best, S.; Jiang, J.; Thein, S.L. Selection of Housekeeping Genes for Gene Expression Studies in Human Reticulocytes Using Real-Time PCR. BMC Mol. Biol. 2006, 7, 33. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔct Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Preisser, E.L.; Zhang, H.; Liu, Y.; Dai, L.; Pan, H.; Zhou, X. Selection of Reference Genes for RT-qPCR Analysis in Coccinella septempunctata to Assess Un-Intended Effects of RNAi Transgenic Plants. Front. Plant Sci. 2016, 7, 1672. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.X.; Pan, H.P.; Liu, Y.; Zhou, X.G. Temperature and Development Impacts on Housekeeping Gene Expression in Cowpea Aphid, Aphis craccivora (Hemiptera Aphidiae). PLoS ONE 2015, 10, e0130593. [Google Scholar] [CrossRef] [PubMed]
- Basnet, S.; Kamble, S.T. RNAi-Mediated Knockdown of vATPase Subunits Affects Survival and Reproduction of Bed Bugs (Hemiptera: Cimicidae). J. Med. Entomol. 2018, 55, 540–546. [Google Scholar] [CrossRef]
- Morales, M.A.; Mendoza, B.M.; Lavine, L.C.; Lavine, M.D.; Walsh, D.B.; Zhu, F. Selection of Reference Genes for Expression Studies of Xenobiotic Adaptation in Tetranychus urticae. Int. J. Biol. Sci. 2016, 12, 1129–1139. [Google Scholar] [CrossRef]
- Sun, W.; Jin, Y.; He, L.; Lu, W.C.; Li, M. Suitable Reference Gene Selection for Different Strains and Developmental Stages of the Carmine Spider Mite, Tetranychus cinnabarinus, Using Quantitative Real-Time PCR. J. Insect Sci. 2010, 10, 208. [Google Scholar] [CrossRef]
- Gao, X.-K.; Zhang, S.; Luo, J.-Y.; Wang, C.-Y.; Lü, L.-M.; Zhang, L.-J.; Zhu, X.-Z.; Wang, L.; Lu, H.; Cui, J.-J. Comprehensive Evaluation of Candidate Reference Genes for Gene Expression Studies in Lysiphlebia japonica (Hymenoptera: Aphidiidae) Using RT-qPCR. Gene 2017, 637, 211–218. [Google Scholar] [CrossRef]
- Morammazi, S.; Shokrollahi, B. The Pattern of Hsp70 Gene Expression, Flight Activity and Temperature in Apis Mellifera Meda Colonies. J. Therm. Biol. 2020, 91, 102647. [Google Scholar] [CrossRef]
- Pan, L.N.; Wang, F.Z.; Zhang, X.Y.; Zhao, Y.N.; Zhu, G.P.; Li, M. Identification and Characterization of Heat Shock Proteins in a Parasitic Wasp Chouioia cuneae (Hymenoptera: Eulophidae). Entomol. Res. 2018, 48, 145–155. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.; et al. The Need for Transparency and Good Practices in the qPCR Literature. Nat. Methods 2013, 10, 1063–1067. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Xie, W.; Zhang, Z.; Wang, S.; Wu, Q.; Liu, Y.; Zhou, X.; Zhou, X.; Zhang, Y. Exploring Valid Reference Genes for Quantitative Real-Time PCR Analysis in Plutella xylostella (Lepidoptera: Plutellidae). Int. J. Biol. Sci. 2013, 9, 792. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′-3′) | Length (bp) | Efficiency (%) | R2 | Linear Regression |
|---|---|---|---|---|---|
| 18S-F | GACGGGGTCTCGTTCGTTAT | 119 | 99.86 | 0.9996 | y = −3.3254x + 11.837 |
| 18S-R | CACCGGAAGGATTGACAGAT | ||||
| V-ATP-A-F | GTCAGGGTTGCCAAGACACT | 118 | 103.39 | 0.9965 | y = −3.2433x + 23.339 |
| V-ATP-A-R | AATTTCAGGACGTTTGGCTG | ||||
| Actin-F | AGAAGCCCTCTTCCAACCAT | 127 | 100.37 | 0.9998 | y = −3.313x + 19.51 |
| Actin-R | GGAGAGAACTGTGTTGGCGT | ||||
| ArgK-F | GCAACACCTGATTGGATGAC | 118 | 97.49 | 0.9988 | y = −3.3836x + 20.294 |
| ArgK-R | TTTAGATGCCATCCAGCCAC | ||||
| α-tubulin-F | GGAGGAAACAATTTGACCGA | 126 | 94.07 | 0.9996 | y = −3.4727x + 20.03 |
| α-tubulin-R | GAACACTCCGACTGTGCCTT | ||||
| EF1A-F | TTTAGATGCCATCCAGCCAC | 120 | 91.12 | 0.9991 | y = −3.5548x + 17.789 |
| EF1A-R | ACACCGGTTTCAACACGAC | ||||
| RPL13-F | CGTTTGCAGTGATAGCACGA | 120 | 95.09 | 0.9990 | y = −3.4455x + 18.943 |
| RPL13-R | AGCTGAAGAAGGGAGAGGCT | ||||
| RPS18-F | CTCACAGATCACAGCCTCCA | 108 | 91.79 | 0.9992 | y = −3.5357x + 19.349 |
| RPS18-R | ACACGGAGTCCCCAATGAT |
| Conditions | Rank | geNorm | NormFinder | BestKeeper | ΔCt | ||||
|---|---|---|---|---|---|---|---|---|---|
| Gene | Stability | Gene | Stability | Gene | Stability | Gene | Stability | ||
| Developmental stage | 1 | V-ATP-A | 0.315 | V-ATP-A | 0.158 | 18S | 0.29 | V-ATP-A | 0.82 |
| 2 | RPS18 | 0.315 | RPL13 | 0.306 | α-tubulin | 0.66 | RPS18 | 0.84 | |
| 3 | RPL13 | 0.348 | RPS18 | 0.362 | RPS18 | 0.66 | RPL13 | 0.87 | |
| 4 | EF1A | 0.398 | α-tubulin | 0.567 | RPL13 | 0.82 | α-tubulin | 0.96 | |
| 5 | α-tubulin | 0.464 | EF1A | 0.697 | EF1A | 0.82 | EF1A | 0.99 | |
| 6 | 18S | 0.689 | 18S | 1.124 | V-ATP-A | 0.87 | 18S | 1.36 | |
| 7 | ArgK | 0.965 | ArgK | 1.41 | Actin | 1.61 | ArgK | 1.61 | |
| 8 | Actin | 1.139 | Actin | 1.484 | ArgK | 1.69 | Actin | 1.66 | |
| Sex | 1 | α-tubulin | 0.131 | V-ATP-A | 0.145 | V-ATP-A | 0.06 | 18S | 0.95 |
| 2 | RPS18 | 0.131 | 18S | 0.145 | 18S | 0.22 | α-tubulin | 0.96 | |
| 3 | RPL13 | 0.374 | α-tubulin | 0.401 | α-tubulin | 0.7 | RPS18 | 0.99 | |
| 4 | EF1A | 0.423 | RPS18 | 0.569 | RPS18 | 0.79 | V-ATP-A | 1.03 | |
| 5 | 18S | 0.594 | RPL13 | 1.124 | Actin | 1.12 | RPL13 | 1.24 | |
| 6 | V-ATP-A | 0.716 | EF1A | 1.277 | ArgK | 1.17 | EF1A | 1.34 | |
| 7 | Actin | 1.078 | Actin | 1.654 | RPL13 | 1.18 | Actin | 1.72 | |
| 8 | ArgK | 1.251 | ArgK | 1.721 | EF1A | 1.29 | ArgK | 1.77 | |
| Tissue | 1 | α-tubulin | 0.175 | EF1A | 0.225 | ArgK | 0.18 | EF1A | 0.53 |
| 2 | EF1A | 0.175 | 18S | 0.236 | EF1A | 0.21 | α-tubulin | 0.54 | |
| 3 | RPL13 | 0.256 | α-tubulin | 0.296 | α-tubulin | 0.25 | 18S | 0.56 | |
| 4 | 18S | 0.309 | ArgK | 0.346 | V-ATP-A | 0.31 | RPL13 | 0.59 | |
| 5 | ArgK | 0.41 | RPL13 | 0.38 | 18S | 0.36 | ArgK | 0.61 | |
| 6 | V-ATP-A | 0.453 | V-ATP-A | 0.554 | RPL13 | 0.42 | V-ATP-A | 0.7 | |
| 7 | RPS18 | 0.564 | RPS18 | 0.764 | Actin | 0.72 | RPS18 | 0.87 | |
| 8 | Actin | 0.676 | Actin | 0.924 | RPS18 | 0.73 | Actin | 1.01 | |
| Population | 1 | RPL13 | 0.118 | ArgK | 0.077 | 18S | 0.1 | Actin | 0.27 |
| 2 | V-ATP-A | 0.118 | Actin | 0.084 | ArgK | 0.15 | V-ATP-A | 0.3 | |
| 3 | RPS18 | 0.149 | RPS18 | 0.189 | Actin | 0.27 | ArgK | 0.31 | |
| 4 | EF1A | 0.172 | 18S | 0.193 | α-tubulin | 0.36 | RPS18 | 0.31 | |
| 5 | Actin | 0.183 | V-ATP-A | 0.202 | RPS18 | 0.37 | EF1A | 0.33 | |
| 6 | ArgK | 0.218 | EF1A | 0.233 | EF1A | 0.4 | 18S | 0.36 | |
| 7 | 18S | 0.255 | RPL13 | 0.334 | V-ATP-A | 0.41 | RPL13 | 0.37 | |
| 8 | α-tubulin | 0.375 | α-tubulin | 0.727 | RPL13 | 0.5 | α-tubulin | 0.74 | |
| Temperature | 1 | α-tubulin | 0.115 | RPL13 | 0.114 | RPS18 | 0.08 | 18S | 0.29 |
| 2 | RPS18 | 0.115 | 18S | 0.116 | α-tubulin | 0.08 | RPL13 | 0.29 | |
| 3 | RPL13 | 0.158 | RPS18 | 0.174 | RPL13 | 0.12 | RPS18 | 0.3 | |
| 4 | 18S | 0.192 | α-tubulin | 0.184 | 18S | 0.19 | α-tubulin | 0.3 | |
| 5 | EF1A | 0.235 | EF1A | 0.194 | V-ATP-A | 0.23 | EF1A | 0.32 | |
| 6 | ArgK | 0.248 | ArgK | 0.208 | EF1A | 0.3 | ArgK | 0.32 | |
| 7 | V-ATP-A | 0.301 | V-ATP-A | 0.444 | ArgK | 0.34 | V-ATP-A | 0.48 | |
| 8 | Actin | 0.349 | Actin | 0.461 | Actin | 0.48 | Actin | 0.49 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, W.; Lv, X.; Ran, X.; Mukangango, M.; Eric Derrick, B.; Qiu, B.; Guo, C. Comprehensive Assessment of Reference Gene Expression within the Whitefly Dialeurodes citri Using RT-qPCR. Genes 2024, 15, 318. https://doi.org/10.3390/genes15030318
Kong W, Lv X, Ran X, Mukangango M, Eric Derrick B, Qiu B, Guo C. Comprehensive Assessment of Reference Gene Expression within the Whitefly Dialeurodes citri Using RT-qPCR. Genes. 2024; 15(3):318. https://doi.org/10.3390/genes15030318
Chicago/Turabian StyleKong, Weizhen, Xiaolu Lv, Xiaotong Ran, Marguerite Mukangango, Bugenimana Eric Derrick, Baoli Qiu, and Changfei Guo. 2024. "Comprehensive Assessment of Reference Gene Expression within the Whitefly Dialeurodes citri Using RT-qPCR" Genes 15, no. 3: 318. https://doi.org/10.3390/genes15030318
APA StyleKong, W., Lv, X., Ran, X., Mukangango, M., Eric Derrick, B., Qiu, B., & Guo, C. (2024). Comprehensive Assessment of Reference Gene Expression within the Whitefly Dialeurodes citri Using RT-qPCR. Genes, 15(3), 318. https://doi.org/10.3390/genes15030318