
Shedding a Light on Dark Genes: A Comparative Expression Study of PRR12 Orthologues during Zebrafish Development

 and
and 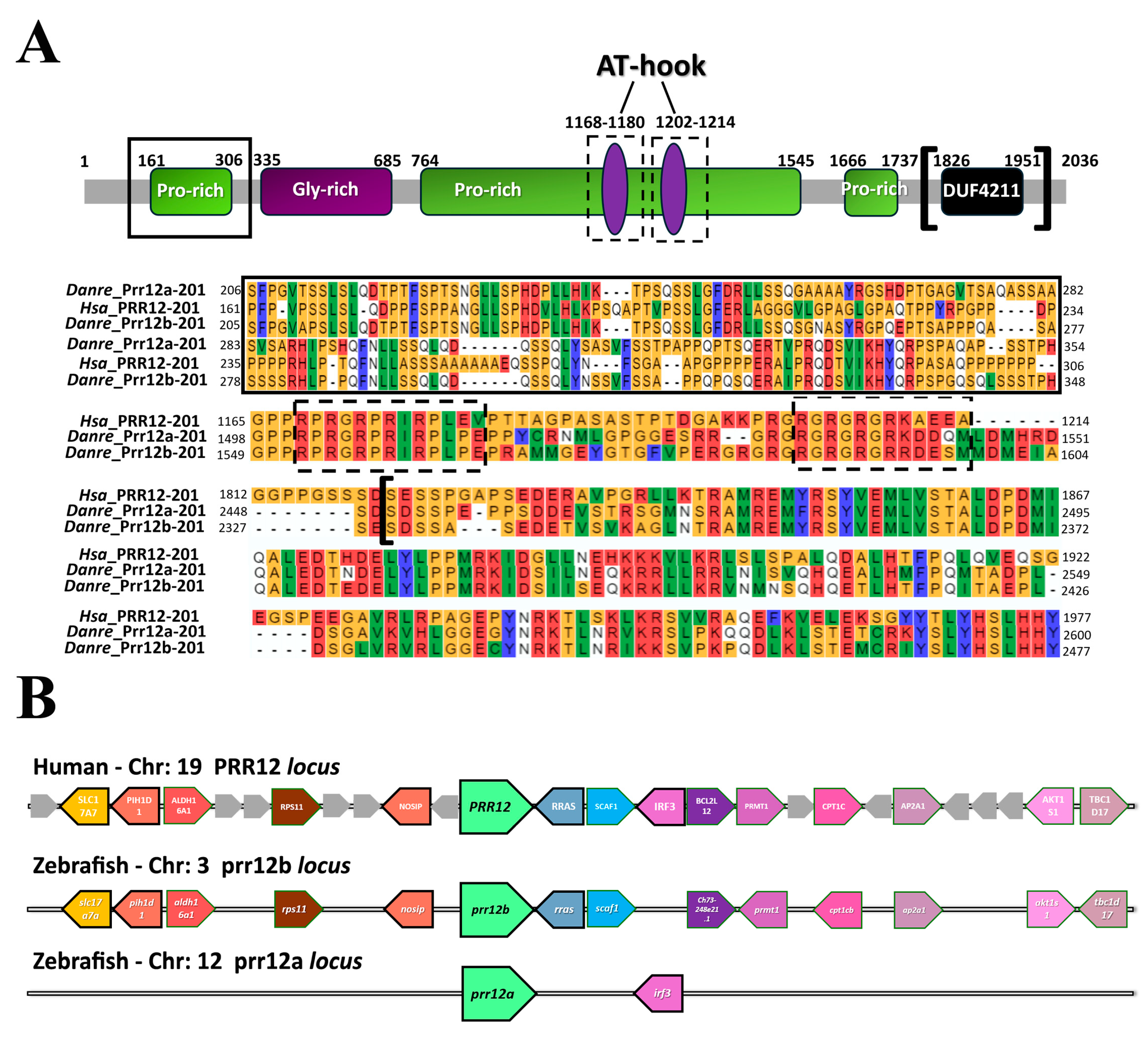{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Zebrafish Husbandry
2.2. Bioinformatic Analysis
| Gene Annotation | Transcript Annotation | |
| PRR12-201 | ENSG00000126464 | ENST00000418929.7 |
| PRR12-202 | ENSG00000126464 | ENST00000593853.1 |
| prr12a-201 | ZDB-GENE-041111-223; ENSDARG00000074229 | ENSDART00000110514.6 |
| prr12a-202 | ENSDARG00000074229 | ENSDART00000152396.2 |
| prr12b-201 | ZDB-GENE-130625-2; ENSDARG00000075849 | ENSDART00000156464.3 |
| prr12b-202 | ENSDARG00000075849 | ENSDART00000188447.1 |
2.3. Adult Brain and Eyes Dissection
2.4. RNA Extraction, RT-PCR and RT-qPCR
2.5. prr12a and prr12b Clones and Whole-Mount In Situ Hybridization (WISH)
2.6. ISH on Adult Brain Cryosections
2.7. Statistical Analysis
3. Results
3.1. In Silico Analysis of Zebrafish PRR12 Orthologs
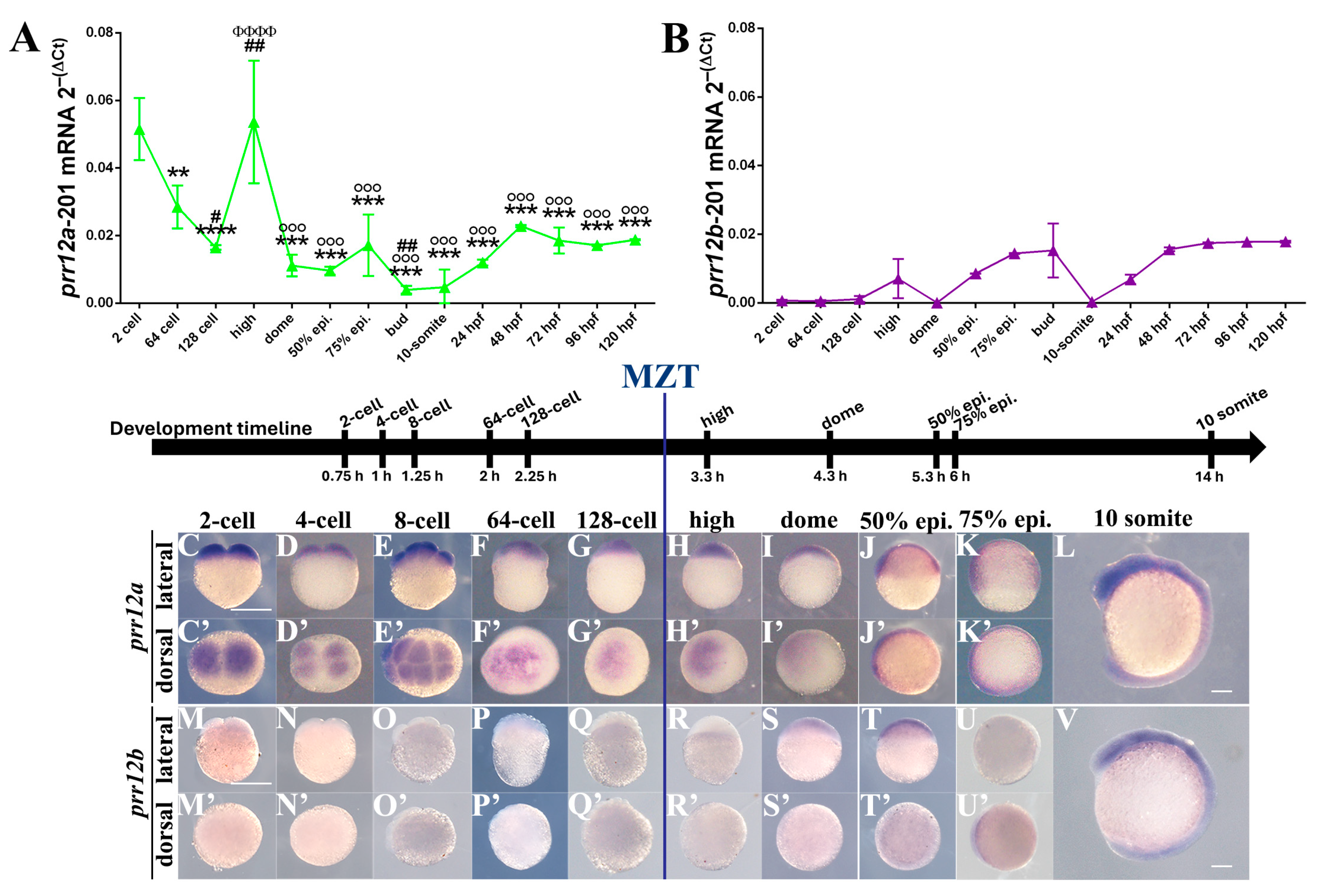
3.2. Temporal Expression: prr12a and prr12b Activation Is Differentially Regulated during Zebrafish Development
3.3. Spatial Expression: prr12a and prr12b Display Distinct Expression Patterns in the Developing and Adult Nervous System
3.3.1. Zygote (0.0 hpf)–MZT (3.00 hpf)
3.3.2. Mid-Blastula (3.33 hpf)–Segmentation (14 hpf)
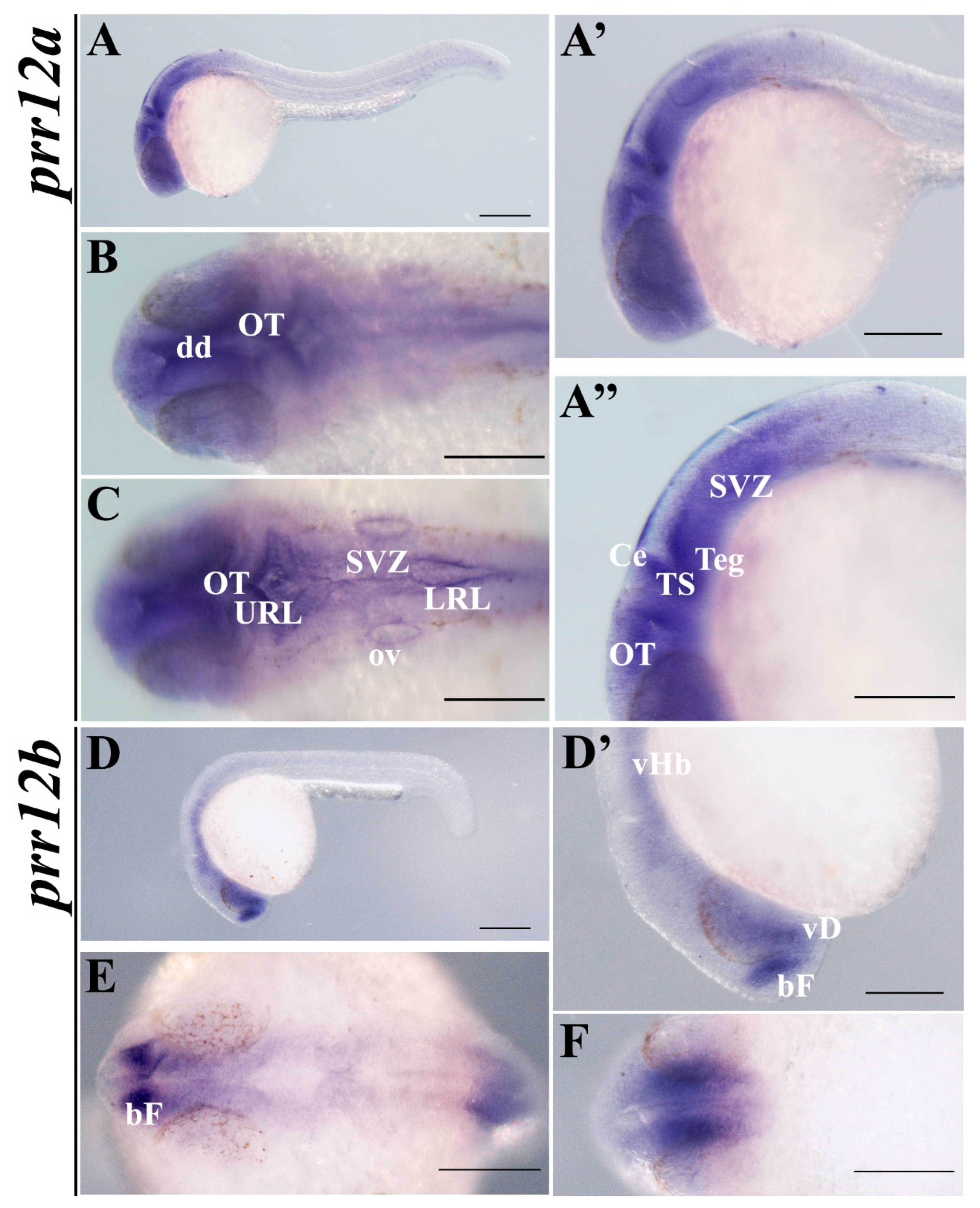
3.3.3. Pharyngula (24 hpf)
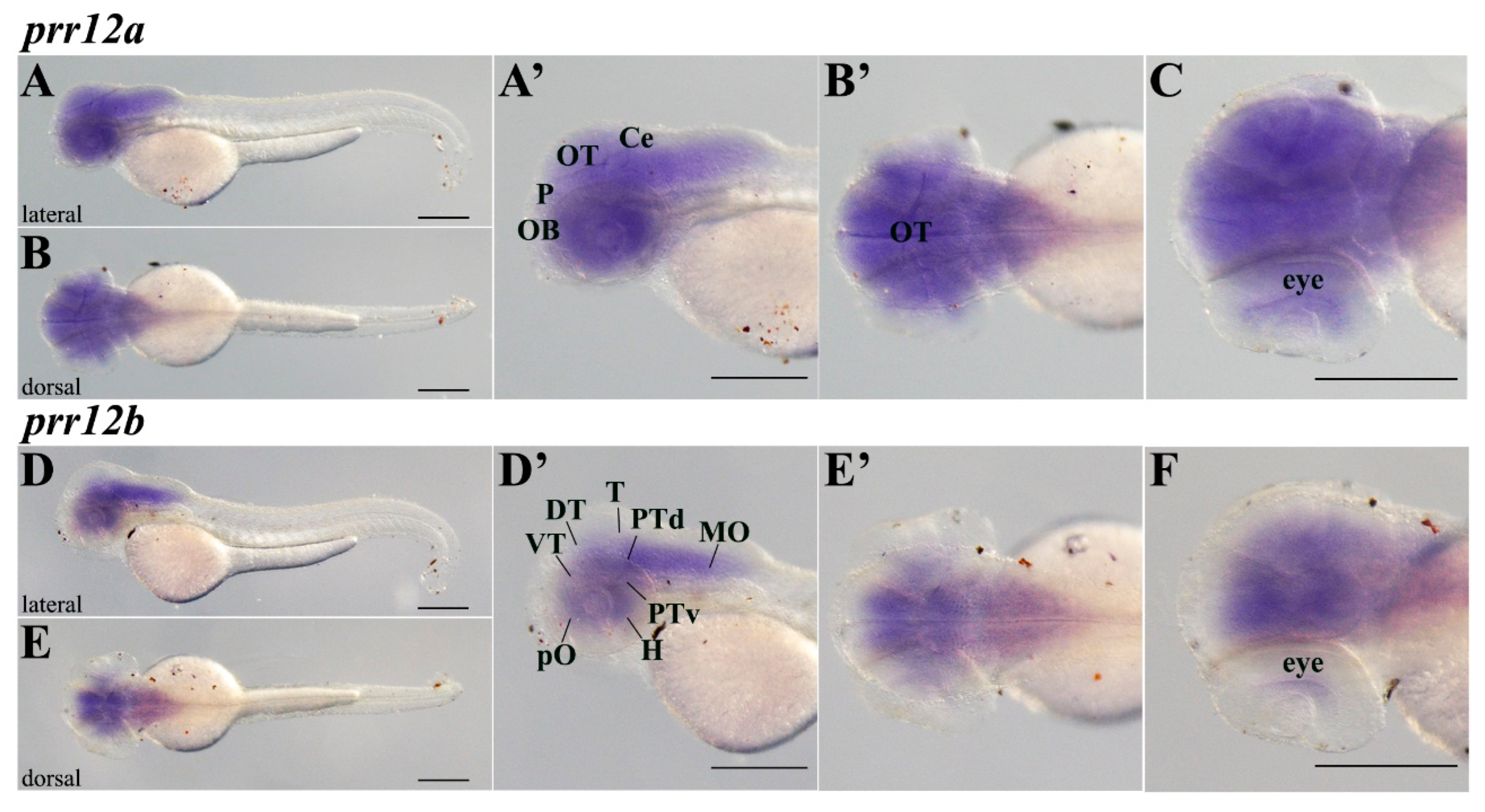
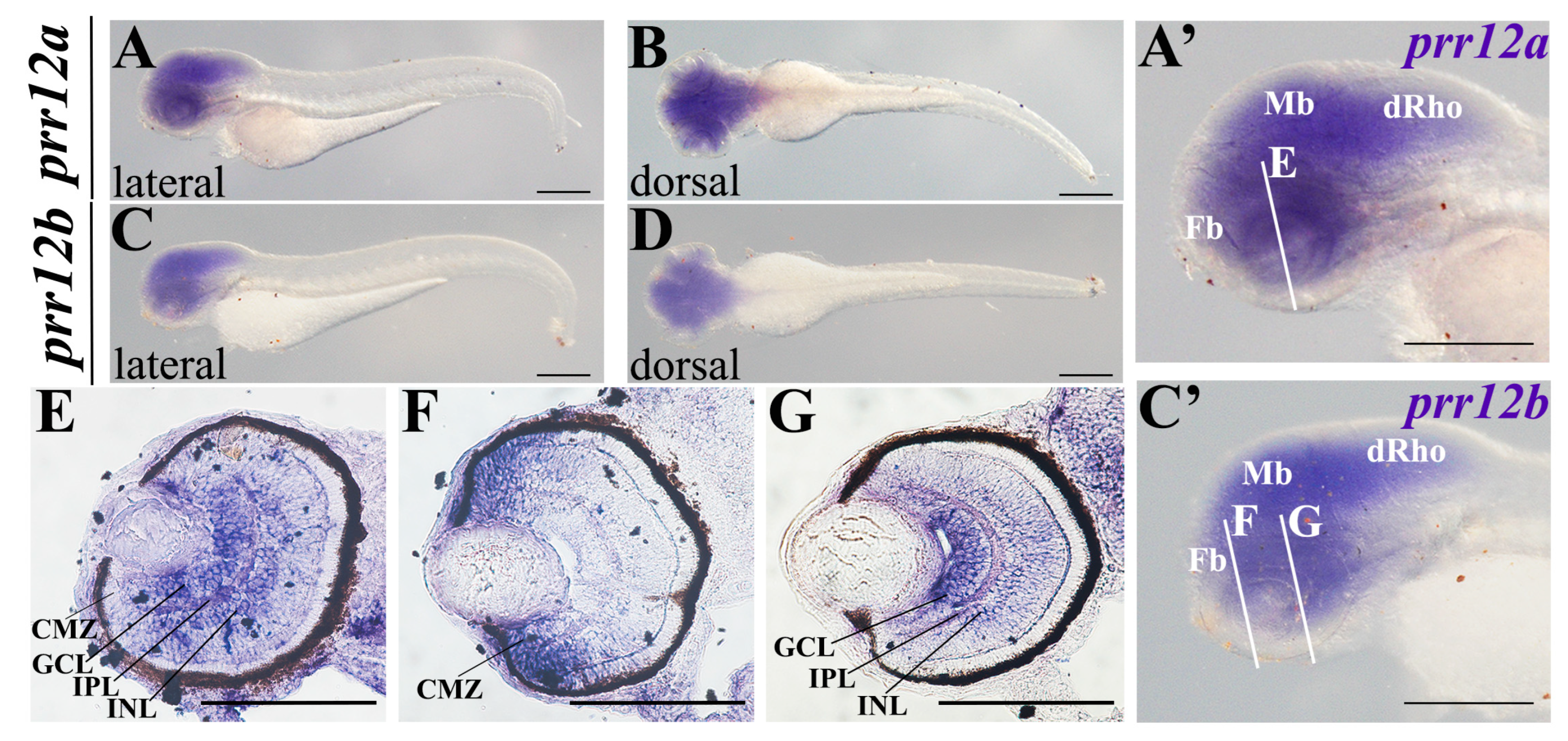
3.3.4. Hatching (48 hpf–72 hpf)
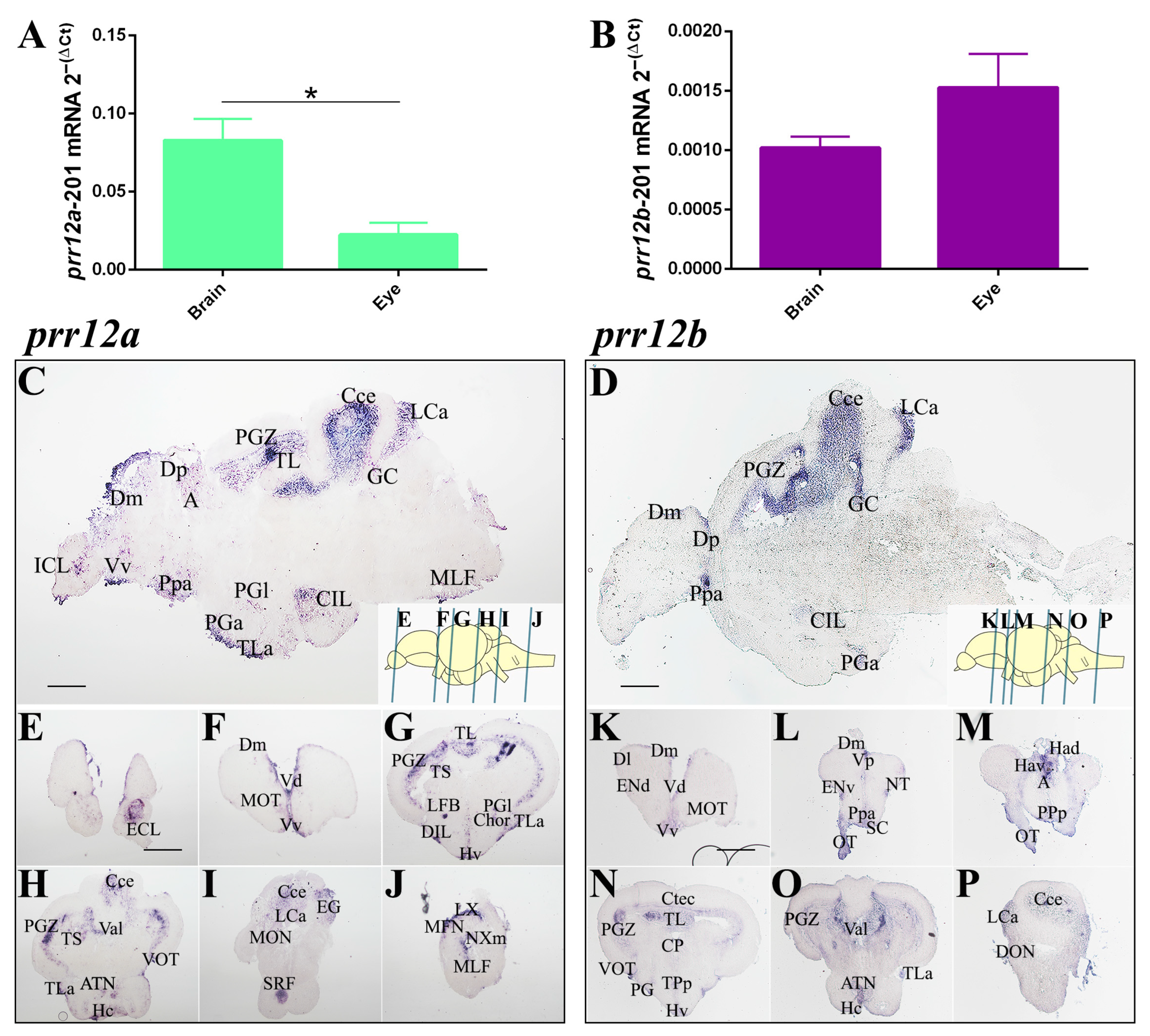
3.3.5. Adult Brain
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ebbert, M.T.W.; Jensen, T.D.; Jansen-West, K.; Sens, J.P.; Reddy, J.S.; Ridge, P.G.; Kauwe, J.S.K.; Belzil, V.; Pregent, L.; Carrasquillo, M.M.; et al. Systematic analysis of dark and camouflaged genes reveals disease-relevant genes hiding in plain sight. Genome Biol. 2019, 20, 97. [Google Scholar] [CrossRef]
- Acosta-Herrera, M.; Kerick, M.; Gonzalez-Serna, D.; Myositis Genetics, C.; Scleroderma Genetics, C.; Wijmenga, C.; Franke, A.; Gregersen, P.K.; Padyukov, L.; Worthington, J.; et al. Genome-wide meta-analysis reveals shared new loci in systemic seropositive rheumatic diseases. Ann. Rheum. Dis. 2019, 78, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Ryan, N.M.; Corvin, A. Investigating the dark-side of the genome: A barrier to human disease variant discovery? Biol. Res. 2023, 56, 42. [Google Scholar] [CrossRef]
- Nerenz, R.D.; Lefferts, J. Our Genome’s “Dark Matter” Is the Next Frontier in Molecular Diagnostics. Clin. Chem. 2017, 63, 792–793. [Google Scholar] [CrossRef] [PubMed]
- Oprea, T.I. Exploring the dark genome: Implications for precision medicine. Mamm. Genome 2019, 30, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Oprea, T.I.; Bologa, C.G.; Brunak, S.; Campbell, A.; Gan, G.N.; Gaulton, A.; Gomez, S.M.; Guha, R.; Hersey, A.; Holmes, J.; et al. Unexplored therapeutic opportunities in the human genome. Nat. Rev. Drug Discov. 2018, 17, 317–332. [Google Scholar] [CrossRef]
- Chowdhury, F.; Wang, L.; Al-Raqad, M.; Amor, D.J.; Baxova, A.; Bendova, S.; Biamino, E.; Brusco, A.; Caluseriu, O.; Cox, N.J.; et al. Haploinsufficiency of PRR12 causes a spectrum of neurodevelopmental, eye, and multisystem abnormalities. Genet. Med. 2021, 23, 1234–1245. [Google Scholar] [CrossRef]
- Cordova-Fletes, C.; Dominguez, M.G.; Delint-Ramirez, I.; Martinez-Rodriguez, H.G.; Rivas-Estilla, A.M.; Barros-Nunez, P.; Ortiz-Lopez, R.; Neira, V.A. A de novo t(10;19)(q22.3;q13.33) leads to ZMIZ1/PRR12 reciprocal fusion transcripts in a girl with intellectual disability and neuropsychiatric alterations. Neurogenetics 2015, 16, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Leduc, M.S.; McGuire, M.; Madan-Khetarpal, S.; Ortiz, D.; Hayflick, S.; Keller, K.; Eng, C.M.; Yang, Y.; Bi, W. De novo apparent loss-of-function mutations in PRR12 in three patients with intellectual disability and iris abnormalities. Hum. Genet. 2018, 137, 257–264. [Google Scholar] [CrossRef]
- Reis, L.M.; Costakos, D.; Wheeler, P.G.; Bardakjian, T.; Schneider, A.; Fung, S.S.M.; University of Washington Center for Mendelian, G.; Semina, E.V. Dominant variants in PRR12 result in unilateral or bilateral complex microphthalmia. Clin. Genet. 2021, 99, 437–442. [Google Scholar] [CrossRef]
- Mao, Y.; Shen, G.; Su, Z.; Du, J.; Xu, F.; Yu, Y. RAD21 inhibited transcription of tumor suppressor MIR4697HG and led to glioma tumorigenesis. Biomed. Pharmacother. 2020, 123, 109759. [Google Scholar] [CrossRef] [PubMed]
- Zarrinpar, A.; Bhattacharyya, R.P.; Lim, W.A. The structure and function of proline recognition domains. Sci. STKE 2003, 2003, RE8. [Google Scholar] [CrossRef] [PubMed]
- Rigbolt, K.T.; Prokhorova, T.A.; Akimov, V.; Henningsen, J.; Johansen, P.T.; Kratchmarova, I.; Kassem, M.; Mann, M.; Olsen, J.V.; Blagoev, B. System-wide temporal characterization of the proteome and phosphoproteome of human embryonic stem cell differentiation. Sci. Signal 2011, 4, rs3. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Bradford, Y.M.; Van Slyke, C.E.; Ruzicka, L.; Singer, A.; Eagle, A.; Fashena, D.; Howe, D.G.; Frazer, K.; Martin, R.; Paddock, H.; et al. Zebrafish information network, the knowledgebase for Danio rerio research. Genetics 2022, 220, iyac016. [Google Scholar] [CrossRef] [PubMed]
- Nikhil, B. Exon-Intron Graphic Maker. Available online: http://wormweb.org/exonintron (accessed on 15 March 2024).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef] [PubMed]
- UniProt, C. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Nguyen, N.T.T.; Vincens, P.; Roest Crollius, H.; Louis, A. Genomicus 2018: Karyotype evolutionary trees and on-the-fly synteny computing. Nucleic Acids Res. 2018, 46, D816–D822. [Google Scholar] [CrossRef]
- White, R.J.; Collins, J.E.; Sealy, I.M.; Wali, N.; Dooley, C.M.; Digby, Z.; Stemple, D.L.; Murphy, D.N.; Billis, K.; Hourlier, T.; et al. A high-resolution mRNA expression time course of embryonic development in zebrafish. eLife 2017, 6, e30860. [Google Scholar] [CrossRef]
- Moreno, P.; Fexova, S.; George, N.; Manning, J.R.; Miao, Z.; Mohammed, S.; Munoz-Pomer, A.; Fullgrabe, A.; Bi, Y.; Bush, N.; et al. Expression Atlas update: Gene and protein expression in multiple species. Nucleic Acids Res. 2022, 50, D129–D140. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Dente, L.; Gestri, G.; Tsang, M.; Kudoh, T.; Wilson, S.W.; Dawid, I.B.; Andreazzoli, M. Cloning and developmental expression of zebrafish pdzrn3. Int. J. Dev. Biol. 2011, 55, 989–993. [Google Scholar] [CrossRef] [PubMed]
- Gabellini, C.; Pucci, C.; De Cesari, C.; Martini, D.; Di Lauro, C.; Digregorio, M.; Norton, W.; Zippo, A.; Sessa, A.; Broccoli, V.; et al. CRISPR/Cas9-Induced Inactivation of the Autism-Risk Gene setd5 Leads to Social Impairments in Zebrafish. Int. J. Mol. Sci. 2022, 24, 167. [Google Scholar] [CrossRef]
- Ebstein, F.; Kury, S.; Papendorf, J.J.; Kruger, E. Neurodevelopmental Disorders (NDD) Caused by Genomic Alterations of the Ubiquitin-Proteasome System (UPS): The Possible Contribution of Immune Dysregulation to Disease Pathogenesis. Front. Mol. Neurosci. 2021, 14, 733012. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.B.; Willis, D.E.; Rodriguez, E.L.; Schwarz, J.M. Maternal immune activation as an epidemiological risk factor for neurodevelopmental disorders: Considerations of timing, severity, individual differences, and sex in human and rodent studies. Front. Neurosci. 2023, 17, 1135559. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Shi, Q.M.; Duan, S.L.; Ou-Yang, D.J.; Chen, P.; Tu, B.; Huang, P. Insights into the Association Between QSER1 and M2 Macrophages and Remarkable Malignancy Characteristics in Hepatocellular Carcinoma. Int. J. Gen. Med. 2022, 15, 1765–1775. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.; Strahle, U.; Scholpp, S. Neurogenesis in zebrafish—From embryo to adult. Neural Dev. 2013, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Ceriani, R.; Whitlock, K.E. Gonadotropin Releasing Hormone (Gnrh) Triggers Neurogenesis in the Hypothalamus of Adult Zebrafish. Int. J. Mol. Sci. 2021, 22, 5926. [Google Scholar] [CrossRef] [PubMed]
- Grandel, H.; Kaslin, J.; Ganz, J.; Wenzel, I.; Brand, M. Neural stem cells and neurogenesis in the adult zebrafish brain: Origin, proliferation dynamics, migration and cell fate. Dev. Biol. 2006, 295, 263–277. [Google Scholar] [CrossRef]
- Tesmer, A.L.; Fields, N.P.; Robles, E. Input from torus longitudinalis drives binocularity and spatial summation in zebrafish optic tectum. BMC Biol. 2022, 20, 24. [Google Scholar] [CrossRef] [PubMed]
- Porter, B.A.; Mueller, T. The Zebrafish Amygdaloid Complex—Functional Ground Plan, Molecular Delineation, and Everted Topology. Front. Neurosci. 2020, 14, 608. [Google Scholar] [CrossRef] [PubMed]
- Yanez, J.; Folgueira, M.; Lamas, I.; Anadon, R. The organization of the zebrafish pallium from a hodological perspective. J. Comp. Neurol. 2022, 530, 1164–1194. [Google Scholar] [CrossRef] [PubMed]
- Turner, K.J.; Hawkins, T.A.; Yanez, J.; Anadon, R.; Wilson, S.W.; Folgueira, M. Afferent Connectivity of the Zebrafish Habenulae. Front. Neural Circuits 2016, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Amo, R.; Fredes, F.; Kinoshita, M.; Aoki, R.; Aizawa, H.; Agetsuma, M.; Aoki, T.; Shiraki, T.; Kakinuma, H.; Matsuda, M.; et al. The habenulo-raphe serotonergic circuit encodes an aversive expectation value essential for adaptive active avoidance of danger. Neuron 2014, 84, 1034–1048. [Google Scholar] [CrossRef] [PubMed]
- Bloch, S.; Thomas, M.; Colin, I.; Galant, S.; Machado, E.; Affaticati, P.; Jenett, A.; Yamamoto, K. Mesencephalic origin of the inferior lobe in zebrafish. BMC Biol. 2019, 17, 22. [Google Scholar] [CrossRef] [PubMed]
- Mueller, T. What is the Thalamus in Zebrafish? Front. Neurosci. 2012, 6, 64. [Google Scholar] [CrossRef] [PubMed]
- Lek, M.; Karczewski, K.J.; Minikel, E.V.; Samocha, K.E.; Banks, E.; Fennell, T.; O’Donnell-Luria, A.H.; Ware, J.S.; Hill, A.J.; Cummings, B.B.; et al. Analysis of protein-coding genetic variation in 60,706 humans. Nature 2016, 536, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Flach, H.; Krieg, J.; Hoffmeister, M.; Dietmann, P.; Reusch, A.; Wischmann, L.; Kernl, B.; Riegger, R.; Oess, S.; Kuhl, S.J. Nosip functions during vertebrate eye and cranial cartilage development. Dev. Dyn. 2018, 247, 1070–1082. [Google Scholar] [CrossRef]
- Ohata, S.; Uga, H.; Okamoto, H.; Katada, T. Small GTPase R-Ras participates in neural tube formation in zebrafish embryonic spinal cord. Biochem. Biophys. Res. Commun. 2018, 501, 786–790. [Google Scholar] [CrossRef]
- Kim, D.I.; Park, M.J.; Choi, J.H.; Kim, I.S.; Han, H.J.; Yoon, K.C.; Park, S.W.; Lee, M.Y.; Oh, K.S.; Park, S.H. PRMT1 and PRMT4 Regulate Oxidative Stress-Induced Retinal Pigment Epithelial Cell Damage in SIRT1-Dependent and SIRT1-Independent Manners. Oxid. Med. Cell Longev. 2015, 2015, 617919. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Sheen, J.M.; Wang, S.C.; Hsu, M.H.; Hsiao, C.C.; Chang, K.A.; Huang, L.T. Methotrexate Neurotoxicity Is Related to Epigenetic Modification of the Myelination Process. Int. J. Mol. Sci. 2021, 22, 6718. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Fukamizu, A.; Nakagawa, T.; Kizuka, Y. Roles of protein arginine methyltransferase 1 (PRMT1) in brain development and disease. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129776. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Jeong, M.H.; Kim, K.R.; Jung, C.Y.; Lee, S.Y.; Kim, H.; Koh, J.; Vuong, T.A.; Jung, S.; Yang, H.; et al. Protein arginine methylation facilitates KCNQ channel-PIP2 interaction leading to seizure suppression. eLife 2016, 5, e17159. [Google Scholar] [CrossRef] [PubMed]
- Lauren, H.B.; Lopez-Picon, F.R.; Brandt, A.M.; Rios-Rojas, C.J.; Holopainen, I.E. Transcriptome analysis of the hippocampal CA1 pyramidal cell region after kainic acid-induced status epilepticus in juvenile rats. PLoS ONE 2010, 5, e10733. [Google Scholar] [CrossRef]
- Scammell, B.H.; Tchio, C.; Song, Y.; Nishiyama, T.; Louie, T.L.; Dashti, H.S.; Nakatochi, M.; Zee, P.C.; Daghlas, I.; Momozawa, Y.; et al. Multi-ancestry genome-wide analysis identifies shared genetic effects and common genetic variants for self-reported sleep duration. Hum. Mol. Genet. 2023, 32, 2797–2807. [Google Scholar] [CrossRef] [PubMed]
- Dixon, G.; Pan, H.; Yang, D.; Rosen, B.P.; Jashari, T.; Verma, N.; Pulecio, J.; Caspi, I.; Lee, K.; Stransky, S.; et al. QSER1 protects DNA methylation valleys from de novo methylation. Science 2021, 372, eabd0875. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, N.; Shimizu, K.; Sawamoto, K. Neuronal regeneration in a zebrafish model of adult brain injury. Dis. Model. Mech. 2012, 5, 200–209. [Google Scholar] [CrossRef]
- Kizil, C.; Kaslin, J.; Kroehne, V.; Brand, M. Adult neurogenesis and brain regeneration in zebrafish. Dev. Neurobiol. 2012, 72, 429–461. [Google Scholar] [CrossRef]
- Mueller, T.; Wullimann, M.F. An evolutionary interpretation of teleostean forebrain anatomy. Brain Behav. Evol. 2009, 74, 30–42. [Google Scholar] [CrossRef]
- Friedman, S.R.; Rapport, L.J.; Lumley, M.; Tzelepis, A.; VanVoorhis, A.; Stettner, L.; Kakaati, L. Aspects of social and emotional competence in adult attention-deficit/hyperactivity disorder. Neuropsychology 2003, 17, 50–58. [Google Scholar] [CrossRef]
- Frye, R.E. Social Skills Deficits in Autism Spectrum Disorder: Potential Biological Origins and Progress in Developing Therapeutic Agents. CNS Drugs 2018, 32, 713–734. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Peterson, R.T. The zebrafish subcortical social brain as a model for studying social behavior disorders. Dis. Model. Mech. 2019, 12, dmm039446. [Google Scholar] [CrossRef] [PubMed]
- Edden, R.A.; Crocetti, D.; Zhu, H.; Gilbert, D.L.; Mostofsky, S.H. Reduced GABA concentration in attention-deficit/hyperactivity disorder. Arch. Gen. Psychiatry 2012, 69, 750–753. [Google Scholar] [CrossRef]
- Zhao, H.; Mao, X.; Zhu, C.; Zou, X.; Peng, F.; Yang, W.; Li, B.; Li, G.; Ge, T.; Cui, R. GABAergic System Dysfunction in Autism Spectrum Disorders. Front. Cell Dev. Biol. 2021, 9, 781327. [Google Scholar] [CrossRef]
- Best, C.; Kurrasch, D.M.; Vijayan, M.M. Maternal cortisol stimulates neurogenesis and affects larval behaviour in zebrafish. Sci. Rep. 2017, 7, 40905. [Google Scholar] [CrossRef]
- Solek, C.M.; Feng, S.; Perin, S.; Weinschutz Mendes, H.; Ekker, M. Lineage tracing of dlx1a/2a and dlx5a/6a expressing cells in the developing zebrafish brain. Dev. Biol. 2017, 427, 131–147. [Google Scholar] [CrossRef]
- Rink, E.; Wullimann, M.F. Development of the catecholaminergic system in the early zebrafish brain: An immunohistochemical study. Brain Res. Dev. Brain Res. 2002, 137, 89–100. [Google Scholar] [CrossRef]
- Del Campo, N.; Chamberlain, S.R.; Sahakian, B.J.; Robbins, T.W. The roles of dopamine and noradrenaline in the pathophysiology and treatment of attention-deficit/hyperactivity disorder. Biol. Psychiatry 2011, 69, e145–e157. [Google Scholar] [CrossRef]
- Herborg, F.; Andreassen, T.F.; Berlin, F.; Loland, C.J.; Gether, U. Neuropsychiatric disease-associated genetic variants of the dopamine transporter display heterogeneous molecular phenotypes. J. Biol. Chem. 2018, 293, 7250–7262. [Google Scholar] [CrossRef]
- Bshary, R.; Gingins, S.; Vail, A.L. Social cognition in fishes. Trends Cogn. Sci. 2014, 18, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Sperber, S.M.; Saxena, V.; Hatch, G.; Ekker, M. Zebrafish dlx2a contributes to hindbrain neural crest survival, is necessary for differentiation of sensory ganglia and functions with dlx1a in maturation of the arch cartilage elements. Dev. Biol. 2008, 314, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Crow, A.J.D.; Janssen, J.M.; Vickers, K.L.; Parish-Morris, J.; Moberg, P.J.; Roalf, D.R. Olfactory Dysfunction in Neurodevelopmental Disorders: A Meta-analytic Review of Autism Spectrum Disorders, Attention Deficit/Hyperactivity Disorder and Obsessive-Compulsive Disorder. J. Autism Dev. Disord. 2020, 50, 2685–2697. [Google Scholar] [CrossRef] [PubMed]
- Romanos, M.; Renner, T.J.; Schecklmann, M.; Hummel, B.; Roos, M.; von Mering, C.; Pauli, P.; Reichmann, H.; Warnke, A.; Gerlach, M. Improved odor sensitivity in attention-deficit/hyperactivity disorder. Biol. Psychiatry 2008, 64, 938–940. [Google Scholar] [CrossRef] [PubMed]
- Karsz, F.R.; Vance, A.; Anderson, V.A.; Brann, P.G.; Wood, S.J.; Pantelis, C.; Brewer, W.J. Olfactory impairments in child attention-deficit/hyperactivity disorder. J. Clin. Psychiatry 2008, 69, 1462–1468. [Google Scholar] [CrossRef] [PubMed]
- Da Fonseca, D.; Seguier, V.; Santos, A.; Poinso, F.; Deruelle, C. Emotion understanding in children with ADHD. Child. Psychiatry Hum. Dev. 2009, 40, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Molnar-Szakacs, I.; Wang, M.J.; Laugeson, E.A.; Overy, K.; Wu, W.L.; Piggot, J. Autism, emotion recognition and the mirror neuron system: The case of music. Mcgill J. Med. 2009, 12, 87. [Google Scholar] [CrossRef]
- Rink, E.; Wullimann, M.F. Connections of the ventral telencephalon (subpallium) in the zebrafish (Danio rerio). Brain Res. 2004, 1011, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Herkenham, M.; Nauta, W.J. Afferent connections of the habenular nuclei in the rat. A horseradish peroxidase study, with a note on the fiber-of-passage problem. J. Comp. Neurol. 1977, 173, 123–146. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, S.; Parhar, I.S. Role of Habenula in Social and Reproductive Behaviors in Fish: Comparison with Mammals. Front. Behav. Neurosci. 2021, 15, 818782. [Google Scholar] [CrossRef]
- Gomez-Marin, C.; Tena, J.J.; Acemel, R.D.; Lopez-Mayorga, M.; Naranjo, S.; de la Calle-Mustienes, E.; Maeso, I.; Beccari, L.; Aneas, I.; Vielmas, E.; et al. Evolutionary comparison reveals that diverging CTCF sites are signatures of ancestral topological associating domains borders. Proc. Natl. Acad. Sci. USA 2015, 112, 7542–7547. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muscò, A.; Martini, D.; Digregorio, M.; Broccoli, V.; Andreazzoli, M. Shedding a Light on Dark Genes: A Comparative Expression Study of PRR12 Orthologues during Zebrafish Development. Genes 2024, 15, 492. https://doi.org/10.3390/genes15040492
Muscò A, Martini D, Digregorio M, Broccoli V, Andreazzoli M. Shedding a Light on Dark Genes: A Comparative Expression Study of PRR12 Orthologues during Zebrafish Development. Genes. 2024; 15(4):492. https://doi.org/10.3390/genes15040492
Chicago/Turabian StyleMuscò, Alessia, Davide Martini, Matteo Digregorio, Vania Broccoli, and Massimiliano Andreazzoli. 2024. "Shedding a Light on Dark Genes: A Comparative Expression Study of PRR12 Orthologues during Zebrafish Development" Genes 15, no. 4: 492. https://doi.org/10.3390/genes15040492
APA StyleMuscò, A., Martini, D., Digregorio, M., Broccoli, V., & Andreazzoli, M. (2024). Shedding a Light on Dark Genes: A Comparative Expression Study of PRR12 Orthologues during Zebrafish Development. Genes, 15(4), 492. https://doi.org/10.3390/genes15040492