Highlights
What are the main findings?
We introduce a SNP array based method for the genetic detection of Mongolian Gerbils.
What is the implication of the main finding?
Our approach enables the efficient and precise genetic analyses on outbred and inbred gerbil populations, which advance the level of genetic detection for characteristic gerbils in China.
Abstract
The Mongolian gerbil is a distinctive experimental animal in China, as its genetic qualities possess significant value in the field of medical biology research. Here, we aimed to establish an economical and efficient panel for genetic quality detection in Mongolian gerbils using single-nucleotide polymorphism (SNP) markers. To search for SNPs, we conducted whole-genome sequencing (WGS) in 40 Mongolian gerbils from outbred populations. Reliable screening criteria were established to preliminarily select SNPs with a wide genome distribution and high levels of polymorphism. Subsequently, a multiple-target regional capture detection system based on second-generation sequencing was developed for SNP genotyping. Based on the results of WGS, 219 SNPs were preliminarily selected, and they were established and optimized in a multiple-amplification system that included 206 SNP loci by genotyping three outbred populations. PopGen.32 analysis revealed that the average effective allele number, Shannon index, observed heterozygosity, expected heterozygosity, average heterozygosity, polymorphism information content, and other population genetic parameters of the Capital Medical University (CMU) gerbils were the highest, followed by those of Zhejiang gerbils and Dalian gerbils. Through scientific screening and optimization, we successfully established a novel, robust, and cost-effective genetic detection system for Mongolian gerbils by utilizing SNP markers for the first time.
1. Introduction
The Mongolian gerbil is a distinctive experimental animal in China, and it possesses significant value in the field of medical biology research. With the Mongolian gerbil, studies have been conducted on variations in the Circle of Willis (CoW) [1,2,3], cerebral ischemia [4,5,6], and gastric diseases associated with Helicobacter pylori [7,8]. Additionally, gerbil stroke models [9,10,11,12] and hereditary spontaneous diabetes models [13,14] have been established. Genetic monitoring is the main method employed to assess genetic diversity and the stability of the genetic background in experimental animals, enabling the detection of potential genetic mutations and genetic pollution [15]. Whole-genome sequencing (WGS) has already been conducted for the Mongolian gerbil [16], and the size of its whole genome length is about 3,931,855,312 bp [17]. Unfortunately, the available genetic testing methods for Mongolian gerbils are extremely limited. At present, the prevailing method of genetic analysis for gerbils in China is based on microsatellites [18,19,20,21], but an internationally acceptable testing approach for genetic quality remains absent.
In comparison with microsatellites, single-nucleotide polymorphisms (SNPs) possess the advantages of widespread distribution, enhanced stability, simplified analysis, and high throughput capacity, rendering them more suitable as markers for genetic monitoring. SNP markers have been widely used in genetic evaluations in many kinds of laboratory animals, such as mice [22,23,24,25] and rats [26,27,28,29]. They have been widely used in animal and plant breeding, disease-resistance gene markers, and the screening and identification of varieties of disease-related genes. The existing standard GB14923-2022 [30] “Genetic Quality Control of Experimental Animals” (China) recommends the utilization of 35 SNP marker genes for the genetic monitoring of inbred mice; however, there is currently a lack of well-defined SNP-based genetic quality assessment methods for Mongolian gerbils. Therefore, it is important to establish an internationally advanced genetic quality testing method using SNPs for Mongolian gerbils. This not only fills the gap in relevant standards but also holds significant implications for the enhancement of the genetic testing technology level and ensuring the overall quality of the Mongolian gerbil.
In this study, we succeeded in screening SNP loci with a wide distribution across the genome, high polymorphism rates, no linkage disequilibrium, and high confidence using large-scale whole-gene resequencing and multiple-target capture techniques. We developed an SNP genotyping method suitable for high-throughput genetic detection in Mongolian gerbils.
2. Materials and Methods
2.1. Animal Samples
Mongolian gerbils were collected from three outbred populations from Zhejiang (n = 37), CMU (n = 31), and Dalian (n = 32) and two inbred populations with diabetes (n = 10) and cerebral ischemia (n = 10). The gender of the gerbils was randomly selected. We randomly selected animals from the inbred populations and selected distantly genetically related individuals from the outbred populations. The original animals in the populations of Zhejiang, CMU, and Dalian were captured in inner Mongolia in 1978, 1982, and 1972, respectively. This study was approved by the Animal Experiments and Experimental Animal Welfare Committee of Capital Medical University (AEEI-2021-309).
2.2. DNA Extraction
A DNA extraction kit (FastPure® Cell/Tissue DNA Isolation Mini Kit, Vazyme Biotech Co., Ltd., Nanjing, China) was used to extract genomic DNA from the Mongolian gerbils’ ear and tail tissues. The concentration and purity of DNA from each sample were determined with an A260/A280 measurement using a Nanodrop 2000C micro-spectrophotometer (Thermo Fisher Scientific Inc., Waltham, MA USA) and further evaluated through agarose gel electrophoresis; then, the samples were stored at −20 °C.
2.3. Sequencing and Quality Control
Due to the limited availability of a comprehensive SNP database for the Mongolian gerbil, we conducted whole-genome sequencing (WGS) with a coverage of 10× on 20 Mongolian gerbils from Zhejiang and 20 CMU Mongolian gerbils, aiming to economically seek polymorphic SNP loci among animals through sequence comparison. Genomic DNA was sequenced with the Combinatorial Probe–Anchor Synthesis method in the DNBSEQ System (company, Beijing GenePlus Clinical Laboratory Co., Ltd., Beijing, China; machine, DNBSEQ-T7RS; read lengths, PE150; date out, 1.75-7T). Raw reads were filtered using the fastp-v0.21.0 software (https://github.com/OpenGene/fastp, accessed on 16 January 2023). Paired-end reads were mapped to MunDraft-v1.0 (https://www.ncbi.nlm.nih.gov/genome/gdv?org=meriones-unguiculatus&group=muridae, accessed on 16 January 2023) using the BWA-v0.7.17 software (https://sourceforge.net/projects/bio-bwa/files/, accessed on 16 January 2023). SNP calling and filtering were performed with the GATK-v4.0.0.0 software (https://gatk.broadinstitute.org, accessed on 16 January 2023). The BCFtools-v1.20-6 software (http://www.htslib.org/download/, accessed on 16 January 2023) was used for quality control of the data, and the following standards were set: (i) removal of SNP loci with a QualByDepth value of less than 2.0; (ii) removal of SNP loci with a FisherStrand value of less than 60.0; (iii) removal of SNP loci with an RMSMappingQuality value of less than 30.0; and (iv) removal of SNP loci with a sequencing depth of less than 8.0.
2.4. SNP Locus Selection
Our SNP screening criteria included the following five points: (i) for a uniform distribution across the genome, candidate loci were selected from different fragments to avoid linkage; (ii) the genotype frequency ranged between 25% and 75%, which was to ensure the polymorphism of the loci; (iii) to improve the amplification efficiency, no other SNPs or indels were found within a 200 bp region upstream or downstream of the selected loci; (iv) no complete linkage was observed among the selected loci; and, additionally, (v) so that they would correlate as much as possible with the phenotype, these SNPs predominantly resided in gene regions with a high concentration of missense mutations, nonsense mutations, and nonstop mutations. In accordance with this procedure, we screened a high-quality SNP locus panel.
2.5. Primer Design and Sequencing
After obtaining the candidate SNPs, we handed over the region of interest (ROI), that is, the information on the physical locations of the SNPs, to iGeneTech Biotechnology Beijing Co. Ltd., Beijing, China, and used MFEprimer-v3.1 to design and validate multiple PCR primers that targeted the genomic sequence of the SNPs in our panel. Based on the thermodynamic stability [31,32], highly specific multiplex primers were designed on both sides of the ROI; the amplicon was 160–260 bp. We then evaluated primer dimerization and non-specific amplification, tested the designed and synthesized primers, and replaced the primers that had a poor detection effect. Subsequently, sequencing was performed on an Illumina® NovaSeqTM 6000 system (Illumina, Inc., San Diego, CA USA) using amplicon-targeted capture in the PE150 paired-end sequencing mode.
2.6. Population Genetic Analysis
The genotype of each SNP locus in the form of AA, BB, and AB for all samples was input into the Popgen.32 Analysis software [33]. We used this software to calculate population genetic parameters such as average effective allele number, Shannon index, observed heterozygosity, expected heterozygosity, and average heterozygosity for different individuals at each SNP locus.
2.7. Compilation of Principal Component Analyses and Population Structure
Principal component analysis (PCA) and R (v4.3.2) were used to generate a PCA figure and a 3D PCA image. To estimate the genetic structure of our dataset, STRUCTURE v2.3.4 (https://web.stanford.edu/group/pritchardlab/structure.html, accessed on 16 January 2023) was run with 10,000 burn-ins and 50,000 iterations for the Markov Chain Monte Carlo (MCMC) method [34,35]. The frequency model of ‘correlated allele frequencies’ among populations was used, and an ‘admixture’ with K values ranging from 2 to 6 was tested across a total of ten runs. The estimated ln probability values (−LnP(D)) for each K value were plotted using the online STRUCTURE harvester tool v0.6.94 [35] (http://alumni.soe.ucsc.edu/~dearl/software/structureHarvester/, accessed on 16 January 2023).
3. Results
3.1. Establishment and Optimization of the SNP Detection System
By conducting a comparative analysis of the WGS results for 40 Mongolian gerbils, 3,853,611 SNP loci were identified in the genetic region. To screen an efficient SNP panel for genetic quality control in Mongolian gerbils, we established criteria for selecting high-quality SNPs from the SNP dataset derived from WGS (Figure 1). Based on the SNP screening criteria, we selected 219 high-quality loci for the development of the Mongolian gerbil SNP detection system (Supplementary Table S1).

Figure 1.
SNP screening criteria.
In order to further optimize the effective SNPs, we genotyped 219 SNP loci in 120 samples from five Mongolian gerbil populations. We deleted thirteen loci for the following reasons: (1) MAPQ < 59, indicating that the reads located at these loci were not unique compared with the reference genome and that the genotyping results were unreliable; (2) the loci were homomorphic in all samples; (3) there was complete linkage (Supplementary Table S2). In summary, a panel of 206 high-quality SNP loci was optimized and recommended for the genetic evaluation of Mongolian gerbil populations (Supplementary Table S3).
3.2. Genetic Analysis of the Outbred Mongolian Gerbil Populations
We applied the SNP detection system to the analysis of the genetic structure of three outbred Mongolian gerbil populations using the Popgen.32 software (Table 1). Our findings revealed that the Mongolian gerbils from CMU exhibited the highest values for various parameters, including the average effective allele numbers (ne), Shannon index (I), observed heterozygosity (Obs Het), expected heterozygosity (Exp Het), and average heterozygosity (Ave Het). The gerbils in the Zhejiang, diabetes, Dalian, and cerebral ischemia populations displayed progressively lower values.

Table 1.
Genetic structure of the three outbred Mongolian gerbil populations.
A dendrogram based on Nei’s genetic distance and the UPGMA method was created for the three outbred populations (CMU, Zhejiang, and Dalian), revealing that the Dalian Mongolian gerbils exhibited the greatest genetic divergence from the other Mongolian gerbil populations. Conversely, the Zhejiang population displayed a closer genetic distance to the CMU population (Figure 2).
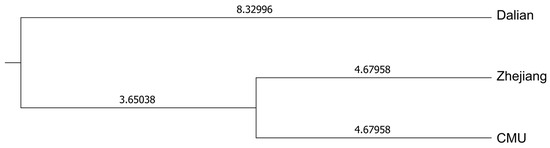
Figure 2.
Dendrogram based on Nei’s genetic distance and the UPGMA method. The numbers on each branch represent the length of each branch.
3.3. Structure Analysis of the Three Mongolian Gerbil Populations
The results of the population genetic structure analysis are shown in Figure 3. The number of assumed populations (K) was set to 2–6 in this study. It was found that the ΔK value was highest at K = 2, and it was recommended to divide this into two populations. The Dalian populations were separated from the other populations at K = 2. This was consistent with the results of the genetic distance analysis. Coherently, we found that the three populations showed different proportions of components at K = 3. These data indicate that the genetic structure of the outbred CMU population was similar to that of the outbred Zhejiang population.
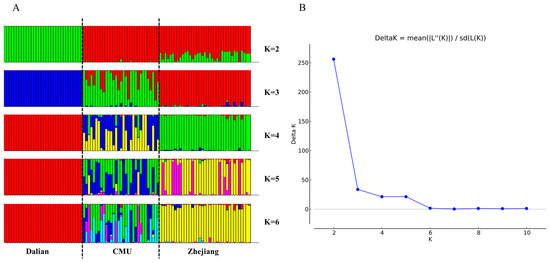
Figure 3.
Structure analysis of 206 SNPs in the three populations: (A) bar plot of K values ranging from 2 to 6; (B) the distribution of K values with ΔK.
3.4. Principal Component Analysis in the Three Mongolian Gerbil Populations
It is well established that PCA can be used to show the main differences in genetic distances between complex samples through a reduction in the data dimensionality. The results of PCA based on the allele frequency distributions of 206 polymorphic SNP loci in all three outbred populations are shown in Figure 4. In the genotype-based PCA results, PC1 (21.5%) and PC2 (8.2%) extracted 29.7% of the total genetic variation. The principal component analysis completely separated the Dalian population in a two-dimensional schematic diagram of PC1 (Figure 4A). All populations were separated by three PCs (Figure 4B). Again, the results of the PCA demonstrated that the Zhejiang and CMU populations were closer in terms of genetic distance.
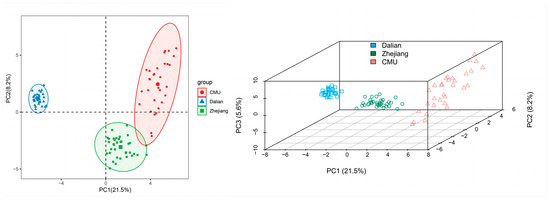
Figure 4.
The plot of the two-dimensional (A) and three-dimensional (B) principal component analyses among the three populations with 206 SNPs.
3.5. Genetic Analysis of the Outbred Mongolian Gerbil Populations
We also observed that the genotypic distribution of some loci was population-specific, and there were significant differences between populations. For instance, the locus MG15 exhibited polymorphism in the Dalian gerbils and Zhejiang gerbils with genotypes AG/GG/AA, but it exhibited monomorphism in the CMU gerbils, with only one kind of genotype (GG). The locus MG126 exhibited homomorphism in the Dalian and CMU gerbils, with only one genotype (CC or TT, respectively), whereas the Zhejiang gerbils exhibited three genotypes: TT/TC/CC. The three outbred populations could be easily distinguished with these two loci (Table 2). In total, we found seven loci that were specific in different populations (Table 2); they could be regarded as unique loci of a certain population and applied to easily distinguish the three outbred populations.
Table 2.
Allele frequencies of seven SNP loci in the three outbred populations.
3.6. Genetic Analysis of the Inbred Mongolian Gerbil Lines
In the panel of 206 SNP loci, 156 were found to be monomorphic in the inbred cerebral ischemia and 77 loci were found to be monomorphic in the inbred diabetes lines; 64 loci were monomorphic in both lines, but 15 of them exhibited different genotypes between the two lines and could be used to distinguish the two inbred populations (Table 3).
Table 3.
Fifteen loci with different phenotypes between the two inbred lines.
4. Discussion
This study presents the pioneering utilization of WGS to identify and select SNPs in Mongolian gerbils. By implementing stringent criteria, from a genetic region with 3,853,611 SNPs, we successfully identified 219 high-quality SNP loci with high levels of polymorphism and wide distribution across the genome and subsequently developed a multiple-SNP detection system.
In comparison with single-SNP amplification and genotyping methods, the multiple-target region capture technology (MultipSeq Primer for the genotyping of SNP loci) based on next-generation sequencing technology is more cost-effective and has higher efficiency, enabling the simultaneous acquisition of a greater number of loci and genotyping results [32,36,37]. In the present study, by testing the 219 selected SNPs in 120 animals, it was found that three loci had a low amplification quality, seven loci presented the same genotype in all animals, and three loci were completely linked with other loci. These loci were omitted due to their low capabilities for distinguishing different gerbil populations.
The use of reasonable criteria in the selection of high-quality SNPs that are suitable for genetic detection is a critically important issue. We noted that the loci were on separate chromosomes or sufficiently far apart on the same chromosome to ensure a wide distribution and show minimal linkage disequilibrium [38,39,40]. First, candidate loci were selected from different fragments across the entire genome. Second, no complete linkage was observed among the loci, thus improving the detection efficiency. To ensure that the loci were SNPs rather than SNVs, SNP sites were only conserved if MAF > 0.3 [40]. Third, the genotype frequency ranged between 25% and 75% in this study. To ensure the specific amplification of each locus [41], Wang et al. compared the reference genome of grapes and selected SNP loci with no other variations within the range of 100 bp before and after [40]. Fourth, no other SNPs or indels were found within a 200 bp region upstream or downstream of the selected loci in this study, thus further ensuring the specificity of locus amplification. SNPs were preferably located in coding regions [41], ensuring their close association with phenotypic changes. Fifth, in this study, the SNPs predominantly resided in gene regions with a high concentration of missense mutations, nonsense mutations, and nonstop mutations. These strict screening criteria were reasonable and creative and ensured the high reliability, polymorphism, and even distribution of all selected loci.
The average effective heterozygosity is an important parameter of population genetic diversity, effectively reflecting the richness of the identified genes [15,33]. When STR is employed as a marker for the detection of genetic diversity, it is generally believed that the population genetic diversity is low when the average effective heterozygosity is less than 0.5. When the average effective heterozygosity of a population is greater than 0.7, the genetic diversity is high [15]. However, the absolute values of the average effective heterozygosity of the three populations of gerbils in the present study were all lower than 0.5 [33,42]. This may be attributed to the inherent characteristics of different genetic markers; STR is always multi-allelic, and the number of alleles at a single locus can even reach higher than 10, while SNPs usually have only two alleles, indicating that we should apply different criteria to evaluate the population genetic diversity when using different markers [33]. Thus, our data indicated good genetic diversity among the three outbred populations. The results showed that the CMU population exhibited the highest genetic diversity; the Dalian group, on the other hand, exhibited the lowest values of genetic heterozygosity, which may have been related to the time at which the original animals were captured from the wild; the longer the duration of their confinement, the greater the reduction in their genetic diversity.
The loci exhibiting population-specific genotype distributions can serve as highly informative markers for population identification. At present, the existing standard GB14923-2022 [30] “Genetic Quality Control of Experimental Animals” (China) recommends the utilization of 35 SNP marker genes for the genetic monitoring of four inbred strains of mice. Wei et al. [43] also developed a combination of eight SNP loci that can distinguish 10 inbred mice strains. In the present study, seven SNP loci could be regarded as unique loci of a certain population and applied to easily distinguish the three outbred populations. The MG15 locus may be used to effectively differentiate the CMU population from the Dalian and Zhejiang populations, while a single locus, MG126, may be used to distinguish between the three populations of CMU, Zhejiang, and Dalian. A combination of fifteen SNP loci may be used to distinguish the two inbred populations.
Compared to the STR detection method, STR detection has advantages such as simple operation, easy interpretation, and low cost. The mutation rate of SNPs is only one-thousandth of STR, which is more stable during population inheritance. However, there are certain limitations of this study. The population cultivation of Mongolian gerbils is not as mature as that of mice. Only three populations were collected in the outbred population, and the number of animals selected for sequencing validation was small, resulting in a scarce amount of data. Some differential loci need to be further determined by expanding the sample size of the population in later stages of research.
5. Conclusions
We successfully established a novel genetic detection system for SNPs in Mongolian gerbils that is efficient and precise when conducting genetic analyses on several outbred and inbred gerbil populations. These findings will significantly advance the level of genetic detection for characteristic gerbils in China.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/genes15060817/s1: Table S1: Panel with 219 SNPs from Mongolian gerbils; Table S2: The thirteen SNP loci that were deleted; Table S3: Panel with 206 SNPs from Mongolian gerbils; Table S4: Mongolian gerbil primers; Table S5: PCA software codes for analysis.
Author Contributions
Conceptualization, X.H., Z.C., J.L. (Jing Lu) and X.D.; methodology, Y.G., M.S., Y.C., X.Z., C.L. and Y.Z.; software, Y.G., M.S., X.Z., J.L. (Jianyi Lv) and Y.Z.; validation, Y.G.; formal analysis, Y.G.; investigation, Y.G., M.S., X.Z., Y.C. and M.G.; resources, X.H., Z.C. and X.D.; data curation, Y.G., M.S., X.Z., X.L. and Y.C.; writing—original draft preparation, Y.G. and Y.C.; writing—review and editing, Y.G., Y.C., X.H., Z.C. and X.D.; visualization, Y.G. and M.S.; supervision, X.H., Z.C. and X.D.; project administration, X.H., Z.C. and X.D.; funding acquisition, X.H., Z.C. and X.D. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the National Key Research and Development Program for Young Scientists (no. 2021YFF0703200), the National Key Research and Development Program (no. 2022YFF0711005), and the National Natural Science Foundation of China (no. 32170540).
Institutional Review Board Statement
This study was approved by the Animal Experiments and Experimental Animal Welfare Committee of Capital Medical University (AEEI-2021-309).
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
We cordially thank Jingyu Wang of Dalian Medical University Laboratory Animal Center and Huazhong Ying of Hangzhou Medical College Laboratory Animal Center for sharing Mongolian gerbils.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Abe, Y.; Toyama, K.; Shinohara, A.; Nagura-Kato, G.A.; Ikai, Y.; Koshimoto, C.; Spin, J.M.; Hato, N. Message to researchers: The characteristic absence of a posterior communicating artery is easily lost in the gerbil. Anat. Sci. Int. 2023, 98, 426–433. [Google Scholar] [CrossRef]
- Du, X.; Wang, D.; Li, Y.; Huo, X.; Li, C.; Lu, J.; Wang, Y.; Guo, M.; Chen, Z. Newly breeding an inbred strain of ischemia-prone Mongolian gerbils and its reproduction and genetic characteristics. Exp. Anim. 2018, 67, 83–90. [Google Scholar] [CrossRef]
- Kondo, T.; Yoshida, S.; Nagai, H.; Takeshita, A.; Mino, M.; Morioka, H.; Nakajima, T.; Kusakabe, K.T.; Okada, T. Transient forebrain ischemia induces impairment in cognitive performance prior to extensive neuronal cell death in Mongolian gerbil (Meriones unguiculatus). J. Vet. Sci. 2018, 19, 505–511. [Google Scholar] [CrossRef]
- Selvaraj, B.; Le, T.T.; Kim, D.W.; Jung, B.H.; Yoo, K.Y.; Ahn, H.R.; Thuong, P.T.; Tran, T.T.T.; Pae, A.N.; Jung, S.H.; et al. Neuroprotective Effects of Ethanol Extract of Polyscias fruticosa (EEPF) against Glutamate-Mediated Neuronal Toxicity in HT22 Cells. Int. J. Mol. Sci. 2023, 24, 3969. [Google Scholar] [CrossRef]
- Shen, Y.; Lu, H.; Xu, R.; Tian, H.; Xia, X.; Zhou, F.H.; Wang, L.; Dong, J.; Sun, L. The Expression of GLAST and GLT1 in a Transient Cerebral Ischemia Mongolian Gerbil Model. Neuropsychiatr. Dis. Treat. 2020, 16, 789–800. [Google Scholar] [CrossRef]
- Ye, Q.; Hai, K.; Liu, W.; Wang, Y.; Zhou, X.; Ye, Z.; Liu, X. Investigation of the protective effect of heparin pre-treatment on cerebral ischaemia in gerbils. Pharm. Biol. 2019, 57, 519–528. [Google Scholar] [CrossRef]
- Noto, J.M.; Rose, K.L.; Hachey, A.J.; Delgado, A.G.; Romero-Gallo, J.; Wroblewski, L.E.; Schneider, B.G.; Shah, S.C.; Cover, T.L.; Wilson, K.T.; et al. Carcinogenic Helicobacter pylori Strains Selectively Dysregulate the In Vivo Gastric Proteome, Which May Be Associated with Stomach Cancer Progression. Mol. Cell Proteom. 2019, 18, 352–371. [Google Scholar] [CrossRef]
- Amalia, R.; Panenggak, N.S.R.; Doohan, D.; Rezkitha, Y.A.A.; Waskito, L.A.; Syam, A.F.; Lubis, M.; Yamaoka, Y.; Miftahussurur, M. A comprehensive evaluation of an animal model for Helicobacter pylori-associated stomach cancer: Fact and controversy. Helicobacter 2023, 28, e12943. [Google Scholar] [CrossRef]
- Lee, T.K.; Hong, J.; Lee, J.W.; Kim, S.S.; Sim, H.; Lee, J.C.; Kim, D.W.; Lim, S.S.; Kang, I.J.; Won, M.H. Ischemia-Induced Cognitive Impairment Is Improved via Remyelination and Restoration of Synaptic Density in the Hippocampus after Treatment with COG-Up(®) in a Gerbil Model of Ischemic Stroke. Vet. Sci. 2021, 8, 321. [Google Scholar] [CrossRef]
- Zhou, L.; Jia, R.; Zeng, W.; Cai, Q.; Qu, Y. Study on the Difference of Protective Efficacy and Mechanism of Radix Aconiti Coreani and Rhizoma Typhonii in Gerbils with Ischemic Stroke. ACS Chem. Neurosci. 2023, 14, 3686–3693. [Google Scholar] [CrossRef]
- Jeong, D.Y.; Jeong, S.Y.; Zhang, T.; Wu, X.; Qiu, J.Y.; Park, S. Chungkookjang, a soy food, fermented with Bacillus amyloliquefaciens protects gerbils against ishcmeic stroke injury, and post-stroke hyperglycemia. Food Res. Int. 2020, 128, 108769. [Google Scholar] [CrossRef]
- Kim, B.; Lee, T.K.; Park, C.W.; Kim, D.W.; Ahn, J.H.; Sim, H.; Lee, J.C.; Yang, G.E.; Kim, J.D.; Shin, M.C.; et al. Pycnogenol(®) Supplementation Attenuates Memory Deficits and Protects Hippocampal CA1 Pyramidal Neurons via Antioxidative Role in a Gerbil Model of Transient Forebrain Ischemia. Nutrients 2020, 12, 2477. [Google Scholar] [CrossRef]
- Li, X.; Lu, J.; Wang, Y.; Huo, X.; Li, Z.; Zhang, S.; Li, C.; Guo, M.; Du, X.; Chen, Z. Establishment and Characterization of a Newly Established Diabetic Gerbil Line. PLoS ONE 2016, 11, e0159420. [Google Scholar] [CrossRef]
- Gong, J.; Du, X.; Li, Z.; Li, X.; Guo, M.; Lu, J.; Wang, Y.; Chen, Z.; Li, C. Differential expression of genes identified by suppression subtractive hybridization in liver and adipose tissue of gerbils with diabetes. PLoS ONE 2018, 13, e0191212. [Google Scholar] [CrossRef]
- Zhang, X.; He, Y.; Zhang, W.; Wang, Y.; Liu, X.; Cui, A.; Gong, Y.; Lu, J.; Liu, X.; Huo, X.; et al. Development of Microsatellite Marker System to Determine the Genetic Diversity of Experimental Chicken, Duck, Goose, and Pigeon Populations. Biomed. Res. Int. 2021, 2021, 8851888. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, P.; Song, Z.; Du, X.; Huo, X.; Lu, J.; Liu, X.; Lv, J.; Li, C.; Guo, M.; et al. Generation of Gene-Knockout Mongolian Gerbils via CRISPR/Cas9 System. Front. Bioeng. Biotechnol. 2020, 8, 780. [Google Scholar] [CrossRef]
- Li, J. Full-Length Transcriptome Analysis of Mongolian Gerbils. 2023. Available online: https://link.cnki.net/doi/10.27229/d.cnki.gnmnu.2023.001321 (accessed on 15 June 2024).
- Liu, X.; Yu, X.; Xu, Y.; Du, X.; Huo, X.; Li, C.; Lv, J.; Guo, M.; Lu, J.; Chen, Z. Development of an effective microsatellite marker system to determine the genetic structure of Meriones meridianus populations. Exp. Anim. 2020, 69, 224–232. [Google Scholar] [CrossRef]
- Deng, K.; Liu, W.; Wang, D.H. Relatedness and spatial distance modulate intergroup interactions: Experimental evidence from a social rodent. Curr. Zool. 2019, 65, 527–534. [Google Scholar] [CrossRef]
- Du, X.Y.; Li, W.; Sa, X.Y.; Li, C.L.; Lu, J.; Wang, Y.Z.; Chen, Z.W. Selection of an effective microsatellite marker system for genetic control and analysis of gerbil populations in China. Genet. Mol. Res. 2015, 14, 11030–11042. [Google Scholar] [CrossRef]
- Wang, G.; Liu, W.; Wang, Y.; Wan, X.; Zhong, W. Restricted dispersal determines fine-scale spatial genetic structure of Mongolian gerbils. Curr. Zool. 2017, 63, 687–691. [Google Scholar] [CrossRef]
- Mekada, K.; Yoshiki, A. Substrains matter in phenotyping of C57BL/6 mice. Exp. Anim. 2021, 70, 145–160. [Google Scholar] [CrossRef]
- Liu, Y.; Chudgar, N.; Mastrogiacomo, B.; He, D.; Lankadasari, M.B.; Bapat, S.; Jones, G.D.; Sanchez-Vega, F.; Tan, K.S.; Schultz, N.; et al. A germline SNP in BRMS1 predisposes patients with lung adenocarcinoma to metastasis and can be ameliorated by targeting c-fos. Sci. Transl. Med. 2022, 14, eabo1050. [Google Scholar] [CrossRef]
- Brynildsen, J.K.; Yang, K.; Lemchi, C.; Dani, J.A.; De Biasi, M.; Blendy, J.A. A common SNP in Chrna5 enhances morphine reward in female mice. Neuropharmacology 2022, 218, 109218. [Google Scholar] [CrossRef]
- Chen, C.L.; Rodiger, J.; Chung, V.; Viswanatha, R.; Mohr, S.E.; Hu, Y.; Perrimon, N. SNP-CRISPR: A Web Tool for SNP-Specific Genome Editing. G3 2020, 10, 489–494. [Google Scholar] [CrossRef]
- Wang, X.; Lu, C.; Chen, Y.; Wang, Q.; Bao, X.; Zhang, Z.; Huang, X. Resveratrol promotes bone mass in ovariectomized rats and the SIRT1 rs7896005 SNP is associated with bone mass in women during perimenopause and early postmenopause. Climacteric 2023, 26, 25–33. [Google Scholar] [CrossRef]
- Fowler, S.; Wang, T.; Munro, D.; Kumar, A.; Chitre, A.S.; Hollingsworth, T.J.; Garcia Martinez, A.; St Pierre, C.L.; Bimschleger, H.; Gao, J.; et al. Genome-wide association study finds multiple loci associated with intraocular pressure in HS rats. Front. Genet. 2022, 13, 1029058. [Google Scholar] [CrossRef]
- Devyatkin, V.A.; Redina, O.E.; Kolosova, N.G.; Muraleva, N.A. Single-Nucleotide Polymorphisms Associated with the Senescence-Accelerated Phenotype of OXYS Rats: A Focus on Alzheimer’s Disease-Like and Age-Related-Macular-Degeneration-Like Pathologies. J. Alzheimers Dis. 2020, 73, 1167–1183. [Google Scholar] [CrossRef]
- Kitagawa, A.; Kizub, I.; Jacob, C.; Michael, K.; D’Alessandro, A.; Reisz, J.A.; Grzybowski, M.; Geurts, A.M.; Rocic, P.; Gupte, R.; et al. CRISPR-Mediated Single Nucleotide Polymorphism Modeling in Rats Reveals Insight Into Reduced Cardiovascular Risk Associated With Mediterranean G6PD Variant. Hypertension 2020, 76, 523–532. [Google Scholar] [CrossRef]
- GB 14923-2022; Laboratory Animal—Genetic Quality Control. CHINESE GB Standards: Beijing, China, 2022. Available online: http://c.gb688.cn/bzgk/gb/showGb?type=download&hcno=D5AD6845D0955F91D1F53D3FE8DE2FDB (accessed on 15 June 2024).
- Wang, K.; Li, H.; Xu, Y.; Shao, Q.; Yi, J.; Wang, R.; Cai, W.; Hang, X.; Zhang, C.; Cai, H.; et al. MFEprimer-3.0: Quality control for PCR primers. Nucleic Acids Res. 2019, 47, W610–W613. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Xue, J.; Tan, M.; Chen, D.; Xiao, Y.; Liu, G.; Zheng, Y.; Wu, Q.; Liao, M.; Lv, M.; et al. An MPS-Based 50plex Microhaplotype Assay for Forensic DNA Analysis. Genes 2023, 14, 865. [Google Scholar] [CrossRef] [PubMed]
- LI, Y.; Wang, H.; Li, C.; Guo, M.; Wei, J.; Zhang, Y.; Huang, R.; Chen, Z.; Du, X. Population Genetic Structure Analysis of 3 Outbred Mice Populations from National Rodent Seed Center Using 45 loci of Single Nucleotide Polymorphism. Lab. Anim. Sci. 2018, 35, 1–7. [Google Scholar]
- Porras-Hurtado, L.; Ruiz, Y.; Santos, C.; Phillips, C.; Carracedo, A.; Lareu, M.V. An overview of STRUCTURE: Applications, parameter settings, and supporting software. Front. Genet. 2013, 4, 98. [Google Scholar] [CrossRef]
- Oldoni, F.; Yoon, L.; Wootton, S.C.; Lagacé, R.; Kidd, K.K.; Podini, D. Population genetic data of 74 microhaplotypes in four major U.S. population groups. Forensic Sci. Int. Genet. 2020, 49, 102398. [Google Scholar] [CrossRef]
- Wang, X.; Shi, W.; Zhao, S.; Gong, D.; Li, S.; Hu, C.; Chen, Z.J.; Li, Y.; Yan, J. Whole exome sequencing in unexplained recurrent miscarriage families identified novel pathogenic genetic causes of euploid miscarriage. Hum. Reprod. 2023, 38, 1003–1018. [Google Scholar] [CrossRef]
- Zhang, Q.; Qi, Y.; Wang, S.; Zhao, F.; Zou, L.; Zhou, Q.; Geng, P.; Hong, Y.; Yang, H.; Luo, Q.; et al. Identification and in vitro functional assessment of 10 CYP2C9 variants found in Chinese Han subjects. Front. Endocrinol. 2023, 14, 1139805. [Google Scholar] [CrossRef]
- Kidd, K.K.; Pakstis, A.J.; Speed, W.C.; Lagacé, R.; Chang, J.; Wootton, S.; Haigh, E.; Kidd, J.R. Current sequencing technology makes microhaplotypes a powerful new type of genetic marker for forensics. Forensic Sci. Int. Genet. 2014, 12, 215–224. [Google Scholar] [CrossRef]
- Andrews, K.R.; Hunter, S.S.; Torrevillas, B.K.; Céspedes, N.; Garrison, S.M.; Strickland, J.; Wagers, D.; Hansten, G.; New, D.D.; Fagnan, M.W.; et al. A new mouse SNP genotyping assay for speed congenics: Combining flexibility, affordability, and power. BMC Genom. 2021, 22, 378. [Google Scholar] [CrossRef]
- Wang, F.-Q.; Fan, X.-C.; Zhang, Y.; Sun, L.; Liu, C.-H.; Jiang, J.-F. Establishment and application of an SNP molecular identification system for grape cultivars. J. Integr. Agric. 2022, 21, 1044–1057. [Google Scholar] [CrossRef]
- Petkov, P.M.; Ding, Y.; Cassell, M.A.; Zhang, W.; Wagner, G.; Sargent, E.E.; Asquith, S.; Crew, V.; Johnson, K.A.; Robinson, P.; et al. An efficient SNP system for mouse genome scanning and elucidating strain relationships. Genome Res. 2004, 14, 1806–1811. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Z.; Liu, J.; Li, C.; Liu, X.; Lv, J.; Guo, M.; Du, X. Genetic Mornitoring for Inbred Mice with Single Nucleotide Polymorphisms. Lab. Anim. Comp. Med. 2018, 38, 16–21. [Google Scholar]
- Wei, J.; Wang, H.; Zhou, J.; Zhao, L.; Li, H.; Yue, B. Application of SNP Markers in Identification of Inbred Mice Strains and Primer Sequences. CN115976226A, 18 April 2023. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).