
Protein Assemblies in Translesion Synthesis

Department of Molecular Biology and Biophysics, University of Connecticut Health Center, Farmington, CT 06030, USA
*
Author to whom correspondence should be addressed.
Genes 2024, 15(7), 832; https://doi.org/10.3390/genes15070832
Submission received: 29 May 2024
/
Revised: 19 June 2024
/
Accepted: 20 June 2024
/
Published: 24 June 2024
(This article belongs to the Special Issue DNA Damage and Repair in Microorganisms, Plants and Mammalian Systems)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Translesion synthesis (TLS) is a mechanism of DNA damage tolerance utilized by eukaryotic cells to replicate DNA across lesions that impede the high-fidelity replication machinery. In TLS, a series of specialized DNA polymerases are employed, which recognize specific DNA lesions, insert nucleotides across the damage, and extend the distorted primer-template. This allows cells to preserve genetic integrity at the cost of mutations. In humans, TLS enzymes include the Y-family, inserter polymerases, Polη, Polι, Polκ, Rev1, and the B-family extender polymerase Polζ, while in S. cerevisiae only Polη, Rev1, and Polζ are present. To bypass DNA lesions, TLS polymerases cooperate, assembling into a complex on the eukaryotic sliding clamp, PCNA, termed the TLS mutasome. The mutasome assembly is contingent on protein–protein interactions (PPIs) between the modular domains and subunits of TLS enzymes, and their interactions with PCNA and DNA. While the structural mechanisms of DNA lesion bypass by the TLS polymerases and PPIs of their individual modules are well understood, the mechanisms by which they cooperate in the context of TLS complexes have remained elusive. This review focuses on structural studies of TLS polymerases and describes the case of TLS holoenzyme assemblies in action emerging from recent high-resolution Cryo-EM studies.
1. Introduction
Eukaryotic DNA is continuously altered by endogenous and exogenous genotoxic agents [1,2,3,4,5]. The resulting DNA lesions are removed by DNA repair mechanisms such as base excision repair (BER) [6] or nucleotide excision repair (NER) [7]; however, some damage persists into S-phase, impeding DNA replication. The high-fidelity replicative DNA polymerases, Polδ and Polε, can stall, unable to accommodate structurally aberrant DNA in their restrictive active sites. Replication fork stalling can lead to fork collapse and the build-up of single-stranded (ss) DNA prone to deleterious damage such as double stranded breaks (DSBs) [8,9,10,11]. To protect genomic integrity, cells have evolved DNA damage tolerance (DDT) pathways that enable the replication of damaged DNA without repairing the damage [12,13,14]. The error-prone branch of DDT called translesion synthesis (TLS) employs specialized, low-fidelity DNA polymerases that can take over replication and insert nucleotides across DNA lesions or fill lesion-containing ssDNA gaps left after replication during G2/M-phase [15,16,17,18,19,20,21,22,23,24]. While critical for DNA replication in healthy cells, TLS DNA polymerases are upregulated in many cancers and contribute to cancer resistance to DNA damage induced by genotoxic therapies such as cisplatin [25,26,27,28,29,30,31]. As such, TLS inhibition has emerged as a strategy to enhance the efficacy of first-line chemotherapy in cancer treatment [32,33,34,35,36], highlighting the importance of understanding the structural mechanisms of TLS. Here we review the recent advances in the structural characterization of eukaryotic TLS DNA polymerases and their higher-order assemblies, providing new insights into the concerted action of TLS enzymes during the replicative bypass of DNA lesions.
In humans (Hs), enzymes of the TLS pathway include the Y-family DNA polymerases Polη, Polι Polκ, Rev1, and B-family Polζ, while S. cerevisiae (Sc) or baker’s yeast, which has long served as a model organism for studying TLS in eukaryotes, possesses Polη, Rev1, and Polζ only [15,16,17,18,19,20,21,22,23,24] (Figure 1A,B). Unlike Y-family DNA polymerases that comprise a single polypeptide chain, Polζ is a heteroprotein complex of the catalytic subunit Rev3, the two copies of HORMA domain subunit Rev7, and two accessory subunits, Hs PolD2/PolD3 or Sc Pol31/Pol32, that are also subunits of the replicative B-family DNA polymerase Polδ [37,38,39,40,41] (Figure 1). Beyond their catalytic domains, TLS polymerases feature accessory motifs, domains, and subunits that mediate protein–protein interactions (PPIs) involved in TLS regulation (Figure 1A,B) that control TLS polymerase recruitment and assembly into multiprotein complexes at sites of DNA of damage.
The catalytic domains of TLS DNA polymerases generally (i) have active sites that are adapted to accommodate DNA lesions, (ii) form overall fewer contacts with primer/template (p/t) DNA than the replicative polymerases, and (iii) are more prone to incorrect base pairing due to the lack of 3′-5′ exonuclease activity [23,42]. These properties provide TLS enzymes with the ability to replicate past sites of DNA damage at the expense of decreased replication fidelity, processivity, and efficiency on undamaged DNA [15,16,17,18,19,20,21,22,23,24]. While the nucleotide misincorporation rates of the replicative DNA polymerases Polδ and Polε are ~10–6–10–8, the misincorporation rates of the TLS DNA polymerases range from 10–1 to 10–4 [43,44].
The access of mutagenic TLS enzymes to DNA is regulated through PPIs with proliferating cell nuclear antigen (PCNA), a homo-trimeric sliding clamp that serves as a polymerase processivity factor and a scaffold for DNA replication and damage response proteins [45,46,47,48] (Figure 1C). Like other PCNA binding partners, Polη, Polι, Polκ, and the Hs PolD3/Sc Pol32 subunit of Polζ contain PCNA-interacting protein box (PIP-box) motifs that mediate recruitment of TLS polymerases to DNA damage-induced foci [49,50,51,52,53,54,55,56] (Figure 1A,B). In contrast, Rev1 lacks a PIP-box motif and instead binds PCNA via its BRCA1 C-terminus (BRCT) domain [57,58], which is preceded by an α-helical M1 region that reportedly interacts with the recessed 5′ phosphorylated terminus of ssDNA [59] (Figure 1A,B). The switch from high-fidelity replication to TLS is prompted by the mono-ubiquitination of PCNA at residue K164 by the Rad6 ubiquitin-conjugating enzyme (E2) and Rad18 ubiquitin-protein ligase (E3) pair, which is recruited to ssDNA coated with replication protein A (RPA) formed at stalled replication forks [60,61,62,63,64,65]. This post-translational modification enhances the affinity of PCNA for the Y-family TLS DNA polymerases, which bind a PCNA-attached ubiquitin moiety via ubiquitin-binding motif (UBM) or ubiquitin binding zinc-finger (UBZ) domains [66,67] (Figure 1A,B).
Individual TLS DNA polymerases can copy over certain DNA lesions, as exemplified by the accurate and efficient bypass of UV-induced T-T cyclobutane pyrimidine dimers (CPDs) by Polη [68]. However, the replicative bypass of most types of DNA damage generally requires a two-step Rev1/Polζ-dependent TLS [69,70,71,72], which includes (i) base insertion across from the lesion by Polη, Polι, or Polκ, and (ii) the extension of the structurally aberrant primer-template, typically handled by Polζ [69,70,71,72]. This process involves the coordinated action of several TLS DNA polymerases assembled into a complex called the TLS mutasome whereby PCNA and Rev1 serve as scaffolds (Figure 1C). Rev1 plays a key structural role in Rev1/Polζ-dependent TLS by coordinating polymerase recruitment, selection, and switching via PPIs of its C-terminal domain (Rev1-CT), while its limited catalytic activity as dCMP transferase is dispensable for TLS [73,74,75,76,77]. The Rev1-CT domain features two independent PPI interfaces, one for the Rev7 subunit of Polζ and one for Rev1-interacting regions (RIRs) of Hs Polη, Polι, Polκ, and the PolD3 subunit of Polζ that are missing in TLS proteins in yeast (Figure 1) [78,79,80,81,82,83,84,85].
The architecture of Polζ, which serves as a master ‘extender’ polymerase within the TLS mutasome [37], resembles that of other multi-subunit B-family enzymes such as the primase Polα or the replicative polymerases Polδ and Polε [24,86,87,88,89]. The Rev3 subunit of Polζ includes the catalytic domain followed by a cysteine-rich C-terminal domain (Rev3-CTD) that binds Zn and [4Fe-4S] cluster [90] via CysA and CysB motifs, respectively (Figure 1A,B) [37,38,39,40,41]. Rev3-CTD aligns with the corresponding region of Polδ harboring the [4Fe-4S] cluster [90], and mediates PPIs with the Hs PolD2/PolD3/Sc Pol31/Pol32 module shared between Polδ and Polζ [37,38,39,40,41]. The Rev3 subunit also features a long insert between its N-terminal domain (NTD) and an inactive 3′-5′ exonuclease domain [17,91] containing a pair of Rev7-binding motifs (RBMs) that bind two copies of the accessory subunit Rev7 [92,93,94] (Figure 1A,B). Both Hs and Sc Rev7 can form homodimers within Polζ, although the dimerization mode is reported to be different in each case [94,95]. The accessory Rev7 and Hs PolD2/PolD3/Sc Pol31/Pol32 subunits are important for Polζ function, significantly enhancing its catalytic efficiency [38,40,41].
This review focuses on structural studies underpinning the mechanisms by which TLS polymerases catalyze replication across DNA lesions in the context of multiprotein complexes. Following a brief overview of the key proteins of the TLS pathway provided in the introduction, we will discuss the structural basis of DNA synthesis by catalytic domains of Y-family Polη, Polι, Polκ, Rev1, and B-family Polζ revealed by X-ray crystallography studies (reviewed in [21,22,23,24]) and recent cryo-EM structures of the Sc Polζ holoenzyme [96,97,98,99,100]. While much is known about TLS PPIs, the majority of their structural studies have focused on individual domain–domain or domain–peptide interactions. Given the number of possible PPIs within the TLS mutasome, it represents a multivalent complex that adopts different configurations at each step of the lesion bypass, allowing for effective polymerase recruitment, selection, and switching. A structural picture of the full TLS mutasome in the act of nucleotide insertion across from the lesion or p/t DNA extension has yet to be realized. Hoverer, the recent landmark Cryo-EM structures of TLS holoenzyme complexes, including the structure of Hs Polκ in association with PCNA and DNA [96], the structures of Sc Polζ on and off p/t DNA [97,98,99], and the partial structure of the Rev1/Polζ complex during p/t extension [100], provide new mechanistic insights into the TLS DNA polymerase assemblies. Herein, we will overview the key findings of these works along with the previous structural studies of PPIs within the TLS mutasome, and discuss their implications for TLS mechanisms.
2. Structural Basis of DNA Synthesis by TLS Polymerases
2.1. Catalytic Domains of the Y-Family TLS Polymerases
Crystal structures of the catalytic cores of the Y-family TLS DNA polymerases have been determined with and without unmodified or damage-containing DNA substrates (reviewed in [21,22,23,24]). These structures have unveiled the mechanisms by which TLS DNA polymerases recognize, accommodate, and insert bases across various DNA lesions. The catalytic regions of Polη, Polι, Polκ, and Rev1 consist of finger, palm, and thumb domains common to all DNA polymerase families, and a C-terminal ‘little finger’ (LF) or polymerase associated domain (PAD), which is unique to Y-family polymerases (Figure 2A–D) [15,16,17,18,19,20,21,22,23,24]. The catalytic core structure resembles a right hand holding p/t DNA between the thumb and little finger domains positioning it for incoming dNTP. As with all DNA polymerase families, the catalytic site in the palm domain contains carboxylate residues that coordinate three Mg2+ ions that are needed to catalyze phosphodiester bond formation between the 3′-OH of the primer strand and α-phosphate of the incoming nucleotide [23,101,102]. Contributing to the ability to accommodate aberrant DNA, the finger and thumb domains of the Y-family enzymes are shorter and form fewer interactions with DNA compared to the replicative B-family polymerases, while the PAD domain serves to stabilize the templating base through contacts with the DNA backbone. Another contributing factor to the proficiency of the Y-family polymerases to incorporate nucleotides over DNA lesions is a lack of the 3′-5′ exonuclease domain and thus proofreading capability, which greatly decreases replication fidelity on undamaged DNA. It has also been noted that, unlike replicative polymerases, some Y-family polymerases (such as Hs Polη) preexist in a ready-for-catalysis state and undergo little conformational changes induced by nucleotide binding [103,104].
Like B-family replicative polymerases, the Y-family TLS polymerases feature steric gating residues within their active sites that allow for sugar discrimination during catalysis. By this mechanism, a bulky residue within the active site of a B- or Y-family DNA polymerase (typically Tyr or Phe) clashes with the 2′-OH of an incoming rNTP, averting ribonucleotide incorporation via steric exclusion [105,106,107,108,109,110]. Hs Rev1 is unique in that it utilizes a steric gate in combination with a polar filter residue, which interacts with the 3′-OH and brings the primer nucleotide closer to the steric gate [111]. In general, the incorporation efficiency of rNTP by TLS DNA polymerases is lower relative to dNTP; however, it varies across the different enzymes. For example, Sc Polη and Polζ are fairly intolerant of rNTPs and do not incorporate them frequently [109,112,113,114,115]. In contrast, Hs Polη incorporates dNTP and rNTPs with similar efficiency across 8-oxo-G and TT CPD, while Hs Polι was reported to incorporate rNTPs across abasic sites [109,112,116,117].
Despite having similar architecture, the catalytic cores of Y-family TLS polymerases have unique structural features and vary in interactions with DNA. For example, the catalytic region of Polκ includes an N-terminal extension (N-clasp) that links the core and PAD domains, stabilizing the latter on DNA (Figure 2B) [118]. In a similar manner, the core and PAD domains of Rev1 are not contiguous but held together by an N-terminal extension (N-digit), which makes further contacts with the DNA minor groove (Figure 2D) [119,120,121]. These structural details determine the types of DNA damage (cognate lesions) that each Y-family polymerase can recognize and accurately bypass [23].
Polη specializes in the bypass of cyclobutane pyrimidine dimers (CPDs) [68], which are the most common lesion induced by UV light, and interstrand crosslinks induced by platinum-based chemotherapies, such as cisplatin [122]. Crystal structures of the Hs Polη in complex with DNA containing a TT CPD [123] or platinum cross-linked guanines (GpG) (Figure 2A) [30,124] reveal that the active site of Polη constrains DNA in B-form, minimizing the distortions caused by the lesion and thus stabilizing Watson–Crick (WC) geometry with the correct incoming nucleotide. While Polη can insert dATP across TT CPD and dCTP across GpG in an error-free manner, it tends to misincorporate dGTP across T and has an error rate of 10−2–10−3 on undamaged DNA [125].
Polκ is particularly adept at the accurate bypass of DNA lesions induced by bulky polycyclic aromatic hydrocarbons (PAHs), such as benzo[a]pyrene guanine adducts (BPdG) [23]. The crystal structure of Polκ in complex with p/t DNA containing a BPdG lesion reveals an open active site adapted to accommodate the bulky minor groove adduct (Figure 2B) [126]. This property stems from the loose association between the catalytic core and PAD domains bridged by the N-clasp, which stabilizes Polκ on aberrant p/t DNA allowing it to insert the correct dCTP nucleotide. Following the nucleotide insertion, the BPdG adduct in the 5′ position translocates down one position and orients towards the DNA minor groove. Not only is this conformation energetically favorable, but the lesion also sterically clashes with mismatches in the nascent base pair, thus favoring correct pairing and allowing Polκ to extend past the lesion with reasonable accuracy.
Polι, unlike Polη and Polκ, has a much narrower active site, which binds and constrains template bases in a syn conformation and favors Hoogsteen over WC base pairing [23]. The constraints placed by its active site make Polι accurate across template purines (dA, dG), since favorable hydrogen bonds to dTTP and dCTP in an anti conformation are still formed along the Hoogsteen edge. However, this also makes Polι much more promiscuous when inserting dNTPs across template pyrimidines (dC, dT). Crystal structures of Hs Pol ι in complex with an 8-oxo-guanine (8-oxoG) lesion demonstrate how this property aids Polι in correctly incorporating dCTP across this lesion (Figure 2C) [127]. The 8-oxoG is stabilized in the syn conformation and forms optimal hydrogen bonds to dCTP in the anti conformation, thus promoting accurate lesion bypass.
Rev1 is unique among Y-family TLS polymerases in that it primarily functions as a dCMP transferase, preferentially inserting cytosine regardless of the template base [23,73]. Crystal structures of Hs Rev1 on undamaged DNA suggest this property stems from a protein templating mechanism (Figure 2D) [111,119]. The residue L358 on the N-digit of Hs Rev1 evicts the template base, orienting it away from the DNA helix and stabilizing it through interactions with H774 in the PAD domain. Hs Rev1 then uses R357 (R324 in Sc Rev1) on the N-digit as the new ‘template’, which forms WC-like hydrogen bonds with the incoming dCTP. This unusual mechanism enables Rev1 to bypass lesions such as adducted guanines or abasic sites.
2.2. Catalytic Domain of the B-Family TLS Polymerase Polζ
The catalytic domain of Polζ harbored in the Rev3 subunit has proven a particular challenge for structural characterization. Historically, due to its large size (3103 and 1505 amino acids for Hs and Sc Rev3) and tendency toward aggregation, the purification of Rev3 alone has been difficult, hindering efforts to obtain high-resolution structures [41,128]. In 2013, Gómez-Llorente et al. reported the first low-resolution EM structure of the multisubunit Sc Polζ complex at 22 Å resolution [128]. In 2020, Malik et al. presented breakthrough high-resolution Cryo-EM structures of the Sc Polζ holoenzyme on (3.1 Å) and off (4.1 Å) undamaged p/t DNA [97], followed by a structure of Polζ with a base pair mismatch at the primer terminus (3.1 Å) [98]. These studies not only uncovered the architecture of the heteropentameric Sc Polζ assembly, consisting of Rev3, Pol31, Pol32, and two copies of Rev7 (Rev7A and Rev7B) subunits, but also provided structural insights into the function of Polζ as a master extender polymerase during Rev1/Polζ-dependent TLS.
The catalytic core of Sc Polζ [97] bears resemblance to that of replicative B-family polymerases Polδ and Polε [24,86,87,88,89], featuring a palm domain that carries catalytic carboxylate residues, a finger domain that makes contacts to the nascent DNA pair, a thumb domain that interacts with the minor groove of DNA, an exonuclease domain which is inactive in Polζ, and an N-terminal domain (NTD) that bridges exonuclease and fingers domains (Figure 3A). The thumb, palm, and fingers of Rev3 grasp DNA tightly, creating an active site that is fairly intolerant to mismatches. Like high-fidelity replicative polymerases, Rev3 makes contacts to the nascent base pair via residues in its fingers and palm domain that enforce WC geometry at the templating position T0P0 (here T is template, P is primer; the subscript denotes the number of bases past the templating position) (Figure 3A, inset). However, Rev3 can tolerate mismatches and DNA lesions at the T1P1 position, which sets Polζ apart from the replicative polymerases and contributes to its extender function in TLS. While in the Pol3 subunit of Polδ, an NTD-palm linker spans along p/t and forms contacts constraining the T1 nucleotide; the NTD-palm linker in Rev3 moves away from the template, creating a cleft near the T1 position that can accommodate structurally aberrant DNA (Figure 3B). Another key feature that enables Polζ to extend past DNA lesions is an inactive Rev3 exonuclease domain, which lacks the carboxylate residues found in other B-family polymerases that mediate 3′-5′ exonuclease activity and misses a crucial β-hairpin that regulates the transition of mismatched p/t from the active site to the exonuclease domain for proofreading (Figure 3C).
In 2022, Malik et al. reported another Cryo-EM structure of the Sc Polζ holoenzyme, this time in complex with a mismatched DNA primer-template [98]. While the overall architecture of Polζ is preserved in this structure, the fingers domain adopts a more open conformation and, based on a less defined EM density, becomes more dynamic, loosening the enzyme’s grip on DNA and allowing it to accommodate the mismatch. Despite a reduced number of contacts with the active site, the nascent base pair is still constrained in WC geometry by some interactions with the fingers domain, revealing how Polζ may catalyze DNA synthesis past the mismatch. Taken together, the structures of Sc Polζ on matched and mismatched p/t DNA have unmasked the structural determinants of Polζ function during the extension step of TLS.
3. Protein Interaction Modules—The Building Blocks of TLS Complexes
Catalytic domains of TLS polymerases furnish an enzymatic toolbox for the replicative bypass of DNA lesions. In addition, the recruitment of TLS polymerases to DNA, selection of polymerase(s) appropriate for the type of damage, and polymerase switching are governed by a network of PPIs mediated by accessory motifs, domains, and subunits of the TLS enzymes (Figure 1). Structural studies of individual PPIs within this network mainly focused on the TLS scaffold proteins, PCNA and Rev1, and the Rev7 subunit of Polζ. These works portrayed the building blocks of the multiprotein TLS complexes and provided insights into TLS regulation.
3.1. Interactions with PCNA
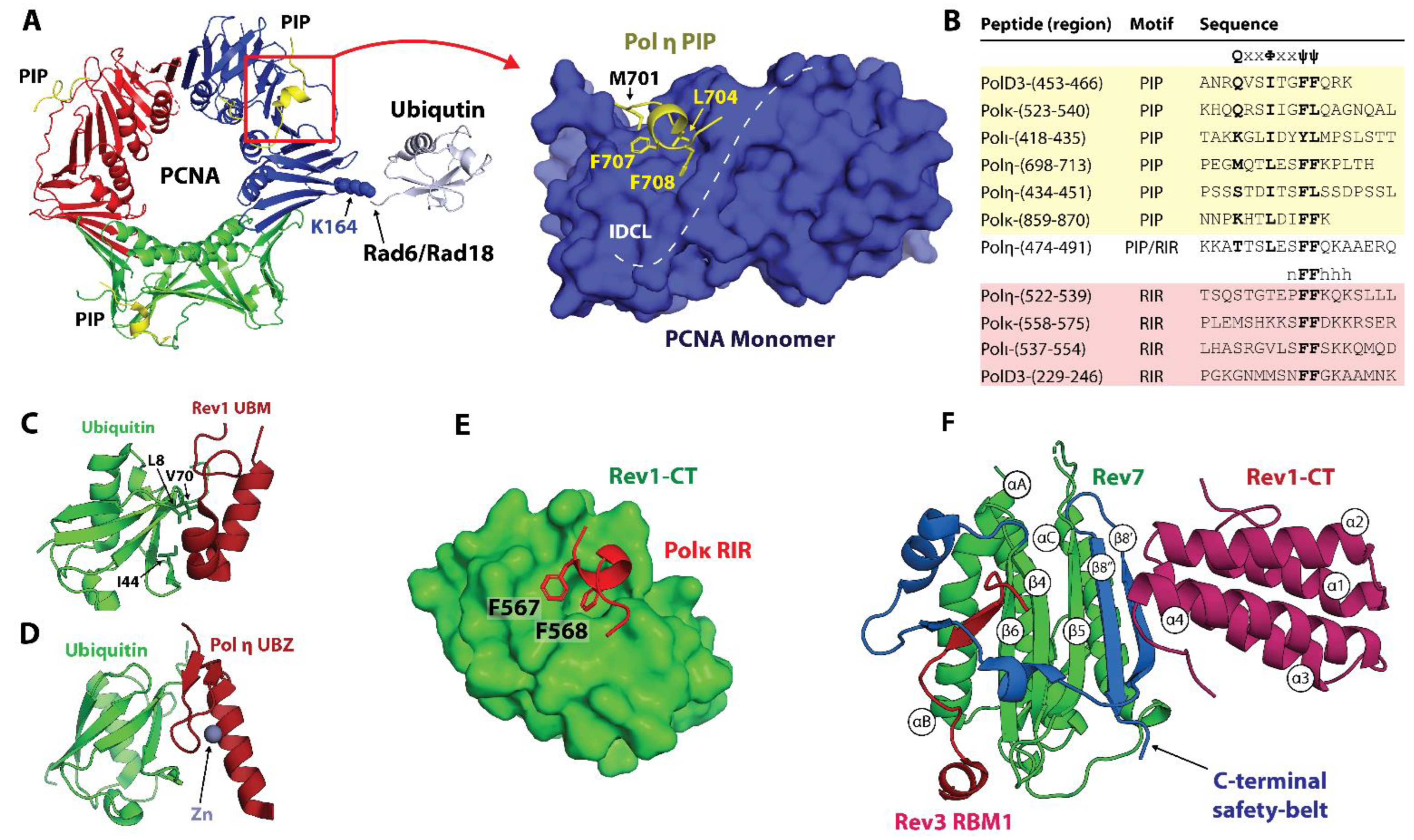
The sliding clamp PCNA is a polymerase processivity factor and an interaction hub that orchestrates protein recruitment and transactions taking place on DNA during replication and repair [45,49,50]. Eukaryotic PCNA is a ring-shaped trimer of identical subunits that encircles DNA, with each subunit consisting of the two domains linked by the interdomain connector loop (IDCL) (Figure 4A). Most PCNA partners harbor one or several PCNA-interacting protein box (PIP-box) motifs with the consensus sequence Q1x2x3φ4x5x6ψ7ψ8 (where φ is I/L/M/V, ψ is F/Y, x is any residue), which bind a conserved site in the IDCL region of PCNA [49,50,51,52,53,54,55,56] (Figure 4A,B). Upon binding, the PIP-box forms a helix that orients residues in positions 4, 7, and 8 (the ‘three-pronged plug’) towards a hydrophobic pocket on PCNA, while Q in position 1 inserts into a site termed the ‘Q-pocket’. For the tightest-binding PIP-box motifs such as the one from p21WAF1/CIP1 (Kd for Hs PCNA of 80 nM [50]), the PPI is enhanced by residues C-terminal to the consensus sequence that forms an intermolecular β-sheet with the IDCL of PCNA and/or other peripheral contacts [50,129].
The Y-family TLS polymerases Polη, Polι, and Polκ bind PCNA using non-canonical PIP-box motifs that deviate from the consensus sequence [51,52,53,54,55,56,130] (Figure 1 and Figure 4B). In contrast, the B-family TLS polymerase Polζ borrows a canonical PIP-box from the Hs PolD3/Sc Pol32 subunit shared with the replicative DNA polymerase Polδ (Figure 1). Unlike other Y-family polymerases, Rev1 lacks a PIP-box and binds PCNA via its N-terminal BRCT domain (Figure 1) [57,58]. Crystal structures of Hs PCNA in complex with PIP-box peptides derived from Hs Polη, Polι, and Polκ reveal that these non-canonical motifs bind the IDCL region of PCNA using the ‘three-pronged fork’ mechanism, while the residue in position 1 lacks (Polι, Polκ) or forms suboptimal (Hs Polη) interactions with the ‘Q-pocket’ (Figure 4A, left) [130]. The overall fewer contacts with PCNA lead to weaker PCNA binding affinities for non-canonical sequences (Kd in the μM range) relative to the canonical PIP-boxes [50,129,130]. The relatively weak PPIs of non-canonical PIP-boxes with PCNA promote TLS polymerase recruitment to DNA [51,52,53,54,55,56], while also permitting the selection and switching of the TLS polymerases as they compete for space on the sliding clamp.
In addition to PIP-box motifs, all Y-family TLS polymerases have ubiquitin-binding UBZ and UBM domains lacking in replicative polymerases (Figure 1). Rad6/Rad18-dependent mono-ubiquitination of PCNA at sites of DNA damage leads to UBM and UBZ binding to PCNA-attached ubiquitin moieties and promotes the replacement of replicative with TLS polymerases by an affinity-driven competition [66,67]. Polη and Polκ harbor one and two UBZ domains, respectively (Figure 1 and Figure 4D), while Polι and Rev1 each feature two UBM domains (Figure 1 and Figure 4C). Solution NMR studies reveal that the Hs Polη UBZ domain adopts a canonical ββα type-III C2H2 zinc-finger fold coordinating a single Zn2+ ion [131,132], while Hs Rev1 and Polι UBM domains have an αα structure [133,134,135,136]. Both UBZ and UBM domains engage a conserved site on ubiquitin centered at L8, I44, and V70 (Figure 4E), although mutation of I44 is not sufficient to disrupt the binding of the Rev1 UBM2 domain [135]. Notably, only one of the two UBMs in Rev1 (UBM2) was shown to interact with ubiquitin in both humans and yeast [135].
Figure 4.
Protein-protein interactions in TLS. (A) Hs PCNA with a Polη PIP-box peptide bound near the interdomain connector loop (IDCL) [130]. Ubiquitin (gray) is ligated at K164 of PCNA by the Rad6/Rad18 E2/E3 pair. A close-up view of the PCNA monomer showing a “three-pronged fork” of Polη PIP (L704, F707, F708) inserting into a hydrophobic pocket in the IDCL region of PCNA, and M701 binding to the Q-pocket. (B) PIP-box and RIR sequences in the human TLS polymerases aligned to their minimal motifs. (C) Hs Rev1 UBM2 bound to a conserved site on ubiquitin centered at L8, I44, and V70 [135]. (D) A model of Hs Polη UBZ bound to ubiquitin (based on the Rad18-UBZ/ubiquitin complex) [132,137]. (E) Hs Polκ RIR motif bound to Rev1-CT with F567 and F568 inserted in a hydrophobic pocket in the N-terminal part of the domain [85]. (F) Ternary complex of Hs Rev7, Rev3 RBM1, and Rev1-CT [83]. Rev7 uses a C-terminal ‘safety-belt’ loop (blue) to latch the Rev3 RBM1 peptide to the core. The C-terminus of the ‘safety-belt’ region forms a β8′-β8′’ hairpin, which extends the core β-sheet (β4-6) and provides the binding interface for Rev1-CT.
Figure 4.
Protein-protein interactions in TLS. (A) Hs PCNA with a Polη PIP-box peptide bound near the interdomain connector loop (IDCL) [130]. Ubiquitin (gray) is ligated at K164 of PCNA by the Rad6/Rad18 E2/E3 pair. A close-up view of the PCNA monomer showing a “three-pronged fork” of Polη PIP (L704, F707, F708) inserting into a hydrophobic pocket in the IDCL region of PCNA, and M701 binding to the Q-pocket. (B) PIP-box and RIR sequences in the human TLS polymerases aligned to their minimal motifs. (C) Hs Rev1 UBM2 bound to a conserved site on ubiquitin centered at L8, I44, and V70 [135]. (D) A model of Hs Polη UBZ bound to ubiquitin (based on the Rad18-UBZ/ubiquitin complex) [132,137]. (E) Hs Polκ RIR motif bound to Rev1-CT with F567 and F568 inserted in a hydrophobic pocket in the N-terminal part of the domain [85]. (F) Ternary complex of Hs Rev7, Rev3 RBM1, and Rev1-CT [83]. Rev7 uses a C-terminal ‘safety-belt’ loop (blue) to latch the Rev3 RBM1 peptide to the core. The C-terminus of the ‘safety-belt’ region forms a β8′-β8′’ hairpin, which extends the core β-sheet (β4-6) and provides the binding interface for Rev1-CT.
3.2. Scaffolding Function of Rev1
The two unique modular domains in Rev1 that are not found in other Y-family polymerases, Rev1-BRCT and Rev1-CT (Figure 1), enable its function as a central scaffold protein of the TLS pathway [57,74,77,82,84]. In 1971, the rev1-1 strain of yeast was discovered, which contains the G193R mutation in Rev1, displaying a DNA damage-sensitive phenotype and a marked decrease in mutagenesis [138]. This mutation maps onto the BRCT region of Sc Rev1 and has no effect on catalysis, which has led to the suggestion of a ‘second’ structural role of Rev1 in TLS [74]. The Hs Rev1-BRCT domain promotes resistance to DNA damage and mediates Hs Rev1 localization to replication foci [57,139]. While the structures of both Hs and Sc Rev1-BRCT have been determined [58,140], the interactions of this domain have long remained unknown, with tentative partners including PCNA [57,58], DNA [59], and phosphopeptides [141]. The direct interaction of Sc Rev1-BRCT and PCNA with Kd of ~79 μM was demonstrated by NMR, revealing that the BRCT domain competes with PIP-box motifs for the same site in the IDCL region of PCNA [58]. The precise structural mechanism by which PCNA can accommodate Rev1-BRCT in a binding site reserved for other partners remains to be established.
The Rev1-CT domain is a versatile interaction module that mediates Rev1 PPIs with other Y-family TLS polymerases and the accessory subunits of the B-family TLS polymerase Polζ [57,74,77,82,84]. The two independent interfaces of Hs Rev1-CT can simultaneously bind the Rev7 subunit of Polζ and the Rev1-interacting regions (RIRs) of Hs Polη, Polι, and Polκ (Figure 1 and Figure 4B) [78,79,80,81,82,83,84,85], bridging the extender and inserter polymerases during Rev1/Polζ-dependent TLS. In addition, an RIR motif has been identified in the Hs PolD3 subunit of Polζ, which is implicated in an inserter-to-extender polymerase switch (Figure 1 and Figure 4B) [82]. The RIR motif has a consensus sequence of nFFhhhh, where h is a helix-forming residue, and n is an N-capping residue [142] (Figure 4B) [78,80]. Solution NMR structures of Rev1-CT bound to RIR peptides from Hs Polη [80] and PolD3 [82] reveal that, upon binding, the RIR motif folds into an α-helix, directing the FF pair towards a pocket in the N-terminal part of Rev1-CT (Figure 4E). Crystal structures of the Rev1-CT/Rev7 complexes show that Rev7 binds to the opposite C-terminal face of Rev1-CT without obstructing the RIR interface (Figure 4F) [83,84,85]. The overlapping PIP-box and RIR motifs were identified in human TLS polymerases sharing a pair of aromatic residues (Figure 4B), challenging the notion of PIP and RIR as distinct entities [143]. Notably, the TLS polymerases in yeast lack a putative RIR motif, although Sc Rev1-CT can bind an RFFD sequence from Sc Rad5 in a manner similar to Hs Rev1-CT/RIR PPI [144]. It has been proposed, therefore, that Sc Polη utilizes its C-terminal PIP-box motif for Rev1-CT binding [145].
3.3. Interactions of Polζ Subunits
Previous structural studies of Polζ mainly focused on interactions of its accessory Rev7 subunit with the Rev7-binding motifs (RBMs) from Rev3 [95,146] and with the Rev1-CT domain [83,84,85], although structures of the Hs PolD2/PolD3 and Sc Pol31/Pol32 accessory modules are also available [86,147]. The two consecutive RBMs were identified in both Hs and Sc Rev3 located in the long insert between its NTD and exonuclease domains (Figure 1) [92,93,94]. In humans, the core RBM sequence is defined as ΦΦψPxxxpP, where Φ is an aromatic or bulky aliphatic residue, ψ is a residue with a long aliphatic side-chain, n is an N-capping residue [142], while in yeast the first conserved P may be replaced with Y [94]. Rev7 interacts with RBMs using a mechanism unique to HORMA proteins whereby the Rev7 ‘safety-belt’ loop closes on the top of the ligand, securing it to the protein core by the β8′-β8′′ hairpin, which extends the Rev7 β-sheet (Figure 4F) [95,146]. This newly formed β-hairpin creates a binding site of Rev7 for the Rev1-CT domain [83,84,85]. Within Polζ, the two sequential RBMs of Rev3 recruit two copies of Rev7 [94,95]. The two Hs Rev7 protomers tethered by the Rev3 fragment form a symmetric head-to-head dimer via the canonical HORMA interface centered at helix αC [95], while the two copies of Sc Rev7 are forced into an unusual head-to-head dimer in the Sc Polζ assembly [94,98]. The Rev3/Rev7 complex formation results in a ~20–30 fold increase in the Rev3 catalytic activity [148], which is further improved by the addition of the Hs PolD2/PolD3 or Sc Pol31/Pol32 block [38,41]. The loss of Hs Rev7 PPIs with Rev3-RBMs [92] or Hs Rev7 dimerization [95] sensitizes cells to DNA damage induced by UV and cisplatin, highlighting the importance of Rev7 as a key interaction module of the Polζ assembly.
4. Molecular Assemblies in TLS
The recent advent of high-resolution Cryo-EM has opened an avenue for the structural characterization of multiprotein assemblies acting in TLS. Following the high-resolution structures of the multisubunit Polδ and Polε holoenzymes [86,87,88,89], structures have been determined for TLS complexes, including full-length Hs Polκ bound to PCNA and p/t DNA [96] (Figure 5), the Sc Polζ holoenzyme on and off matched and unmatched p/t DNA [97,98,99], and the Rev1/Polζ complex during p/t extension [100] (Figure 6). These structures captured TLS machines in action, piecing together the catalytic and regulatory modules and unveiling how TLS enzymes cooperate during replication and lesion bypass.
4.1. Polκ on PCNA in the Act of Catalysis
In 2021, Lancey et al. [96] reported the high-resolution structures of full-length Hs Polκ in association with p/t DNA and unmodified (3.4 Å) or ubiquitinated (6.4 Å) PCNA, providing the first view of a Y-family TLS polymerase in association with PCNA during catalysis (Figure 5A,B). The Cryo-EM structures of the Polκ catalytic core, including palm, thumb, fingers, and PAD domains with p/t DNA in the active site, align well with the previous X-ray crystal structures [118], albeit DNA orientation below the PAD domain deviates from that in the crystal structure and the N-clasp is poorly resolved in Cryo-EM maps. In both structures, Polκ sits on the front face of PCNA and engages a single PCNA protomer through an internal PIP-box motif (PIP1) following the Polκ PAD domain (Figure 1 and Figure 5A). A short linker between Polκ PAD and PIP1 forms an α-helix (the ‘inverting helix’), which contributes 733 Å2 to the buried surface area of the PCNA/Polκ interface (Figure 5A, right). The C-terminal part of Polκ beyond the PIP1 motif, including the UBZ domains and the PIP2 motif, is invisible in Cryo-EM maps, suggesting this region is dynamic.
The structure of Polκ in complex with p/t DNA and ubiquitinated PCNA reveals a transient nature of PPIs between Polκ UBZs and PCNA-attached ubiquitin moieties [96]. While the UBZ domains are invisible, weak electron density is present on the back face of PCNA, corresponding to a ubiquitin moiety attached to each protomer at residue K164 (Figure 5B). The best-defined density is for ubiquitin attached to the PCNA protomer bound to Polκ, which is more accessible for interactions with Polκ UBZs. The authors demonstrated that the synthetic activity of the Polκ PIP1 mutant is severely impaired relative to wild-type Polκ in the presence of unmodified PCNA, but is only moderately reduced in the presence of ubiquitinated PCNA. Consistent with the previous functional studies [53,54], this result implies that the internal PIP-box serves as the major tether of Polκ to PCNA, while the UBZ domains supply accessory contacts, partially rescuing Polκ activity when the PIP-box interaction is impaired.
The most striking feature of the Polκ holoenzyme bound to PCNA and p/t DNA is its tilted orientation relative to the clamp, such that DNA exits the catalytic core at a 47° angle relative to the PCNA axis (Figure 5C) [96]. As a result, DNA that extends beyond the core bends by 30° to thread through the PCNA ring and avoid clashing with its inner rim. While the role of such DNA bending remains unclear, the authors suggest it may facilitate the assembly of PCNA ‘tool belts’ [47] whereby several DNA processing enzymes can engage the clamp at the same time and handover DNA without dissociation. An example of such a ‘tool belt’ is provided by the Cryo-EM structure of Hs Polδ and Flap endonuclease 1 (FEN1) bound to different protomers of the same PCNA ring [88]. The tilted orientation of Polκ relative to PCNA observed by Cryo-EM may serve to shift Polκ away and make space on the clamp to accommodate another DNA polymerase, as illustrated by modeling Polκ and Polδ onto a single PCNA molecule (Figure 5D).
4.2. Polζ Holoenzyme and Rev1-Polζ Assembly—A View of the TLS Mutasome
The first high-resolution Cryo-EM structures of the Sc Polζ holoenzyme on p/t DNA [97,98,99] reveal a pentameric “daisy-chain” or ring of Rev3, Pol31, Pol32, and two copies of Rev7 (Rev7A and Rev7B) in a dimer (Figure 6A). In the Polζ structure, the catalytic subunit Rev3 alone interacts with DNA, and forms contacts with all other subunits but Pol32, while the accessory subunits arrange into a regulatory module facing away from DNA. Rev3 utilizes its C-terminal CysB domain to recruit the Pol31/Pol32 module via PPIs with Pol31, and binds a dimer of Rev7A and Rev7B subunits with its RBM1 and RBM2 motifs. The Rev3 segment connecting RBM1 and RBM2 forms additional contacts with Rev7 protomers and bridges the Rev7 dimer with the palm and thumb domains of the Rev3 catalytic core. Rev7B interacts with the Pol31/Pol32 module near the interface of Pol31 and the N-terminal domain of Pol32 (Pol32N) closing the Polζ ring. The pentameric ring structure of Polζ is unique among the B-family polymerases in that the accessory subunits of Polζ are organized in a more rigid manner [86,87,88,89]. While Sc Polδ adopts a two-lobed structure comprising the globular catalytic subunit Pol3 and a loosely attached regulatory Pol31/Pol32 N module [86], Pol31 and Pol32N within Sc Polζ are stabilized by extensive contacts with the CysB and exonuclease domains of Rev3 and with Rev7B (Figure 6A). The Pol32 C-terminal region (Pol32C) containing a terminal PIP-box motif is disordered. However, given the Pol32 orientation in the Pol ζ structure, Pol32C points outwards and could extend to bind PCNA (Figure 6A).
Figure 6.
Cryo-EM structures of the Sc Polζ and Sc Rev1/Polζ complex. (A) “Daisy-chain” or ring-like architecture of the five-subunit Sc Polζ holoenzyme on p/t DNA. Rev3 accommodates p/t DNA (red) and makes contacts to Pol31, Rev7A, and Rev7B but not Pol32 [97]. Rotation of the structure shows an unstructured C-terminus of Pol32, which contains a PIP-box motif, reaching a tentative PCNA location. (B) An unusual head-to-tail Sc Rev7 dimer within Polζ (top) that does not utilize the canonical HORMA dimerization interface [97]. For comparison, a head-to-head dimer is shown (bottom) formed by Hs Rev7 within the shielding complex [149]. Here the heterodimer of closed (C-Rev7) and open (O-Rev7) protomers is formed via the interface centered at helix αC. (C) Partial structure of the Sc Rev1/Polζ complex in which the Sc Polζ holoenzyme is engaged with p/t DNA [100]. Only the Rev1 CT, BRCT, and M1 regions are visible. The PCNA ring is modeled based on the structure of Polδ/PCNA complex [87]. A close-up view of the Rev1-M1 and Rev1-BRCT interactions with p/t DNA is shown on the right [100].
Figure 6.
Cryo-EM structures of the Sc Polζ and Sc Rev1/Polζ complex. (A) “Daisy-chain” or ring-like architecture of the five-subunit Sc Polζ holoenzyme on p/t DNA. Rev3 accommodates p/t DNA (red) and makes contacts to Pol31, Rev7A, and Rev7B but not Pol32 [97]. Rotation of the structure shows an unstructured C-terminus of Pol32, which contains a PIP-box motif, reaching a tentative PCNA location. (B) An unusual head-to-tail Sc Rev7 dimer within Polζ (top) that does not utilize the canonical HORMA dimerization interface [97]. For comparison, a head-to-head dimer is shown (bottom) formed by Hs Rev7 within the shielding complex [149]. Here the heterodimer of closed (C-Rev7) and open (O-Rev7) protomers is formed via the interface centered at helix αC. (C) Partial structure of the Sc Rev1/Polζ complex in which the Sc Polζ holoenzyme is engaged with p/t DNA [100]. Only the Rev1 CT, BRCT, and M1 regions are visible. The PCNA ring is modeled based on the structure of Polδ/PCNA complex [87]. A close-up view of the Rev1-M1 and Rev1-BRCT interactions with p/t DNA is shown on the right [100].

Within the Sc Polζ holoenzyme, Rev7A and Rev7B form an unexpected head-to-tail dimer, not seen before in other HORMA proteins (Figure 6B, top). In this dimer, the αC helices of Rev7A and Rev7B are spaced far apart, with the new interface formed between the αC region of Rev7A and the C-terminal safety-belt region of Rev7B (Figure 6B, top). In contrast, Hs Rev7, alone [95] or within the shieldin complex [149] acting in DSB repair, forms a symmetric head-to-head dimer via the canonical HORMA interface centered at the helix αC (Figure 6B, bottom). Malik et al. [97] noted that the entire Rev7 interacting region of Rev3 within Sc Polζ “weave a path” through the two Rev7 subunits, making extensive contacts to both protomers and likely forcing them into an unusual head-to-tail dimer. This view is supported by a biophysical study of Sc Rev7 revealing that, unlike its human counterpart, the isolated Sc Rev7 lacks the ability to dimerize in solution, which is a property acquired later in evolution [94]. Interestingly, only the Rev7B subunit within Sc Polζ can accommodate Rev1-CT, while the respective site on Rev7A is occluded, suggesting Sc Polζ binds Rev1 in 1:1 stoichiometry.
In 2024, Malik et al. [100] reported a Cryo-EM structure (3.5 Å) of the Sc Rev1/Polζ holocomplex during p/t DNA extension by the Rev3 subunit of Polζ. The ring-like architecture of Sc Polζ is well preserved in this complex, while only three regions of a single Rev1 protein bound to Polζ are resolved, including Rev1-CT, Rev1-BRCT, and a Rev1-M1 α-helix preceding the BRCT domain (Figure 6C). The Rev1 catalytic core and the following region, including UBM domains (Figure 1), were flexible and invisible in the Cryo-EM map. As expected, Rev1-CT uses one face to bind the Rev7B subunit of Polζ, while also making contacts with Pol32N and a Rev3 segment connecting the two RBMs, and leaves the opposite face open for PPIs with the Y-family TLS polymerases (Figure 6C). Remarkably, the N-terminal Rev1-M1 α-helix and the following Rev1-BRCT domain form extensive contacts with the NTD-palm linker in the Rev3 catalytic domain, and with the primer and template strands of DNA (Figure 6C, right), which is consistent with previous observations [59]. This module is located similarly to the PAD domain in the Y-family TLS polymerases, tightening their grip on DNA. Furthermore, modeling of PCNA onto the Sc Rev1/Polζ structure reveals that Rev1-BRCT is primed for binding to the IDCL region of PCNA (Figure 6C), which is in line with NMR studies of the Sc Rev1-BRCT/PCNA PPIs [58]. Overall, the structure of the Sc Rev1/Polζ complex has uncovered the architecture of this essential module of the TLS mutasome and provided insights into the cooperation of TLS polymerases during DNA lesion bypass.
5. Concluding Remarks
Over the past two decades, a wealth of structural information has been amassed unveiling the mechanisms of catalysis and the interaction network of TLS DNA polymerases, which complements ample functional data available for these enzymes. However, until recently the mechanistic understanding of how the TLS polymerases assemble into multiprotein complexes and cooperate within supramolecular assemblies has markedly lagged behind. The recent high-resolution Cryo-EM structures of the multisubunit replicative [86,87,88,89] and TLS [96,97,98,99,100] DNA polymerases and their higher-order assemblies have been a remarkable feat in clarifying their activity on damaged and undamaged DNA. For example, the structural studies of Hs Polκ [96] and Polδ [87,88] holoenzymes on a sliding clamp PCNA have provided new insights into PCNA ‘tool belts’ [47], which facilitate DNA transactions between polymerases attached to the same clamp. The structures of the Sc Polζ holoenzyme and Rev1/Polζ holocomplex [96,97,98,99,100] unraveled the mechanism of p/t extension by Polζ and provided the first glimpse of the TLS mutasome in action. In addition, the Sc Polζ and Rev1/Polζ structures have elucidated the functional significance of already-characterized PPIs and described new interfaces and interactions. Because of their involvement in the replicative bypass of DNA adducts formed by genotoxic chemotherapeutics, which promotes drug resistance in cancers [25,26,27,28,29,30,31], TLS DNA polymerases have emerged as promising anti-cancer drug targets [32,33,34,35,36]. To date, efforts to develop small-molecule TLS inhibitors have mainly focused on targeting key PPIs within the TLS mutasome mediated by Rev1-CT and Rev7 [150,151,152,153,154,155,156,157]. The newly characterized TLS interfaces, therefore, extend the range of targets for TLS inhibition, providing new opportunities for anti-cancer drug design.
Author Contributions
G.A.A. and D.M.K. prepared the graphics and wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Institutes of Health grant R01CA233959 to M. Kyle Hadden and Dmitry M. Korzhnev.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
The authors thank Irina Bezsonova for help with the preparation of the manuscript graphics.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Friedberg, E.C.; Wood, R.D.; Walker, G.C.; Schultz, R.A.; Slede, W.; Ellenberger, T. DNA Repair and Mutagenesis, 2nd ed.; ASM Press: Washington, DC, USA, 2006. [Google Scholar]
- De Bont, R.; van Larebeke, N. Endogenous DNA damage in humans: A review of quantitative data. Mutagenesis 2004, 19, 169–185. [Google Scholar] [CrossRef]
- Bjelland, S.; Seeberg, E. Mutagenicity, toxicity and repair of DNA base damage induced by oxidation. Mutat. Res. 2003, 531, 37–80. [Google Scholar] [CrossRef]
- Davies, R.J.H. Ultraviolet radiation damage in DNA. Biochem. Soc. Trans. 1995, 23, 407–418. [Google Scholar] [CrossRef]
- Chatterjee, N.; Walker, G.C. Mechanisms of DNA damage, repair, and mutagenesis. Env. Mol. Mutagen. 2017, 58, 235–263. [Google Scholar] [CrossRef]
- Krokan, H.E.; Bjørås, K. Base excision repair. Cold Spring Harb. Perspect. Biol. 2013, 5, a012583. [Google Scholar] [CrossRef] [PubMed]
- Schärer, O.D. Nucleotide excision repair in eukaryotes. Cold Spring Harb. Perspect. Biol. 2013, 5, a012609. [Google Scholar] [CrossRef] [PubMed]
- Vilenchik, M.M.; Knudson, A.G. Endogenous DNA double-strand breaks: Production, fidelity of repair, and induction of cancer. Proc. Natl. Acad. Sci. USA 2003, 100, 12871–12876. [Google Scholar] [CrossRef]
- Povirk, L.F. Biochemical mechanisms of chromosomal translocations resulting from DNA double-strand breaks. DNA Rep. 2006, 5, 1199–1212. [Google Scholar] [CrossRef] [PubMed]
- Cannan, W.J.; Pederson, D.S. Mechanisms and consequences of double-strand DNA break formation in chromatin. J. Cell. Physiol. 2016, 231, 3–14. [Google Scholar] [CrossRef]
- Ramsden, D.A.; Nussenzweig, A. Mechanisms driving chromosomal translocations: Lost in time and space. Oncogene 2021, 40, 4263–4270. [Google Scholar] [CrossRef]
- Ulrich, H.D. Conservation of DNA damage tolerance pathways from yeast to humans. Biochem. Soc. Trans. 2007, 35, 1334–1337. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.J.; Cimprich, K.A. DNA damage tolerance: When it’s OK to make mistakes. Nat. Chem. Biol. 2009, 5, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Zeman, M.K.; Cimprich, K.A. Causes and consequences of replication stress. Nat. Cell Biol. 2014, 16, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Waters, L.S.; Minesinger, B.K.; Wiltrout, M.E.; D’Souza, S.; Woodruff, R.V.; Walker, G.C. Eukaryotic translesion polymerases and their roles and regulation in DNA damage tolerance. Microbiol. Mol. Biol. Rev. 2009, 73, 134–154. [Google Scholar] [CrossRef] [PubMed]
- Sale, J.E.; Lehmann, A.R.; Woodgate, R. Y-family DNA polymerases and their role in tolerance of cellular DNA damage. Nat. Rev. Mol. Cell Biol. 2012, 13, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Vaisman, A.; Woodgate, R. Translesion DNA polymerases in eukaryotes: What makes them tick? Crit. Rev. Biochem. Mol. Biol. 2017, 52, 274–303. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, A.A.; Korzhnev, D.M. The Rev1-Polζ translesion synthesis mutasome: Structure, interactions and inhibition. Enzymes 2019, 45, 139–181. [Google Scholar] [PubMed]
- Jacobs, J.J.L.; Paniagua, I. Freedom to err: The epanding celullar functions of translesion DNA polymerases. Mol. Cell 2023, 83, 3608–3621. [Google Scholar]
- Khatib, J.B.; Nicolae, C.M.; Moldovan, G.L. Role of translesion DNA synthesis in the metabolism of replication-associated nascent strand gaps. J. Mol. Biol. 2024, 436, 168275. [Google Scholar] [CrossRef]
- Pata, J.D. Structural diversity of the Y-family DNA polymerases. Biochim. Biophys. Acta 2010, 1804, 1124–1135. [Google Scholar] [CrossRef]
- Yang, W. An overview of Y-family DNA polymerases and a aase study of human DNA polymerase η. Biochemistry 2014, 53, 2793–2803. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Gao, Y. Translesion and repair DNA polymerases: Diverse structure and mechanism. Annu. Rev. Biochem. 2018, 87, 239–261. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Aggarwal, A.K.; Rechkoblit, O. Eukaryotic DNA polymerases. Curr. Opin. Struct. Biol. 2018, 53, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.S.; Takata, K.-I.; Wood, R.D. DNA polymerases and cancer. Nat. Rev. Cancer 2011, 11, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Lin, X.; Okuda, T.; Howell, S.B. DNA polymerase zeta regulates cisplatin cytotoxicity, mutagenicity, and the rate of development of cisplatin resistance. Cancer Res. 2004, 64, 8029–8035. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Trang, J.; Okuda, T.; Howell, S.B. DNA Polymerase ζ accounts for the reduced cytotoxicity and enhanced mutagenicity of cisplatin in human colon carcinoma cells that have lost DNA mismatch repair. Clin. Cancer Res. 2006, 12, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, S.Y.; Wang, S.; Lu, J.; Wu, W.; Weng, L.; Chen, D.; Zhang, Y.; Zhipeng, L.; Yang, J.; et al. REV3L confers chemoresistance to cisplatin in human gliomas: The potential of its RNAi for synergistic therapy. Neuro-Oncology 2009, 11, 790–802. [Google Scholar] [CrossRef] [PubMed]
- Xie, K.; Doles, J.; Hemann, M.T.; Walker, G.C. Error-prone translesion synthesis mediates acquired chemoresistance. Proc. Natl. Acad. Sci. USA 2010, 107, 20792–20797. [Google Scholar] [CrossRef]
- Zhao, Y.; Biertümpfel, C.; Gregory, M.T.; Hua, Y.-J.; Hanaoka, F.; Yang, W. Structural basis of human DNA polymerase η-mediated chemoresistance to cisplatin. Proc. Natl. Acad. Sci. USA 2012, 109, 7269–7274. [Google Scholar] [CrossRef]
- Vassel, F.M.; Bian, K.; Walker, G.C.; Hemann, M.T. Rev7 loss alters cisplatin response and increases drug efficacy in chemotherapy-resistance lung cancer. Proc. Natl. Acad. Sci. USA 2020, 117, 28922–28924. [Google Scholar] [CrossRef]
- Korzhnev, D.M.; Hadden, K. Targeting the translesion synthesis pathway for the development of anti-cancer chemotherapeutics. J. Med. Chem. 2016, 59, 9321–9336. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-S.; Kim, S.; Motea, E.; Berdis, A. Inhibiting translesion DNA synthesis as an approach to combat drug resistance to DNA damaging agents. Oncotarget 2017, 8, 40804–40816. [Google Scholar] [CrossRef]
- Yamanaka, K.; Chatterjee, N.; Hemann, M.T.; Walker, G.C. Inhibition of mutagenic translesion synthesis: A possible strategy for improving chemotherapy? PLoS Genet. 2017, 13, e1006842. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.M.; Dash, R.C.; Hadden, M.K. Translesion synthesis inhibitors as a new class of cancer chemotherapeutics. Expert Opin. Investig. Drugs 2021, 30, 13–24. [Google Scholar] [CrossRef]
- McPherson, K.S.; Korzhnev, D.M. Targeting protein-protein interactions in the DNA damage response pathways for cancer chemotherapy. RSC Chem. Biol. 2021, 2, 1167–1195. [Google Scholar] [CrossRef] [PubMed]
- Makarova, A.V.; Burgers, P.M. Eukaryotic DNA polymerase ζ. DNA Rep. 2015, 29, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Makarova, A.V.; Stodola, J.L.; Burgers, P.M. A four-subunit DNA polymerase ζ complex containing Pol δ accessory subunits is essential for PCNA-mediated mutagenesis. Nucleic Acids Res. 2012, 40, 11618–11626. [Google Scholar] [CrossRef] [PubMed]
- Baranovskiy, A.G.; Lada, A.G.; Siebler, H.M.; Zhang, Y.; Pavlov, Y.I.; Tahirov, T.H. DNA polymerase δ and ζ switch by sharing accessory subunits of DNA polymerase δ. J. Biol. Chem. 2012, 287, 17281–17287. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.E.; Prakash, L.; Prakash, S. Pol31 and Pol32 subunits of yeast DNA polymerase delta are also essential subunits of DNA polymerase zeta. Proc. Natl. Acad. Sci. USA 2012, 109, 12455–12460. [Google Scholar] [CrossRef]
- Lee, Y.S.; Gregory, M.T.; Yang, W. Human Pol ζ purified with accessory subunits is active in translesion DNA synthesis and complements Pol η in cisplatin bypass. Proc. Natl. Acad. Sci. USA 2014, 111, 2954–2959. [Google Scholar] [CrossRef]
- Reha-Krantz, L.J. DNA polymerase proofreading: Multiple roles maintain genome stability. Biochem. Biophys. Acta 2010, 1804, 1049–1063. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, T.A. DNA replication fidelity. J. Biol. Chem. 2004, 279, 16895–16898. [Google Scholar] [CrossRef] [PubMed]
- McCulloch, S.D.; Kunkel, T.A. The fidelity of DNA synthesis by eukaryotic replicative and translesion synthesis polymerases. Cell Res. 2008, 18, 148–161. [Google Scholar] [CrossRef] [PubMed]
- Moldovan, G.L.; Pfander, B.; Jentsch, S. PCNA, the maestro of the replication fork. Cell 2007, 129, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Dieckman, L.M.; Freudenthal, B.D.; Washington, M.T. PCNA structure and function: Insights from structures of PCNA complexes and post-translationally modified PCNA. In The Eukaryotic Replisome: A Guide to Protein Structure and Function; MacNeill, S., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 281–299. [Google Scholar]
- Boehm, E.M.; Spies, M.; Washington, M.T. PCNA tool belts and polymerase bridges form during translesion synthesis. Nucleic Acids Res. 2016, 44, 8250–8260. [Google Scholar] [CrossRef] [PubMed]
- Choe, K.N.; Moldovan, G.L. Forging ahead through darkness: PCNA, still the principal conductor at the replication fork. Mol. Cell 2017, 65, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Acharya, N.; Patel, S.K.; Sahu, S.R.; Kumari, P. ‘PIPs’ in DNA polymerase: PCNA interaction affairs. Biochem. Soc. Trans. 2020, 48, 2811–2822. [Google Scholar] [CrossRef]
- Prestel, A.; Wichmann, N.; Martins, J.M.; Marabini, R.; Kassem, N.; Broendum, S.S.; Otterlei, M.; Nielsen, O.; Willemoës, M.; Ploug, M.; et al. The PCNA interaction motifs revisited: Thinking outside the PIP-box. Cell. Mol. Life Sci. 2019, 76, 4923–4943. [Google Scholar] [CrossRef]
- Haracska, L.; Unk, I.; Johnson, R.E.; Phillips, B.B.; Hurwitz, J.; Prakash, L.; Prakash, S. Stimulation of DNA synthesis activity of human DNA polymerase kappa by PCNA. Mol. Cell. Biol. 2002, 22, 784–791. [Google Scholar] [CrossRef]
- Acharya, N.; Yoon, J.H.; Gali, H.; Unk, I.; Haracska, L.; Johnson, R.E.; Hurvvitz, J.; Prakash, L.; Prakash, S. Roles of PCNA-binding and ubiquitin-binding domains in human DNA polymerase eta in translesion DNA synthesis. Proc. Natl. Acad. Sci. USA 2008, 105, 17724–17729. [Google Scholar] [CrossRef]
- Yoon, J.-H.; Acharya, N.; Park, J.; Basu, D.; Prakash, S.; Prakash, L. Identification of two functional PCNA-binding domains in human DNA polymerase κ. Genes Cells 2014, 19, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Masuda, Y.; Kanao, R.; Kaji, K.; Ohmori, H.; Hanaoka, F.; Masutani, C. Different types of interaction between PCNA and PIP boxes contribute to distinct cellular functions of Y-family DNA polymerases. Nucleic Acids Res. 2015, 43, 7898–7910. [Google Scholar] [CrossRef] [PubMed]
- Haracska, L.; Acharya, N.; Unk, I.; Johnson, R.E.; Hurwitz, J.; Prakash, L.; Prakash, S. A single domain in human DNA polymerase iota mediates interaction with PCNA: Implications for translesion DNA synthesis. Mol. Cell. Biol. 2005, 25, 1183–1190. [Google Scholar] [CrossRef] [PubMed]
- Vidal, A.E.; Kannouche, P.F.; Podust, V.N.; Yang, W.; Lehmann, A.R.; Woodgate, R. Proliferating cell nuclear antigen-dependent coordination of the biological functions of human DNA polymerase iota. J. Biol. Chem. 2004, 279, 48360–48368. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.X.; Sonoda, E.; Tang, T.S.; Parker, J.L.; Bielen, A.B.; Takeda, S.; Ulrich, H.D.; Friedberg, E.C. REV1 protein interacts with PCNA: Significance of the REV1 BRCT domain in vitro and in vivo. Mol. Cell 2006, 23, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Pustovalova, Y.; Maciejewski, M.W.; Korzhnev, D.M. NMR Mapping of PCNA Interaction with translesion synthesis DNA polymerase Rev1 mediated by Rev1-BRCT domain. J. Mol. Biol. 2013, 425, 3091–3105. [Google Scholar] [CrossRef] [PubMed]
- de Groote, F.H.; Jansen, J.G.; Masuda, Y.; Shah, D.M.; Kamiya, K.; de Wind, N.; Siegal, G. The Rev1 translesion synthesis polymerase has multiple distinct DNA binding modes. DNA Rep. 2011, 10, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Hoege, C.; Pfander, B.; Moldovan, G.-L.; Pyrowolakis, G.; Jentsch, S. RAD6-dependent DNA repair is linked to modification of PCNA by ubiquitin and SUMO. Nature 2002, 419, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Davies, A.A.; Huttner, D.; Daigaku, Y.; Chen, S.; Ulrich, H.D. Activation of ubiquitin-dependent DNA damage bypass is mediated by replication protein A. Mol. Cell 2008, 29, 625–636. [Google Scholar] [CrossRef]
- Ulrich, H.D. Regulating post-translational modifications of the eukaryotic replication clamp PCNA. DNA Rep. 2009, 8, 461–469. [Google Scholar] [CrossRef]
- Friedberg, E.C.; Lehmann, A.R.; Fuchs, R.P.P. Trading places: How do DNA polymerases switch during translesion DNA synthesis? Mol. Cell 2005, 18, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Z.H.; Johnson, R.E.; Haracska, L.; Prakash, L.; Prakash, S.; Benkovic, S.J. Regulation of polymerase exchange between Pol η and Pol δ by monoubiquitination of PCNA and the movement of DNA polymerase holoenzyme. Proc. Natl. Acad. Sci. USA 2008, 105, 5361–5366. [Google Scholar] [CrossRef] [PubMed]
- Freudenthal, B.D.; Gakhar, L.; Ramaswamy, S.; Washington, M.T. Structure of monoubiquitinated PCNA and implications for translesion synthesis and DNA polymerase exchange. Nat. Struct. Mol. Biol. 2010, 17, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Kannouche, P.L.; Wing, J.; Lehmann, A.R. Interaction of human DNA polymerase η with monoubiquitinated PCNA: A possible mechanism for the polymerase switch in response to DNA damage. Mol. Cell 2004, 14, 491–500. [Google Scholar] [CrossRef]
- Bienko, M.; Green, C.M.; Crosetto, N.; Rudolf, F.; Zapart, G.; Coull, B.; Kannouche, P.; Wider, G.; Peter, M.; Lehmann, A.R.; et al. Ubiquitin-binding domains in Y-family polymerases regulate translesion synthesis. Science 2005, 310, 1821–1824. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.E.; Prakash, S.; Prakash, L. Efficient bypass of a thymine-thymine dimer by yeast DNA polymerase Pol η. Science 1999, 283, 1001–1004. [Google Scholar] [CrossRef] [PubMed]
- Prakash, S.; Prakash, L. Translesion DNA synthesis in eukaryotes: A one- or two-polymerase affair. Genes Dev. 2002, 16, 1872–1883. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, C.W. Cellular functions of DNA polymerase ζ and Rev1 protein. Adv. Prot. Chem. 2004, 69, 167–203. [Google Scholar]
- Livneh, Z.; Ziv, O.; Shachar, S. Multiple two-polymerase mechanisms in mammalian translesion DNA synthesis. Cell Cycle 2010, 9, 729–735. [Google Scholar] [CrossRef]
- Shachar, S.; Ziv, O.; Avkin, S.; Adar, S.; Wittschieben, J.; Reißner, T.; Chaney, S.; Friedberg, E.C.; Wang, Z.; Carell, T. Two-polymerase mechanisms dictate error-free and error-prone translesion DNA synthesis in mammals. EMBO J. 2009, 28, 383–393. [Google Scholar] [CrossRef]
- Nelson, J.R.; Lawrence, C.W.; Hinkle, D.C. Deoxycytidyl transferase activity of yeast REV1 protein. Nature 1996, 382, 729–731. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.R.; Gibbs, P.E.; Nowicka, A.M.; Hinkle, D.C.; Lawrence, C.W. Evidence for a second function for Saccharomyces cerevisiae Rev1p. Mol. Microbiol. 2000, 37, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Fischhaber, P.L.; Luk-Paszyc, M.J.; Masuda, Y.; Zhou, J.; Kamiya, K.; Kisker, C.; Friedberg, E.C. Mouse Rev1 protein interacts with multiple DNA polymerases involved in translesion DNA synthesis. EMBO J. 2003, 22, 6621–6630. [Google Scholar] [CrossRef] [PubMed]
- Tissier, A.; Kannouche, P.; Reck, M.-P.; Lehmann, A.R.; Fuchs, R.P.; Cordonnier, A. Co-localization in replication foci and interaction of human Y-family members, DNA polymerase polη and REVl protein. DNA Rep. 2004, 3, 1503–1514. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, S.; Walker, G.C. Novel role for the C-terminus of Saccharomyces cerevisiae Rev1 in mediating protein-protein interactions. Mol. Cell. Biol. 2006, 26, 8173–8182. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, E.; Hanafusa, T.; Kamei, K.; Song, I.; Tomida, J.; Hashimoto, H.; Vaziri, C.; Ohmori, H. Identification of a novel REV1-interacting motif necessary for DNA polymerase kappa function. Genes Cells 2009, 14, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Pustovalova, Y.; Bezsonova, I.; Korzhnev, D.M. The C-terminal domain of human Rev1 contains independent binding sites for DNA polymerase η and Rev7 subunit of polymerase ζ. FEBS Lett. 2012, 586, 3051–3056. [Google Scholar] [CrossRef] [PubMed]
- Pozhidaeva, A.; Pustovalova, Y.; D’Souza, S.; Bezsonova, I.; Walker, G.C.; Korzhnev, D.M. NMR structure and dynamics of the C-terminal domain from human Rev1 and its complex with Rev1 interacting region of DNA polymerase η. Biochemistry 2012, 51, 5506–5520. [Google Scholar] [CrossRef] [PubMed]
- Wojtaszek, J.; Liu, J.; D’Souza, S.; Wang, S.; Xue, Y.; Walker, G.C.; Zhou, P. Multifaceted recognition of vertebrate Rev1 by translesion polymerases ζ and κ. J. Biol. Chem. 2012, 287, 26400–26408. [Google Scholar] [CrossRef]
- Pustovalova, Y.; Magalhaes, M.T.; D’Souza, S.; Rizzo, A.A.; Korza, G.; Walker, G.C.; Korzhnev, D.M. Interaction between the Rev1 C-terminal domain and the PolD3 subunit of Polζ suggests a mechanism of polymerase exchange upon Rev1/Polζ-dependent translesion synthesis. Biochemistry 2016, 55, 2043–2053. [Google Scholar] [CrossRef]
- Kikuchi, S.; Hara, K.; Shimizu, T.; Sato, M.; Hashimoto, H. Structural basis of recruitment of DNA polymerase ζ by interaction between REV1 and REV7 proteins. J. Biol. Chem. 2012, 287, 33847–33852. [Google Scholar] [CrossRef] [PubMed]
- Wojtaszek, J.; Lee, C.J.; D’Souza, S.; Minesinger, B.; Kim, H.; D’Andrea, A.D.; Walker, G.C.; Zhou, P. Structural basis of Rev1-mediated assembly of a quaternary vertebrate translesion polymerase complex consisting of Rev1, heterodimeric Pol ζ and Pol κ. J. Biol. Chem. 2012, 287, 33836–33846. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Yang, X.; Xu, M.; Jiang, T. Structural insights into the assembly of human translesion polymerase complexes. Protein Cell 2012, 3, 864–874. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Rice, W.J.; Malik, R.; Johnson, R.E.; Prakash, L.; Ubarretxena-Belandia, I.; Aggarwal, A.K. Cryo-EM structure and dynamics of eukaryotic DNA polymerase δ holoenzyme. Nat. Struct. Mol. Biol. 2019, 26, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; Georgescu, R.E.; Li, H.; O’Donnell, M.E. Structure of eukaryotic DNA polymerase delta bound to the PCNA clamp while encircling DNA. Proc. Natl. Acad. Sci. USA 2020, 117, 30344–30353. [Google Scholar] [CrossRef] [PubMed]
- Lancey, C.; Tehseen, M.; Raducanu, V.; Rashid, F.; Merino, N.; Ragan, T.J.; Savva, C.G.; Zaher, M.S.; Shirbini, A.; Blanco, F.J.; et al. Structure of the processive human Pol δ holoenzyme. Nat. Commun. 2020, 11, 1109. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Georgescu, R.; Schauer, G.D.; O’Donnell, M.E.; Li, H. Structure of the polymerase ε holoenzyme and atomic model of the leading strand replisome. Nat. Commun. 2020, 11, 3156. [Google Scholar] [CrossRef] [PubMed]
- Netz, D.J.; Stith, C.M.; Stümpfig, M.; Köpf, G.; Vogel, D.; Genau, H.M.; Stodola, J.L.; Lill, R.; Burgers, P.M.; Pierik, A.J. Eukaryotic DNA polymerases require an iron-sulfur cluster for the formation of active complexes. Nat. Chem. Biol. 2012, 8, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, P.E.; McGregor, W.G.; Maher, V.M.; Nisson, P.; Lawrence, C.W. A human homolog of the Saccharomyces cerevisiae REV3 gene, which encodes the catalytic subunit of DNA polymerase zeta. Proc. Natl. Acad. Sci. USA 1998, 95, 6876–6889. [Google Scholar] [CrossRef]
- Tomida, J.; Takata, K.-I.; Lange, S.S.; Schibler, A.C.; Yousefzadeh, M.J.; Bhetawal, S.; Dent, S.Y.R.; Wood, R.D. REV7 is essential for DNA damage tolerance via two REV3L binding sites in mammalian DNA polymerase ζ. Nucleic Acids Res. 2015, 43, 1000–1011. [Google Scholar] [CrossRef]
- Hanafusa, T.; Habu, T.; Tomida, J.; Ohashi, E.; Murakumo, Y.; Ohmori, H. Overlapping in short motif sequences for binding to human REV7 and MAD2 proteins. Genes Cells 2010, 15, 281–296. [Google Scholar] [CrossRef] [PubMed]
- McPherson, K.S.; Rizzo, A.A.; Erlandsen, H.; Chatterjee, N.; Walker, G.C.; Korzhnev, D.M. Evolution of Rev7 interactions in eukaryotic TLS DNA polymerase Polζ. J. Biol. Chem. 2022, 299, 102859. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, A.A.; Vassel, F.M.; Chatterjee, N.; D’Souza, S.; Li, Y.; Hao, B.; Hemann, M.T.; Walker, G.C.; Korzhnev, D.M. Rev7 dimerization is important for assembly and function of the Rev1/Pol zeta translesion synthesis complex. Proc. Natl. Acad. Sci. USA 2018, 115, E8191–E8200. [Google Scholar] [CrossRef] [PubMed]
- Lancey, C.; Tehsen, M.; Bakshi, S.; Percival, M.; Takahashi, M.; Sobhy, M.A.; Raduncanu, V.S.; Muskett, F.W.; Ragan, T.J.; Crehuet, R.; et al. Cryo-EM structure of human Pol κ bound to DNA and mono-ubiquitylated PCNA. Nat. Commun. 2021, 12, 6095. [Google Scholar] [CrossRef] [PubMed]
- Malik, R.; Kopylov, M.; Gomez-Llorente, Y.; Jain, R.; Johnson, R.E.; Prakash, L.; Prakash, S.; Ubarretxena-Belandia, I.; Aggarwal, A.K. Structure and mechanism of B-family DNA polymerase ζ specialized for translesion DNA synthesis. Nat. Struct. Mol. Biol. 2020, 27, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Malik, R.; Johnson, R.E.; Prakash, L.; Prakash, S.; Ubarretxena-Belandia, I.; Aggarwal, A.K. Cryo-EM structure of translesion DNA synthesis polymerase ζ with a base pair mismatch. Nat. Commun. 2022, 12, 1050. [Google Scholar] [CrossRef] [PubMed]
- Du Truong, C.; Craig, T.A.; Cui, G.; Botuyan, M.V.; Serkasevich, R.A.; Chan, K.Y.; Mer, G.; Chiu, P.L.; Kumar, R. Cryo-EM reveals conformational flexibility in apo DNA polymerase zeta. J. Biol. Chem. 2021, 297, 100912. [Google Scholar] [CrossRef] [PubMed]
- Malik, R.; Johnson, R.E.; Ubarretxena-Belandia, I.; Prakash, L.; Prakash, S.; Aggarwal, A.K. Cryo-EM structure of the Rev1-Polζ holocomplex reveals the mechanism of their cooperativity in translesion DNA synthesis. Nat. Struct. Mol. Biol. 2024. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Weng, P.J.; Gao, Y. A new paradigm of DNA synthesis: Three-metal-ion catalysis. Cell Biosci. 2016, 6, 51. [Google Scholar] [CrossRef]
- Chang, C.; Lee Luo, C.; Gao, Y. In crystallo observation of three metal ion promoted DNA polymerase misincorporation. Nat. Commun. 2022, 13, 2346. [Google Scholar] [CrossRef]
- Nevin, P.; Engen, J.R.; Beuning, P.J. Steric gate residues of Y-family DNA polymerases DinB and pol kappa are crucial for dNTP-induced conformational change. DNA Rep. 2015, 29, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Ummat, A.; Silverstein, T.D.; Jain, R.; Buku, A.; Johnson, R.E.; Prakash, L.; Prakash, S.; Aggarwal, A.K. Human DNA polymerase η is pre-aligned for dNTP binding and catalysis. J. Mol. Biol. 2012, 415, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Prindle, M.J.; Loeb, L.A. DNA polymerase delta in dna replication and genome maintenance. Env. Mol. Mutagen. 2012, 53, 666–682. [Google Scholar] [CrossRef] [PubMed]
- Lisova, A.E.; Baranovskiy, A.G.; Morstadt, L.M.; Babayeva, N.D.; Tahirov, T.H. Efficient discrimination against RNA-containing primers by human DNA polymerase ε. Sci. Rep. 2022, 12, 10163. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.A.; Suo, Z. Unlocking the sugar “steric gate” of DNA polymerases. Biochemistry 2011, 50, 1135–1142. [Google Scholar] [CrossRef]
- Joyce, C.M. Choosing the right sugar: How polymerases select a nucleotide substrate. Proc. Natl. Acad. Sci. USA 1997, 94, 1619–1622. [Google Scholar] [CrossRef]
- Vaisman, A.; Woodgate, R. Ribonucleotide discrimination by translesion synthesis DNA polymerases. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 382–402. [Google Scholar] [CrossRef]
- Niimi, N.; Sassa, A.; Katafuchi, A.; Grúz, P.; Fujimoto, H.; Bonala, R.-R.; Johnson, F.; Ohta, T.; Nohmi, T. The steric gate amino acid tyrosine 112 is required for efficient mismatched-primer extension by human DNA polymerase κ. Biochemistry 2009, 48, 4239–4246. [Google Scholar] [CrossRef]
- Weaver, T.M.; Click, T.H.; Khoang, T.H.; Todd Washington, M.; Agarwal, P.K.; Freudenthal, B.D. Mechanism of nucleotide discrimination by the translesion synthesis polymerase Rev1. Nat. Commun. 2022, 13, 2876. [Google Scholar] [CrossRef]
- Donigan, K.A.; McLenigan, M.P.; Yang, W.; Goodman, M.F.; Woodgate, R. The steric gate of DNA polymerase ι regulates ribonucleotide incorporation and deoxyribonucleotide fidelity. J. Biol. Chem. 2014, 289, 9136–9145. [Google Scholar] [CrossRef]
- Donigan, K.A.; Cerritelli, S.M.; McDonald, J.P.; Vaisman, A.; Crouch, R.J.; Woodgate, R. Unlocking the steric gate of DNA polymerase η leads to increased genomic instability in Saccharomyces cerevisiae. DNA Rep. 2015, 35, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gali, V.K.; Balint, E.; Serbyn, N.; Frittmann, O.; Stutz, F.; Unk, I. Translesion synthesis DNA polymerase η exhibits a specific RNA extension activity and a transcription-associated function. Sci. Rep. 2017, 7, 13055. [Google Scholar] [CrossRef] [PubMed]
- Makarova, A.V.; McElhinny, S.A.N.; Watts, B.E.; Kunkel, T.A.; Burgers, P.M. Ribonucleotide incorporation by yeast DNA polymerase ζ. DNA Rep. 2014, 18, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Mentegari, E.; Crespan, E.; Bavagnoli, L.; Kissova, M.; Bertoletti, F.; Sabbioneda, S.; Imhof, R.; Sturla, S.J.; Nilforoushan, A.; Hübscher, U.; et al. Ribonucleotide incorporation by human DNA polymerase η impacts translesion synthesis and RNase H2 activity. Nucleic Acids Res. 2017, 45, 2600–2614. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Egli, M.; Guengerich, F.P. Mechanism of ribonucleotide incorporation by human DNA polymerase η. J. Biol. Chem. 2016, 291, 3747–3756. [Google Scholar] [CrossRef] [PubMed]
- Lone, S.; Townson, S.A.; Uljon, S.N.; Johnson, R.E.; Brahma, A.; Nair, D.T.; Prakash, S.; Prakash, L.; Aggarwal, A.K. Human DNA polymerase kappa encircles DNA: Implications for mismatch extension and lesion bypass. Mol. Cell 2007, 25, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Nair, D.T.; Johnson, R.E.; Prakash, L.; Prakash, S.; Aggarwal, A.K. Rev1 employs a novel mechanism of DNA synthesis using a protein template. Science 2005, 309, 2219–2222. [Google Scholar] [CrossRef] [PubMed]
- Swan, M.K.; Johnson, R.E.; Prakash, L.; Prakash, S.; Aggarwal, A.K. Structure of the human Rev1-DNA-dNTP ternary complex. J. Mol. Biol. 2009, 390, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, Y.; Tang, T.S.; Zhang, H.; Wang, Z.; Friedberg, E.; Yang, W.; Guo, C. Variants of mouse DNA polymerase κ reveal a mechanism of efficient and accurate translesion synthesis past a benzo[a]pyrene dG adduct. Proc. Natl. Acad. Sci. USA 2014, 111, 1789–1794. [Google Scholar] [CrossRef]
- Albertella, M.R.; Green, C.M.; Lehmann, A.R.; O’Connor, M.J. A role for polymerase eta in the cellular tolerance to cisplatin-induced damage. Cancer Res. 2005, 65, 9799–9806. [Google Scholar] [CrossRef]
- Biertümpfel, C.; Zhao, Y.; Kondo, Y.; Ramón-Maiques, S.; Gregory, M.; Lee, J.Y.; Masutani, C.; Lehmann, A.R.; Hanoaka, F.; Yang, W. Structure and mechanism of human DNA polymerase η. Nature 2010, 465, 1044–1048. [Google Scholar] [CrossRef] [PubMed]
- Ummat, A.; Rechkobli, O.; Jain, R.; Choudhury, J.R.; Johnson, R.E.; Silverstein, T.D.; Angeliki, B.; Lone, S.; Prakash, L.; Prakash, S.; et al. Structural basis for cisplatin DNA damage tolerance by human polymerase η during cancer chemotherapy. Nat. Struct. Mol. Biol. 2012, 19, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, T.; Bebenek, K.; Masutani, C.; Rogozin, I.B.; Hanaoka, F.; Kunkel, T.A. Error rate and specificity of human and murine DNA polymerase η. J. Mol. Biol. 2001, 312, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Jha, V.; Ling, H. Structural basis of accurate replication beyond a bulky major benzo[a]pyrene adduct by human DNA polymerase kappa. DNA Rep. 2017, 49, 43–50. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, J. Polymerase iota—An odd sibiling among Y family polymerases. DNA Rep. 2020, 86, 1–11. [Google Scholar] [CrossRef]
- Gomez-Llorente, Y.; Malik, R.; Jain, R.; Choudhury, J.R.; Johnson, R.E.; Prakash, L.; Prakash, S.; Ubarretxena-Belandia, I.; Aggarwal, A.K. The architecture of yeast DNA polymerase zeta. Cell Rep. 2013, 5, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Gulbis, J.M.; Kelman, Z.; Hurwitz, J.; Odonnell, M.; Kuriyan, J. Structure of the C-terminal region of p21(WAF1/CIP1) complexed with human PCNA. Cell 1996, 87, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Hishiki, A.; Hashimoto, H.; Hanafusa, T.; Kamel, K.; Ohashi, E.; Shimizu, T.; Ohmori, H.; Sato, M. Structural basis for novel interactions between human translesion synthesis polymerases and proliferating cell nuclear antigen. J. Biol. Chem. 2009, 284, 10552–10560. [Google Scholar] [CrossRef]
- Bomar, M.G.; Pai, M.T.; Tzeng, S.R.; Li, S.S.C.; Zhou, P. Structure of the ubiquitin-binding zinc finger domain of human DNA Y-polymerase η. EMBO Rep. 2007, 8, 247–251. [Google Scholar] [CrossRef]
- Suzuki, N.; Rohaim, A.; Kato, R.; Dikic, I.; Wakatsuki, S.; Kawasaki, M. A novel mode of ubiquitin recognition by the ubiquitin-binding zinc finger domain of WRNIP1. FEBS J. 2016, 283, 2004–2017. [Google Scholar] [CrossRef]
- Bomar, M.G.; D’Souza, S.; Bienko, M.; Dikic, I.; Walker, G.C.; Zhou, P. Unconventional ubiquitin recognition by the ubiquitin-binding motif within the Y-family DNA polymerases iota and Rev1. Mol. Cell 2010, 37, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Burschowsky, D.; Rudolf, F.; Rabut, G.; Herrmann, T.; Matthias, P.; Wider, G. Structural analysis of the conserved ubiquitin-binding motifs (UBMs) of the translesion polymerase iota in complex with ubiquitin. J. Biol. Chem. 2011, 286, 1364–1373. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Botuyan, M.V.; Mer, G. Structural basis for the interaction of mutasome assembly factor Rev1 with ubiquitin. J. Mol. Biol. 2018, 430, 2042–2050. [Google Scholar] [CrossRef] [PubMed]
- Vanarotti, M.; Grace, C.R.; Miller, D.J.; Actis, M.; Inoue, A.; Evison, B.; Vaithiyalingam, S.; Singh, A.P.; McDonald, E.; Fujii, N. Structures of REV1 UBM2 Domain Complex with Ubiquitin and with a Small-Molecule that Inhibits the REV1 UBM2–Ubiquitin Interaction. J. Mol. Biol. 2018, 430, 2857–2872. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, A.A.; Salerno, P.E.; Bezsonova, I.; Korzhnev, D.M. NMR structure of the human Rad18 zinc finger in complex with ubiquitin defines a class of UBZ domains in proteins linked to the DNA damage response. Biochemistry 2014, 53, 5895–5906. [Google Scholar] [CrossRef] [PubMed]
- Lemontt, J.F. Mutants of yeast defective in mutation induced by ultraviolet light. Genetics 1971, 68, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Jansen, J.G.; Tsaalbi-Shtylik, A.; Langerak, P.; Calléja, F.; Meijers, C.M.; Jacobs, H.; de Wind, N. The BRCT domain of mammalian Rev1 is involved in regulating DNA translesion synthesis. Nucleic Acids Res. 2005, 33, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Pryor, J.M.; Gakhar, L.; Washington, M.T. Structure and functional analysis of the BRCT domain of translesion synthesis DNA polymerase Rev1. Biochemistry 2013, 52, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Chini, C.C.; He, M.; Mer, G.; Chen, J. The BRCT domain is a phospho-protein binding domain. Science 2003, 302, 639–642. [Google Scholar] [CrossRef]
- Doig, A.J.; Baldwin, R.L. N- and C-capping preferences for all 20 amino acids in alpha-helical peptides. Protein Sci. 1995, 4, 1325–1336. [Google Scholar] [CrossRef]
- Boehm, E.M.; Washington, T. R.I.P. to the PIP: PCNA-binding motif no longer considered specific. Bioessays 2016, 38, 1117–1122. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lin, A.; Zhou, C.; Blackwell, S.R.; Zhang, Y.; Wang, Z.; Feng, Q.; Guan, R.; Hanna, M.D.; Chen, Z.; et al. Involvement of budding yeast Rad5 in translesion DNA synthesis through physical interaction with Rev1. Nucleic Acids Res. 2016, 44, 5231–5245. [Google Scholar] [CrossRef] [PubMed]
- Boehm, E.M.; Powers, K.T.; Kondratick, C.M.; Spies, M.; Houtman, J.C.D.; Washington, M.T. The proliferating cell nuclear antigen (PCNA)-interacting protein (PIP) motif of DNA polymerase η mediates its interaction with the C-terminal domain of Rev1. J. Biol. Chem. 2016, 291, 8735–8744. [Google Scholar] [CrossRef] [PubMed]
- Hara, K.; Hashimoto, H.; Murakumo, Y.; Kobayashi, S.; Kogame, T.; Unzai, S.; Akashi, S.; Takeda, S.; Shimizu, T.; Sato, M. Crystal structure of human REV7 in complex with a human REV3 fragment and structural implication of the interaction between DNA polymerase ζ and REV1. J. Biol. Chem. 2010, 285, 12299–12307. [Google Scholar] [CrossRef] [PubMed]
- Baranovskiy, A.G.; Babayeva, N.D.; Liston, V.G.; Rogozin, I.B.; Koonin, E.V.; Pavlov, Y.I.; Vassylyev, D.G.; Tahirov, T.H. X-ray structure of the complex of regulatory subunits of human DNA polymerase delta. Cell Cycle 2008, 7, 3026–3036. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.R.; Lawrence, C.W.; Hinkle, D.C. Thymine-thymine dimer bypass by yeast DNA polymerase ζ. Science 1996, 272, 1646–1649. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Feng, J.; Zuo, P.; Yang, J.; Lu, Y.; Yin, Y. Molecular basis for assembly of the shieldin complex and its implications for NHEJ. Nat. Commun. 2020, 11, 1972. [Google Scholar] [CrossRef] [PubMed]
- Actis, M.; Ambaye, N.; Evison, B.; Shao, Y.; Vanarotti, M.; Inoue, A.; McDonald, E.; Kikuchi, S.; Heath, R.; Hara, K.; et al. Identification of the first small-molecule inhibitor of the REV7 DNA repair protein interaction. Bioorg. Med. Chem. 2016, 24, 4339–4346. [Google Scholar] [CrossRef] [PubMed]
- Wojtaszek, J.L.; Chatterjee, N.; Najeeb, J.; Ramos, A.; Lee, M.; Bian, K.; Xue, J.Y.; Fenton, B.A.; Park, H.; Li, D.; et al. A small molecule targeting mutagenic translesion synthesis improves chemotherapy. Cell 2019, 178, 152–159. [Google Scholar] [CrossRef]
- Chatterjee, N.; Whitman, M.A.; Harris, C.A.; Min, S.M.; Jonas, O.; Lien, E.C.; Luengo, A.; Vander Heiden, M.G.; Hong, J.; Zhou, P.; et al. REV1 inhibitor JH-RE-06 enhances tumor cell response to chemotherapy by triggering senescence hallmarks. Proc. Natl. Acad. Sci. USA 2020, 117, 28918–28921. [Google Scholar] [CrossRef]
- Sail, V.; Rizzo, A.A.; Chatterjee, N.; Dash, R.C.; Ozen, Z.; Walker, G.C.; Korzhnev, D.M.; Hadden, K. Identification of Small Molecule Translesion Synthesis Inhibitors That Target the Rev1-CT/RIR Protein−Protein Interaction. ACS Chem. Biol. 2017, 12, 1903–1912. [Google Scholar] [CrossRef] [PubMed]
- Ozen, Z.; Dash, R.C.; McCarthy, K.R.; Chow, S.A.; Rizzo, A.A.; Korzhnev, D.M.; Hadden, M.K. Small molecule scaffolds that disrupt the Rev1-CT/RIR protein-protein interaction. Bioorg. Med. Chem. 2018, 26, 4301–4309. [Google Scholar] [CrossRef] [PubMed]
- Dash, R.C.; Ozen, Z.; Rizzo, A.A.; Lim, S.; Korzhnev, D.M.; Hadden, M.K. Structural approach to identify a lead scaffold that targets the translesion synthesis polymerase Rev1. J. Chem. Inf. Model. 2018, 58, 2266–2277. [Google Scholar] [CrossRef] [PubMed]
- Dash, R.C.; Ozen, Z.; McCarthy, K.R.; Chatterjee, N.; Harris, C.A.; Rizzo, A.A.; Walker, G.C.; Korzhnev, D.M.; Hadden, M.K. Virtual pharmacophore screening identifies small-molecule inhibitors of the Rev1-CT/RIR protein-protein pnteraction. ChemMedChem 2019, 14, 1610–1617. [Google Scholar] [CrossRef]
- McPherson, K.S.; Zaino, A.M.; Dash, R.C.; Rizzo, A.A.; Li, Y.; Hao, B.; Bezsonova, I.; Hadden, M.K.; Korzhnev, D.M. Structure-based drug design of phenazopyridine derivatives as inhibitors of Rev1 interactions in translesion synthesis. ChemMedChem 2021, 16, 1126–1132. [Google Scholar] [CrossRef]
Figure 1.
TLS DNA polymerases in humans and yeast. (A,B) Domains and subunits of (A) human and (B) yeast Y-family polymerases Polη, Polι, Polκ, Rev1, and B-family polymerase Pol ζ. The core catalytic domain of each Y-family polymerase (including fingers, thumb, and palm subdomains) is followed by a polymerase associated domain (PAD). B-family Polζ lacks the PAD domain but includes an inactive endonuclease domain within the catalytic core. The binding modules of Y-family TLS polymerases include PCNA-interacting PIP and Rev1-interacting RIR motifs, ubiquitin-binding UBM (Rev1, Polι) and UBZ (Polη, Polκ) domains, Rev1 N-terminal BRCT and the C-terminal CT domains, and the N-terminal α-helical region of Rev1 termed the “M1” motif. Polη includes a nuclear localization signal (NLS). Polζ comprises four subunits: Rev3, Rev7, PolD2/PolD3 (humans), or Pol31/Pol32 (yeast). The Rev3 subunit of Polζ harbors a catalytic core domain, an N-terminal domain (NTD), a positively charged domain (PCD, humans), two Rev7-binding motifs (RBMs), and a C-terminal domain (CTD) interacting with the PolD2/PolD3 or Pol31/Pol32 module. (C) Schematic of Rev1/Polζ-dependent TLS mutasome assembled on ubiquitinated PCNA, including an ‘inserter’ TLS polymerase Polη, and ‘extender’ TLS polymerase Polζ and a scaffold protein Rev1. Possible interactions between domains and subunits of the TLS proteins are indicated. Adapted with permission from [32].
Figure 1.
TLS DNA polymerases in humans and yeast. (A,B) Domains and subunits of (A) human and (B) yeast Y-family polymerases Polη, Polι, Polκ, Rev1, and B-family polymerase Pol ζ. The core catalytic domain of each Y-family polymerase (including fingers, thumb, and palm subdomains) is followed by a polymerase associated domain (PAD). B-family Polζ lacks the PAD domain but includes an inactive endonuclease domain within the catalytic core. The binding modules of Y-family TLS polymerases include PCNA-interacting PIP and Rev1-interacting RIR motifs, ubiquitin-binding UBM (Rev1, Polι) and UBZ (Polη, Polκ) domains, Rev1 N-terminal BRCT and the C-terminal CT domains, and the N-terminal α-helical region of Rev1 termed the “M1” motif. Polη includes a nuclear localization signal (NLS). Polζ comprises four subunits: Rev3, Rev7, PolD2/PolD3 (humans), or Pol31/Pol32 (yeast). The Rev3 subunit of Polζ harbors a catalytic core domain, an N-terminal domain (NTD), a positively charged domain (PCD, humans), two Rev7-binding motifs (RBMs), and a C-terminal domain (CTD) interacting with the PolD2/PolD3 or Pol31/Pol32 module. (C) Schematic of Rev1/Polζ-dependent TLS mutasome assembled on ubiquitinated PCNA, including an ‘inserter’ TLS polymerase Polη, and ‘extender’ TLS polymerase Polζ and a scaffold protein Rev1. Possible interactions between domains and subunits of the TLS proteins are indicated. Adapted with permission from [32].
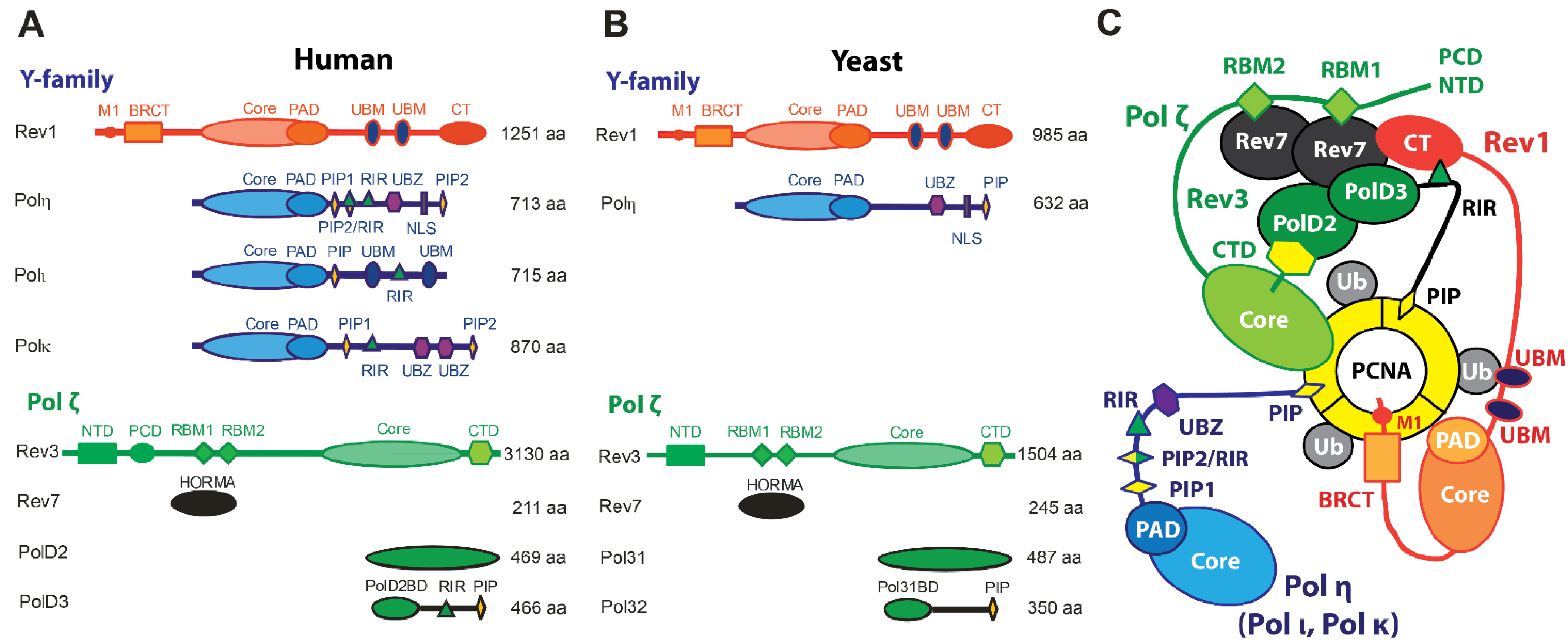
Figure 2.
Catalytic domains of human Y-family TLS DNA polymerases. Each Y-family polymerase features fingers (blue), palm (red), thumb (green), and PAD (yellow) domains. Insets for A–D show close-ups of polymerase active sites in the process of nucleotide insertion. (A) Hs Polη bypassing a cisplatin DNA adduct. (B) Polκ bypassing benzo[a]pyrene adduct. (C) Hs Polι bypassing 8-oxoG lesion. (D) Hs Rev1 inserting cytosine on a normal p/t DNA.
Figure 2.
Catalytic domains of human Y-family TLS DNA polymerases. Each Y-family polymerase features fingers (blue), palm (red), thumb (green), and PAD (yellow) domains. Insets for A–D show close-ups of polymerase active sites in the process of nucleotide insertion. (A) Hs Polη bypassing a cisplatin DNA adduct. (B) Polκ bypassing benzo[a]pyrene adduct. (C) Hs Polι bypassing 8-oxoG lesion. (D) Hs Rev1 inserting cytosine on a normal p/t DNA.

Figure 3.
Cryo-EM structure of the Sc Polζ catalytic domain. (A) The Sc Rev3 catalytic domain on p/t DNA, including the fingers (yellow), thumb (orange), palm (cyan), NTD (blue), and inactive exonuclease (purple) domains [97]. The CysB domain within Rev3 CTD that binds Pol31 is also shown. The inset shows a close-up of the Rev3 active site, revealing the basis for Polζ intolerance to mismatches during nucleotide insertion. Residues from the fingers (yellow) and palm (blue) domains make extensive contacts with the nascent base pair (red). (B) Structural basis for Polζ extension past DNA mismatches [97]. Due to contacts with αxC and palm-loop, the NTD-palm linker in the Rev3 active site is held further away from DNA than in Pol3. (C) Structural basis of the impaired exonuclease activity of Polζ [97]. Unlike Pol3, Rev3 lacks the β-hairpin used to pass mismatched DNA from the active site to the exonuclease domain.
Figure 3.
Cryo-EM structure of the Sc Polζ catalytic domain. (A) The Sc Rev3 catalytic domain on p/t DNA, including the fingers (yellow), thumb (orange), palm (cyan), NTD (blue), and inactive exonuclease (purple) domains [97]. The CysB domain within Rev3 CTD that binds Pol31 is also shown. The inset shows a close-up of the Rev3 active site, revealing the basis for Polζ intolerance to mismatches during nucleotide insertion. Residues from the fingers (yellow) and palm (blue) domains make extensive contacts with the nascent base pair (red). (B) Structural basis for Polζ extension past DNA mismatches [97]. Due to contacts with αxC and palm-loop, the NTD-palm linker in the Rev3 active site is held further away from DNA than in Pol3. (C) Structural basis of the impaired exonuclease activity of Polζ [97]. Unlike Pol3, Rev3 lacks the β-hairpin used to pass mismatched DNA from the active site to the exonuclease domain.

Figure 5.
Cryo-EM structure of the human Polκ holoenzyme. (A) Polκ bound to p/t DNA (gray) and the front face of unmodified PCNA (blue). The catalytic core of Polκ comprises the palm (tan), fingers (yellow), thumb (orange), N-clasp (pink), and PAD domains (green). An ‘inverting’ helix C-terminal to PAD orients the internal Polκ PIP-box motif (red) towards the binding site on the PCNA1 protomer. The right panel shows a structural comparison of the Hs Polκ PIP1 and Hs Polδ PIP motifs bound to a PCNA protomer. (B) Polκ in complex with p/t DNA and mono-ubiquitinated PCNA with ubiquitin moieties visible on the PCNA back face. (C) DNA that exits the Polκ holoenzyme tilts 47° relative to the PCNA axis, resulting in DNA bending when passing through the PCNA ring. (D) A model of a PCNA ‘tool belt’ that simultaneously binds to the Polκ/DNA (yellow/gray) complex and apo Polδ (green/magenta). Adapted with permission from [96].
Figure 5.
Cryo-EM structure of the human Polκ holoenzyme. (A) Polκ bound to p/t DNA (gray) and the front face of unmodified PCNA (blue). The catalytic core of Polκ comprises the palm (tan), fingers (yellow), thumb (orange), N-clasp (pink), and PAD domains (green). An ‘inverting’ helix C-terminal to PAD orients the internal Polκ PIP-box motif (red) towards the binding site on the PCNA1 protomer. The right panel shows a structural comparison of the Hs Polκ PIP1 and Hs Polδ PIP motifs bound to a PCNA protomer. (B) Polκ in complex with p/t DNA and mono-ubiquitinated PCNA with ubiquitin moieties visible on the PCNA back face. (C) DNA that exits the Polκ holoenzyme tilts 47° relative to the PCNA axis, resulting in DNA bending when passing through the PCNA ring. (D) A model of a PCNA ‘tool belt’ that simultaneously binds to the Polκ/DNA (yellow/gray) complex and apo Polδ (green/magenta). Adapted with permission from [96].
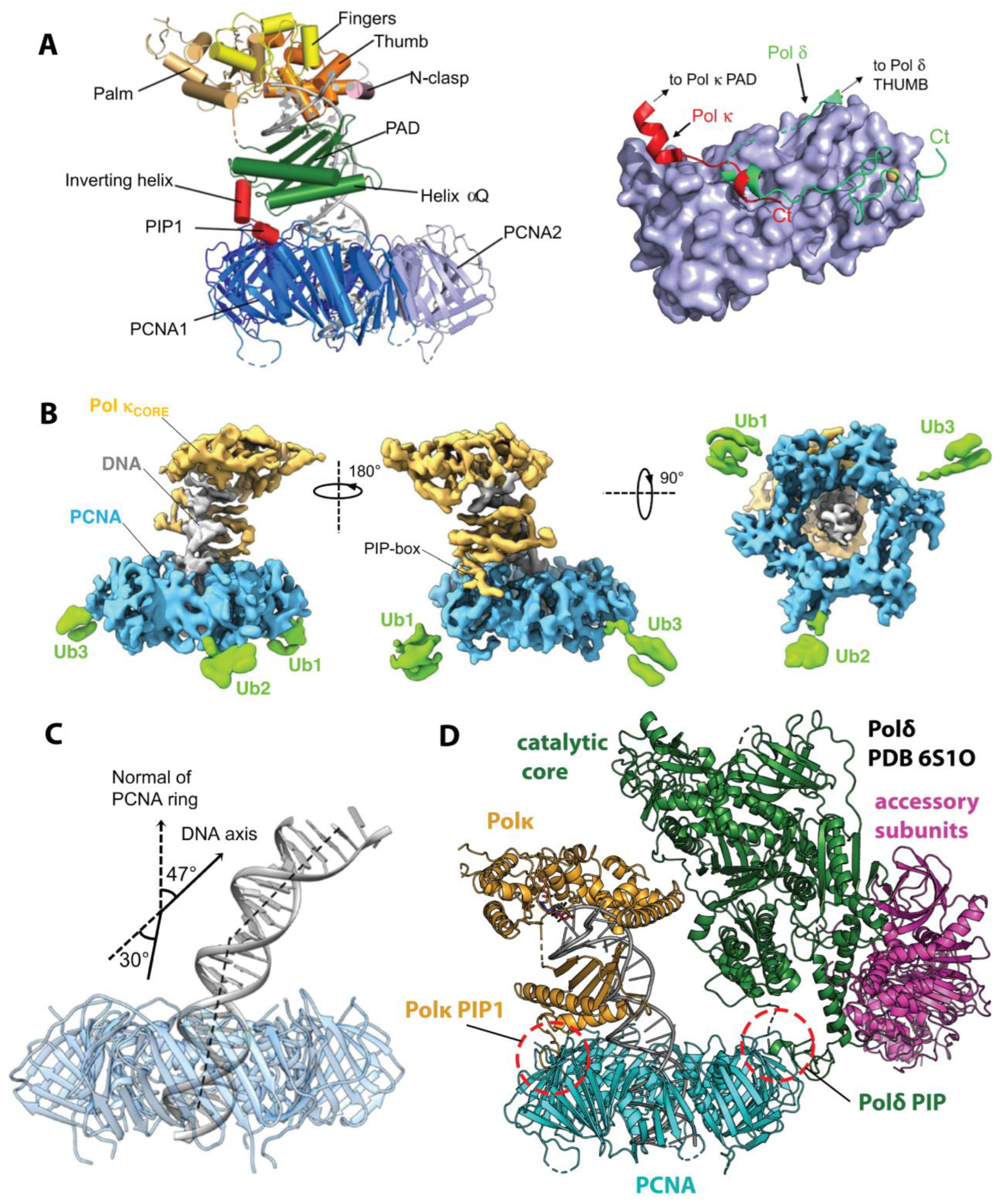
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Arianna, G.A.; Korzhnev, D.M. Protein Assemblies in Translesion Synthesis. Genes 2024, 15, 832. https://doi.org/10.3390/genes15070832
AMA Style
Arianna GA, Korzhnev DM. Protein Assemblies in Translesion Synthesis. Genes. 2024; 15(7):832. https://doi.org/10.3390/genes15070832
Chicago/Turabian StyleArianna, Gianluca A., and Dmitry M. Korzhnev. 2024. "Protein Assemblies in Translesion Synthesis" Genes 15, no. 7: 832. https://doi.org/10.3390/genes15070832
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.