
Expression Patterns of TGF-β1, TβR-I, TβR-II, and Smad2 Reveal Insights into Heterosis for Growth of Hybrid Offspring between Acanthopagrus schlegelii and Pagrus major


Abstract
:1. Introduction
2. Material and Methods
2.1. Ethics Statement
2.2. Fish Sampling
2.3. Sequence Analyses
2.4. RNA Extraction and Real-Time Polymerase Chain Reaction Detection
2.5. Western Blot
2.6. Statistical Analyses
3. Results
3.1. Sequence Analysis of AsTGF-β1, AsTβR-II, and AsSmad2
3.2. Expression of AsTGF-β1, AsTβR-I, AsTβR-II, and AsSmad2 mRNA in Different Developmental Stages of A. schlegelii
3.3. Expression of TGF-β1, TβR-I, TβR-II, and Smad2 mRNA in Different Tissues of A. schlegelii, P. major, and Their Hybrids
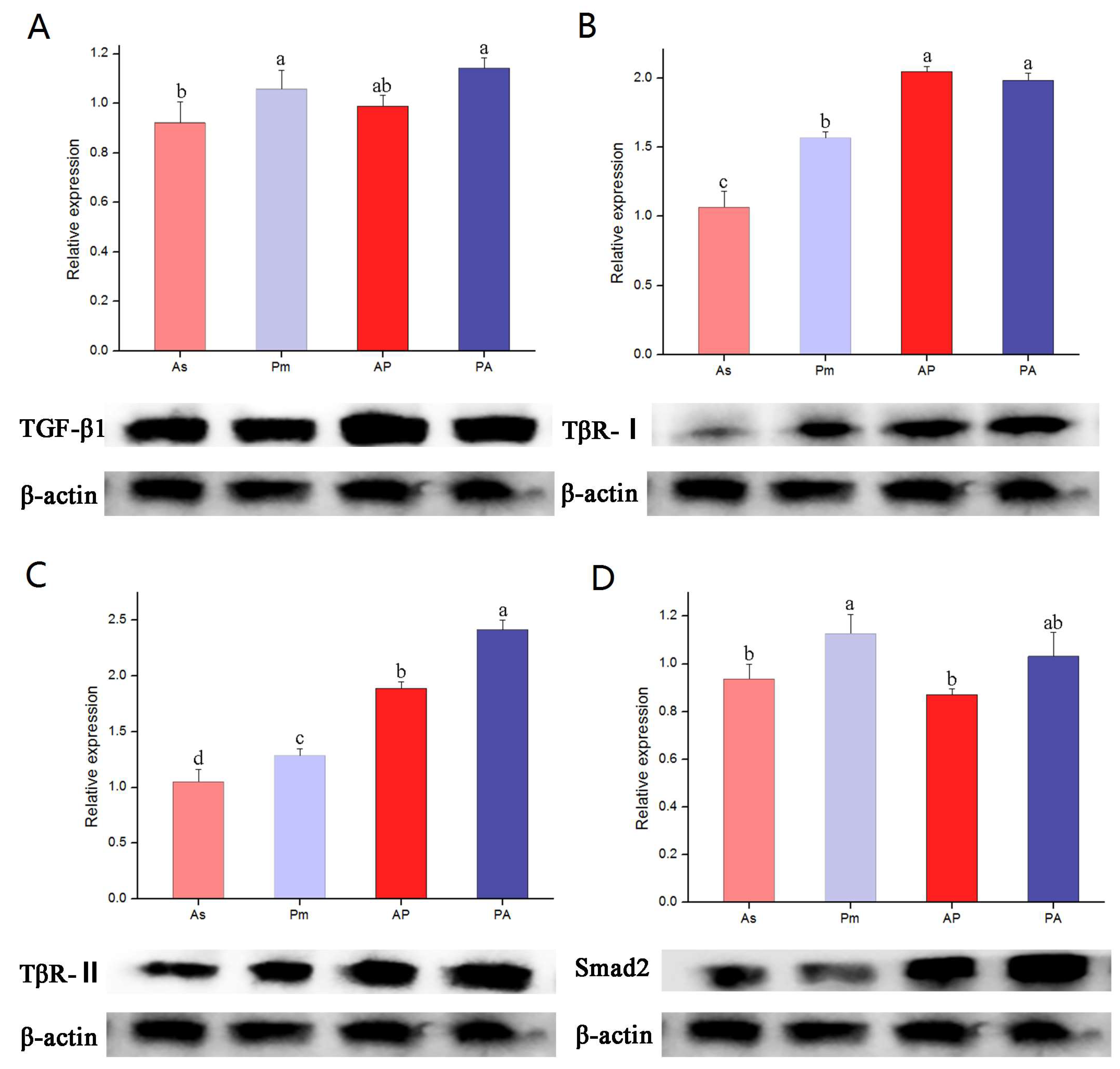
3.4. The Expression of TGF-β1, TβR-I, TβR-II, and Smad2 Genes and Proteins in Larvae of A. schlegelii, P. major, AP, and PA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gorshkov, S.; Gorshkova, G.; Hadani, A.; Gordin, H.; Knibb, W. Chromosome set manipulations and hybridization experiments in gilthead seabream (Sparus aurata). II. Assessment of diploid and triploid hybrids between gilthead seabream and red seabream (Pagrus major). J. Appl. Ichthyol. 2002, 18, 106–112. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, Z.; Ji, H.; Li, P.; Zhao, Y.; Zhang, Z. Cloning and expression analysis of Calmodulin from the hybrid F1 of Acanthopagrus schlegelii male × Pagrus major female and P. major. Mar. Fish. 2018, 40, 435–446. [Google Scholar]
- Chen, S.; Zhang, Z.; Liu, H.; Jia, C.; Xu, J.; Zhu, F.; Zhang, Z.; Ren, Z. Backcross Breeding and Observation of Early Larva Development in Red Bream Pagrosomus major and Black Bream Acanthopagrus schiegelii. Fish. Sci. 2017, 36, 693–699. [Google Scholar]
- Yu, Y.Y. Genetic Analysis of the Inter Genus Hybridization of Pagrosomus major (♀) and Spraus macrocephalus (♂). Master’s Thesis, Ningbo University, Ningbo, China, 2012. [Google Scholar]
- Wang, S.T. Effects of Salinity on the Antioxidant System of Black Porgy (Acanthopagrus schlegelii) and Comparison of Fatty Acid Composition between Hybrid Snapper and Its Parents. Master’s Thesis, Nanjing Normal Universuty, Nanjing, China, 2018. [Google Scholar]
- Wang, S.T.; Li, L.; Chen, S.Y.; Zhang, Z.Y.; Yin, S.W.; Zhu, F.; Jia, C.F.; Liu, H.L. Lipid Content and Fatty Acid Profile of Acanthopagrus schlegeli, Pagrosomus major and Their Hybrids. Food Sci. 2016, 37, 81–85. [Google Scholar]
- Chen, J.L.; Colgan, T.D.; Walton, K.L.; Gregorevic, P.; Harrison, C.A. The TGF-β Signalling Network in Muscle Development, Adaptation and Disease. Adv. Exp. Med. Biol. 2016, 900, 97–131. [Google Scholar] [PubMed]
- Shi, Y. Structural insights on Smad function in TGFβ signaling. BioEssays News Rev. Mol. Cell. Dev. Biol. 2001, 23, 223–232. [Google Scholar] [CrossRef]
- Chen, M.; Lv, Z.; Huang, L.; Zhang, W.; Lin, X.; Shi, J.; Zhang, W.; Liang, R.; Jiang, S. Triptolide inhibits TGF-β1-induced cell proliferation in rat airway smooth muscle cells by suppressing Smad signaling. Exp. Cell Res. 2015, 331, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S.; Huang, J.S. TGF-β control of cell proliferation. J. Cell. Biochem. 2005, 96, 447–462. [Google Scholar] [CrossRef] [PubMed]
- Massagué, J.; Blain, S.W.; Lo, R.S. TGFβ signaling in growth control, cancer, and heritable disorders. Cell 2000, 103, 295–309. [Google Scholar] [CrossRef]
- Wrana, J.L. Phosphoserine-dependent regulation of protein-protein interactions in the Smad pathway. Structure 2002, 10, 5–7. [Google Scholar] [CrossRef]
- Derynck, R.; Zhang, Y.; Feng, X.H. Smads: Transcriptional activators of TGF-β responses. Cell 1998, 95, 737–740. [Google Scholar] [CrossRef] [PubMed]
- Mehra, A.; Attisano, L.; Wrana, J.L. Characterization of Smad phosphorylation and Smad-receptor interaction. Methods Mol. Biol. 2000, 142, 67–78. [Google Scholar] [PubMed]
- Derynck, R.; Zhang, Y.E. Smad-dependent and Smad-independent pathways in TGF-β family signalling. Nature 2003, 425, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Wang, Y.F.; Jayaraman, L.; Yang, H.; Massagué, J.; Pavletich, N.P. Crystal structure of a Smad MH1 domain bound to DNA: Insights on DNA binding in TGF-β signaling. Cell 1998, 94, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Wrana, J.L.; Attisano, L. The Smad pathway. Cytokine Growth Factor Rev. 2000, 11, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Lau, M.T.; Ge, W. Cloning of Smad2, Smad3, Smad4, and Smad7 from the goldfish pituitary and evidence for their involvement in activin regulation of goldfish FSHβ promoter activity. Gen. Comp. Endocrinol. 2005, 141, 22–38. [Google Scholar] [CrossRef] [PubMed]
- Frolik, C.A.; Dart, L.L.; Meyers, C.A.; Smith, D.M.; Sporn, M.B. Purification and initial characterization of a type beta transforming growth factor from human placenta. Proc. Natl. Acad. Sci. USA 1983, 80, 3676–3680. [Google Scholar] [CrossRef] [PubMed]
- Müller, F.; Blader, P.; Rastegar, S.; Fischer, N.; Knöchel, W.; Strähle, U. Characterization of zebrafish smad1, smad2 and smad5: The amino-terminus of smad1 and smad5 is required for specific function in the embryo. Mech. Dev. 1999, 88, 73–88. [Google Scholar] [CrossRef]
- Johnson, A.K.; Harms, C.A.; Levine, J.F.; Law, J.M. A quantitative real-time RT-PCR assay to measure TGF-β mRNA and its correlation with hematologic, plasma chemistry and organo-somatic indices responses in triamcinolone-treated Atlantic menhaden, Brevoortia tyrannus. Dev. Comp. Immunol. 2006, 30, 473–484. [Google Scholar] [CrossRef]
- Kohli, G.; Hu, S.; Clelland, E.; Di Muccio, T.; Rothenstein, J.; Peng, C. Cloning of transforming growth factor-β1 (TGF-β1) and its type II receptor from zebrafish ovary and role of TGF-β1 in oocyte maturation. Endocrinology 2003, 144, 1931–1941. [Google Scholar] [CrossRef]
- Tan, Q.; Zagrodny, A.; Bernaudo, S.; Peng, C. Regulation of membrane progestin receptors in the zebrafish ovary by gonadotropin, activin, TGF-β and BMP-15. Mol. Cell. Endocrinol. 2009, 312, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.X.; Yi, X.J.; Gu, P.L.; Gao, S.X. The role of KDR in intrauterine adhesions may involve the TGF-β1/Smads signaling pathway. Braz. J. Med. Biol. Res. 2019, 52, e8324. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Zhu, Z.; Cui, G.; Zhang, Z.; Yue, J.; Zhang, Y.; Yao, X.; Huo, M.; Wei, J.; Wang, Q.; et al. Gastric cancer derived exosomes induce peritoneal mesothelial cell EMT through TGF-β1/Smads pathway to promote peritoneal metastasis. bioRxiv 2021. [Google Scholar] [CrossRef]
- Holopainen, R.; Tapiovaara, H.; Honkanen, J. Expression analysis of immune response genes in fish epithelial cells following ranavirus infection. Fish Shellfish Immunol. 2012, 32, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.S.W.; Chang, C.; Liu, D.; Derynck, R. Sumoylation of Smad4, the common Smad mediator of transforming growth factor-β family signaling. J. Biol. Chem. 2003, 278, 27853–27863. [Google Scholar] [CrossRef] [PubMed]
- Yu, E.; Xie, J.; Wang, G.; Yu, D.; Gong, W.; Li, Z.; Wang, H.; Xia, Y.; Wei, N. Gene Expression Profiling of Grass Carp (Ctenopharyngodon idellus) and Crisp Grass Carp. Int. J. Genom. 2014, 2014, 639687. [Google Scholar] [CrossRef] [PubMed]
- Zhan, X.L.; Ma, T.Y.; Wu, J.Y.; Yi, L.Y.; Wang, J.Y.; Gao, X.K.; Li, W.S. Cloning and primary immunological study of TGF-β1 and its receptors TβR I/TβR II in tilapia (Oreochromis niloticus). Dev. Comp. Immunol. 2015, 51, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Rindom, E.; Vissing, K. Mechanosensitive Molecular Networks Involved in Transducing Resistance Exercise-Signals into Muscle Protein Accretion. Front. Physiol. 2016, 7, 547. [Google Scholar] [CrossRef] [PubMed]
- Lilleeng, E.; Penn, M.H.; Haugland, O.; Xu, C.; Bakke, A.M.; Krogdahl, A.; Landsverk, T.; Frøystad-Saugen, M.K. Decreased expression of TGF-β, GILT and T-cell markers in the early stages of soybean enteropathy in Atlantic salmon (Salmo salar L.). Fish Shellfish Immunol. 2009, 27, 65–72. [Google Scholar] [CrossRef]
- Maehr, T.; Wang, T.; González Vecino, J.L.; Wadsworth, S.; Secombes, C.J. Cloning and expression analysis of the transforming growth factor-β receptors type 1 and 2 in the rainbow trout Oncorhynchus mykiss. Dev. Comp. Immunol. 2012, 37, 115–126. [Google Scholar] [CrossRef]
- Haddad, G.; Hanington, P.C.; Wilson, E.C.; Grayfer, L.; Belosevic, M. Molecular and functional characterization of goldfish (Carassius auratus L.) transforming growth factor beta. Dev. Comp. Immunol. 2008, 32, 654–663. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Jimmy, K. Molecular isolation and characterisation of carp transforming growth factor β1 from activated leucocytes. Fish Shellfish Immunol. 2000, 10, 309–318. [Google Scholar] [CrossRef]
- Hobbie, K.R.; DeAngelo, A.B.; George, M.H.; Law, J.M. Neoplastic and nonneoplastic liver lesions induced by dimethylnitrosamine in Japanese medaka fish. Vet. Pathol. 2012, 49, 372–385. [Google Scholar] [CrossRef]
- Cai, Z.; Gao, C.; Li, L.; Xing, K. Bipolar properties of red seabream (Pagrus major) transforming growth factor-β in induction of the leucocytes migration. Fish Shellfish Immunol. 2010, 28, 695–700. [Google Scholar] [CrossRef]
- Annes, J.P.; Munger, J.S.; Rifkin, D.B. Making sense of latent TGFβ activation. J. Cell Sci. 2003, 116 Pt 2, 217–224. [Google Scholar] [CrossRef]
- Heldin, C.H.; Miyazono, K.; ten Dijke, P. TGF-β signalling from cell membrane to nucleus through SMAD proteins. Nature 1997, 390, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.W.; Godson, C.; Brazil, D.P.; Martin, F. Extracellular BMP-antagonist regulation in development and disease: Tied up in knots. Trends Cell Biol. 2010, 20, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Wu, X. Comparative Study on Temperature Tolerance and Growth between Pagrus major (Temminck et Schlegel) and Spraus macrocephalus and F1 Hybrid. Mod. Fish. Inf. 2009, 24, 26–28. [Google Scholar]
- Yan, F.; Xu, S.; Gu, J.; Chen, X.; Lyu, H.; Jia, C. Comparison of metabolic and excretion rates of young Sparus macrocephalus and young hybrid porgy. J. Oceanogr. Taiwan Strait 2010, 29, 496–502. [Google Scholar]
- Lin, M.; Miao, L.; Li, M.Y.; Yu, Y.Y.; Xu, W.T. Chromosome composition and AFLP analysis of Pagrosomus major, Spraus macrocephalus and the hybirds. J. Oceanogr. Chin. Version 2014, 36, 117–121. [Google Scholar]
- Harms, C.A.; Howard, K.E.; Wolf, J.C.; Smith, S.A.; Kennedy-Stoskopf, S. Transforming growth factor-β response to mycobacterial infection in striped bass Morone saxatilis and hybrid tilapia Oreochromis spp. Vet. Immunol. Immunopathol. 2003, 95, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Harms, C.A.; Kennedy-Stoskopf, S.; Horne, W.A.; Fuller, F.J.; Tompkins, W.A. Cloning and sequencing hybrid striped bass (Morone saxatilis x M. chrysops) transforming growth factor-β (TGF-β), and development of a reverse transcription quantitative competitive polymerase chain reaction (RT-qcPCR) assay to measure TGF-β mRNA of teleost fish. Fish Shellfish Immunol. 2000, 10, 61–85. [Google Scholar] [PubMed]
- Zi, Z.; Chapnick, D.A.; Liu, X. Dynamics of TGF-β/Smad signaling. FEBS Lett. 2012, 586, 1921–1928. [Google Scholar] [CrossRef] [PubMed]
- Gaarenstroom, T.; Hill, C.S. TGF-β signaling to chromatin: How Smads regulate transcription during self-renewal and differentiation. Semin. Cell Dev. Biol. 2014, 32, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.Y.; Lam, S.S.; Correia, J.J.; Lin, K. Smad3 allostery links TGF-β receptor kinase activation to transcriptional control. Genes Dev. 2002, 16, 1950–1963. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.M.; Nikolic-Paterson, D.J.; Lan, H.Y. TGF-β: The master regulator of fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef]
- Attisano, L.; Wrana, J.L. Smads as transcriptional co-modulators. Curr. Opin. Cell Biol. 2000, 12, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Attisano, L. Mutations in the tumor suppressors Smad2 and Smad4 inactivate transforming growth factor β signaling by targeting Smads to the ubiquitin-proteasome pathway. Proc. Natl. Acad. Sci. USA 2000, 97, 4820–4825. [Google Scholar] [CrossRef]
- Cao, G. Comparative Research of Genetic Characterization in Acanthopagrus schlegelii, Pagrus major and Their Hybrid Offsprings. Master’s Thesis, Shanghai Ocean University, Shanghai, China, 2021. [Google Scholar]
- Chen, Y.; Fan, Y.; Guo, D.Y.; Xu, B.; Shi, X.Y.; Li, J.T.; Duan, L.F. Study on the relationship between hepatic fibrosis and epithelial-mesenchymal transition in intrahepatic cells. Biomed. Pharmacother. 2020, 129, 110413. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primer (5′-3′) | Reverse Primer (3′-5′) |
|---|---|---|
| AsTGF-β1 | TTGGCACCATGAAGCTGGTA | TGAGGCTCTGTTGGTAAGCG |
| PmTGF-β1 | TGGCTGTCCTTCGATGTCAC | TGGCTGTCCTTCGATGTCAC |
| APTGF-β1 | CTCATCAGGCTGGAGACGAG | ATGTCGGTCTGGACCTCAGT |
| PATGF-β1 | ACTGAGGTCCAGACCGACAT | TCAGCAGGCGGTAATCTCCA |
| AsTβR-I | CTCAGGCTCTGGATCAGGTC | TCTCCTGGAGGATGATGGTC |
| AsTβR-II | CATAACCCCACTCTGCCACT | GCCACAGACCATCATGACAC |
| PmTβR-I | CACCATCGACATCGCTCCAA | CGTAGTAGGGCAGCTGGTAG |
| PmTβR-II | GACATGCTGCCTGTGTGACT | GCTGTACCCACCTGTCCACT |
| APTβR-I | CACCATCGACATCGCTCCAA | CGTAGTAGGGCAGCTGGTAG |
| APTβR-II | TGGTGCCTCTGCTGGTTATG | GCGTGGAGCTACTGTCTGAA |
| PATβR-I | CCTCAATGGACCACCCCTTC | TCCTCACCACGCCATTTACC |
| PATβR-II | TGATGTTCGACAAGGGAGCC | CGGTAGAAGTAGAAGGCGGC |
| AsSmad2 | ACGCAAGCCCACGCATAC | TCTTCCAACCCAGGAGTCGTT |
| Asβ-actin | GGACCTGTATGCCAACACTG | TGATCTCCTTCTGCATCCTG |
| PmSmad2 | ACCTTCACAGCCATCACGAG | GGTCTCCACCCTCTGGTAGT |
| Pmβ-actin | TATCGTCATGGACTCCGGTG | TGATGTCACGCACGATTTCC |
| APSmad2 | GAGGCCAGCGATCAACAGAT | CTGGCTCCGAGTAAGTCACC |
| APβ-actin | CAGAGCGTGGCTACTCCTTC | CCATCTCCTGCTCGAAGTCC |
| PASmad2 | CTCCAGCCGGTGACTTACTC | CTCCACAGTGGCATTCCTGT |
| PAβ-actin | GGCTACTCCTTCACCACCAC | CCGATGGTGATGACCTGTCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, X.; Zhao, Y.; Li, J.; Xie, W.; Lyu, L.; Chen, S.; Jia, C.; Yan, J.; Li, P. Expression Patterns of TGF-β1, TβR-I, TβR-II, and Smad2 Reveal Insights into Heterosis for Growth of Hybrid Offspring between Acanthopagrus schlegelii and Pagrus major. Genes 2024, 15, 945. https://doi.org/10.3390/genes15070945
Du X, Zhao Y, Li J, Xie W, Lyu L, Chen S, Jia C, Yan J, Li P. Expression Patterns of TGF-β1, TβR-I, TβR-II, and Smad2 Reveal Insights into Heterosis for Growth of Hybrid Offspring between Acanthopagrus schlegelii and Pagrus major. Genes. 2024; 15(7):945. https://doi.org/10.3390/genes15070945
Chicago/Turabian StyleDu, Xinran, Yue Zhao, Jingbo Li, Wenli Xie, Linna Lyu, Shuyin Chen, Chaofeng Jia, Jie Yan, and Peng Li. 2024. "Expression Patterns of TGF-β1, TβR-I, TβR-II, and Smad2 Reveal Insights into Heterosis for Growth of Hybrid Offspring between Acanthopagrus schlegelii and Pagrus major" Genes 15, no. 7: 945. https://doi.org/10.3390/genes15070945